Nuclear Lamin is required for Winged Eye‐
mediated transdetermination of Drosophila
imaginal disc
著者
Keita Masuko, Hirofumi Furuhashi, Kanae
Komaba, Eriko Numao, Rumi Nakajima, Naoyuki
Fuse, Shoichiro Kurata
journal or
publication title
Genes to cells : devoted to molecular &
cellular mechanisms
volume
23
number
8
page range
724-731
year
2018-07-02
URL
http://hdl.handle.net/10097/00126010
doi: 10.1111/gtc.12608Title
Nuclear Lamin is required for Winged Eye-mediated transdetermination of Drosophila imaginal disc
Short title
Wge interacts with Lam in TD
Authors
Keita Masuko1, Hirofumi Furuhashi1, Kanae Komaba1, Eriko Numao1, Rumi Nakajima1, Naoyuki
Fuse1, Shoichiro Kurata1*
1Graduate School of Pharmaceutical Sciences, Tohoku University, Sendai 980-8578, Japan
Keywords
Drosophila, imaginal disc, transdetermination, reprogramming, cell fate, Lamin, Winged Eye
Corresponding author’s e-mail address
Abstract
Drosophila imaginal discs often change their cell fate under stress conditions, and this phenomenon, called transdetermination (TD), has long been a useful model for studying cell fate plasticity during regeneration. We previously identified a chromatin-associated protein, Winged Eye (Wge), which induces eye-to-wing TD upon its overexpression in eye imaginal discs. However, the molecular mechanism of Wge-mediated TD remains obscure. Here we analyzed Wge-interacting proteins, and found that several heterochromatin-related proteins, including a nuclear lamina protein, Lamin (Lam), were associated with Wge protein in cultured cells. Knockdown experiments revealed that Lam is indeed required for Wge-mediated eye-to-wing TD. Moreover, Wge-overexpression altered the spatial organization of genomic DNA inside the cell nuclei. Accordingly, we suggest that Wge interacts with Lam to link some genomic regions with the nuclear periphery and regulates chromatin dynamics in imaginal disc TD.
Introduction
The developmental potency of cells in multicellular organisms becomes progressively restricted over time, but throughout the course of development some cells exhibit reprogramming ability in response to various perturbations. Sophisticated regulations of cell fate plasticity are involved in many biological processes, including development, regeneration, and disease processes (Eguchi & Kodama 1993; Thowfeequ et al. 2007), and understanding this regulation could make it possible to apply them for regenerative medicine in the future.
The Drosophila imaginal discs are the larval epithelial tissues that form adult appendages after metamorphosis (e.g., eye disc forms adult eye), and have been an invertebrate model for studying regeneration (Repiso et al. 2011; Worley et al. 2012). Notably, a group of disc cells can acquire different disc identity instead of the original one during regeneration, a phenomenon called transdetermination (TD) (Hadorn 1968; Worley et al. 2012). For example, the cells of the eye disc could transdetermine into cells with wing identity, and consequently, an ectopic wing would form in the adult eye region (eye-to-wing TD; Katsuyama et al. 2005; Masuko et al. 2018; Zhu et al. 2018). Drosophila's powerful genetics enables the study of imaginal disc TD to address fundamental features of cell fate plasticity. However, the molecular mechanism of TD is still largely mysterious. We have studied genetically induced TD using the GAL4/UAS system (Kurata et al. 2000), and previously found that overexpression of Winged Eye (Wge) under the control of an eye disc-specific GAL4 driver (eyeless-GAL4) induced eye-to-wing TD (Katsuyama et al. 2005). We demonstrated that Wge is a heterochromatin-related protein (Ozawa et al. 2016), and overexpression of Wge induces ectopic expression of vestigial (a master regulator of wing identity) in the eye disc via heterochromatin-related histone modifications (Masuko et al. 2018); but we are still far from a comprehensive understanding of the complex TD process.
Here, we analyzed Wge-interacting proteins in cultured cells, and found that several heterochromatin-related proteins, including Lamin (Lam), were co-purified with Wge. Lam is an evolutionally conserved intermediate filament protein that coats the nuclear membrane (Burke & Stewart 2013). The nuclear lamina protein family consists of two classes: A-type lamin is expressed in a subset of differentiated cells, and B-type lamin is expressed ubiquitously in many cells. Lam is the sole B-type lamin in Drosophila (Osouda et al. 2005). Several studies showed that nuclear lamina proteins regulate spatial genome organization and consequently gene expression during cell differentiation (Wen et al. 2009; Kohwi et al. 2013; Solovei et al. 2013; Poleshko et al. 2017). Taking these facts together with our present findings, we propose that Wge interacts with Lam to induce imaginal disc TD via spatial genome reorganization.
Results
Analysis of Wge-interacting proteins
To elucidate the molecular mechanisms of Wge-mediated TD, we sought to identify Wge-interacting proteins (WiPs). For this, Flag-tagged Wge protein was transiently expressed in Drosophila S2 cultured cells, and WiPs were co-immunoprecipitated from the nuclear extract using anti-Flag antibody (Fig. 1A). First, we found that core histone proteins (H2A/H2B/H3/H4) were co-purified with Wge (Fig. S1A), suggesting that Wge possesses chromatin-binding activity. We then performed mass-spectrometric analysis of other WiPs detected as bands on SDS-PAGE (Fig. 1A), and listed candidate WiPs (Table S1). It is interesting that the list included some heterochromatin regulators (Linker histone H1 (His1), Modulo, and Lam), because our previous reports showed that wge affects the heterochromatin-formation and heterochromatin-related histone modifications (Ozawa et al. 2016; Masuko et al. 2018). His1 binds to linker DNA between nucleosomes and functions in gene silencing (Lu et al. 2009); Modulo is classified as a suppressor of position effect variegation (PEV) (Perrin et al. 1998), which involves establishment/maintenance of heterochromatin structure (Elgin & Reuter 2013); Lam is a major component of the nuclear lamina on the inner nuclear membrane, and functions to regulate heterochromatin organization (Burke & Stewart 2013; van Steensel & Belmont 2017). We confirmed Wge-His1, -Modulo, and -Lam interactions in S2 cells through immuno-blots using specific antibodies (Fig. 1B and Fig. S1). Thus, we found that Wge associates with these heterochromatin-related factors in living cells.
Wge interacts with Lam in cells
Among these WiPs, we focus on Lam in this report, because there is increasing evidence that nuclear lamina proteins govern spatial genome organization in cell fate decisions (Wen et al. 2009; Kohwi et al. 2013; Solovei et al. 2013; Poleshko et al. 2017). Furthermore, we recently observed that Wge recombinant protein binds to Lam protein in vitro (Masuko et al. 2018). However, it is still not clear whether Wge interacts with Lam in vivo or what is the function of Lam in the TD process. Accordingly, we examined the relationship between Lam and Wge in cells as follows.
The Wge-Lam interaction in S2 cells was evident from the results of reciprocal immunoprecipitation using antibody against each protein (Fig. 1B, C). Moreover, the Wge-BAH (bromo adjacent homology) domain, which is necessary for the chromatin localization of Wge (Katsuyama et al. 2005), was dispensable for the Wge-Lam protein interaction (Fig. 1D), implying that Wge interacts with Lam and chromatin in independent manners. Thus, Wge protein physically associates with Lam protein in cells as well as in vitro.
Lam is required for Wge-mediated TD
We then investigated whether Lam contributes to Wge-mediated eye-to-wing TD in fly. Eye-specific knockdown (KD) of Lam (ey-GAL4; UAS-Lam KD) caused a slightly roughened shape of the adult eye, but no gross defects in overall eye structure, as previously reported (Fig. 2B; Patterson et al. 2004; Osouda et al. 2005). In the Wge-overexpression condition (ey>wge, GFP KD), about 16% of flies showed the TD phenotype and formed ectopic wing structures in eye regions (Fig. 2D). Moreover, the flies with both Wge-overexpression and Lam KD (ey>wge, Lam KD) showed significantly reduced eye-to-wing TD frequency and showed mostly small eyes (Fig. 2E and 2G), suggesting that Lam contributes to Wge-mediated TD.
We recently reported that overexpressed Wge protein in fly cells was not uniformly distributed in the nuclei, but rather was enriched at the nuclear periphery (NP), particularly at contact sites between chromatin and the NP (Masuko et al. 2018, see also Fig. S2A), implying that Lam might tether Wge protein to the NP. We examined the effects of Lam KD on the localization of Wge protein, and observed that its localization at the NP was not disturbed in the Lam KD condition (Fig. S2B and S2C). Therefore, we speculate that other factor(s) in addition to Lam might also contribute to Wge localization at the NP. Lamin C (the sole A-type lamin protein in Drosophila; Reimer et al. 1995) is a candidate of such factors.
Since ey>wge, LamC KD flies suffered tissue collapse during development, probably due to secondary effects (data not shown), we could not address the relationships of Wge and LamC. As an alternative way, we decided to use transgenic flies carrying UAS-Lam∆CAAX, which lacks the
sequence coding for the C-terminal four amino acids of Lam, resulting in defective nuclear membrane targeting of the protein (Kitten & Nigg 1991; Pickersgill et al. 2006; Uchino et al. 2017). We expected that Lam∆CAAX protein might interfere with the Wge localization at the NP. Consistent
with previous reports, Lam∆CAAX protein was distributed in the nucleoplasm rather than at the NP when
overexpressed by ey-GAL4 (Fig. S3A). We found that the expression of Lam∆CAAX clearly attenuated
the enrichment of Wge at the NP (Fig. S3B and S3C), suggesting that Lam∆CAAX antagonizes the
Wge-NP interaction. In this condition (ey>wge, Lam∆CAAX), the eye-to-wing TD frequency was
markedly reduced, to 2.8% (Fig. 2H), and reduced eye size was observed again (Fig. 2F). Accordingly, we concluded that the NP-localized Lam is required for Wge-mediated TD. Overexpression of Lam∆CAAX alone did not affect eye development (Fig. 2C), and Lam null mutant fly
has no obvious defect of wing specification (Osouda et al., 2005), suggesting that Lam contributes to switching the disc identity in Wge-mediated TD, but not to normal development of adult appendages.
The above results suggest that there are interactions among chromatin, Wge, Lam and the NP in cell nuclei. Thus, we hypothesized that topological chromatin dynamics might be changed in the process of Wge-mediated TD. Recent reports suggest that gene loci move to the NP and back to the interior of the nucleus during cell differentiation in context-dependent manners, and that the proximity of a gene locus to the NP is correlated to the suppression of its transcription (Kohwi et al. 2013; Poleshko et al. 2017). To examine whether this kind of regulation operates in Wge-mediated TD, we performed Immuno-FISH (immunohistochemistry combined with fluorescence in situ hybridization) experiments to analyze the spatial genome organization in eye disc cells (Harmon & Sedat 2005). We prepared two fluorescence-labeled DNA probes, one against the Histone Cluster (His-C) locus and the other against the eyes absent (eya) locus. We adopted the His-C probe as a control because this region is actively transcribed in most cells. On the other hand, eya is specifically expressed in the eye disc (Bonini et al. 1993), and is suppressed by Wge-overexpression in a cell-autonomous manner (Katsuyama et al. 2005).
First, we confirmed by FISH that each of these probes correctly marked a single gene locus on a polytene chromosome (Fig. 3A and 3B). Then, we performed immuno-FISH using wild-type eye-antennal discs. We detected signals for each FISH as dot(s) in cell nuclei (up to two dots per cell; Fig. 3D to 3G), and measured the distances of these FISH dots from the NP (see Experimental procedures). Consistent with the fact that eya is expressed in eye disc cells, but not in antennal disc cells, we found that eya dots were more frequently close to the NP in antennal disc cells compared to eye disc cells (Fig. 3E, 3G & 3H). In contrast, the positions of His-C dots were not strikingly different in these two cell types (Fig. 3D, 3F & 3H). These results suggest that the position of the eya locus in nuclei is differentially regulated in these different cell lineages. Next, we generated Wge-overexpressing clones in eye discs (Fig. 3C), and measured the distances of FISH dots from the NP inside Wge-expressing clones (labeled with GFP) or outside of the clones (wild-type cells with no GFP). The results showed that eya dots were more frequently located close to the NP in Wge-expressing clones than in the wild-type cells (Fig. 3J, 3L & 3M). In contrast, the positions of His-C dots were not strikingly affected (Fig. 3I, 3K & 3M). This suggests that Wge-overexpression affects the spatial genome organization of the eya locus in eye disc cells.
Discussion
Here we advanced our understanding of the molecular action of the TD-inducer Wge, and identified some chromatin regulators acting together with Wge. We previously reported that Wge protein associates with two proteins, Mute (Muscle wasted) and Su(var)3-9 (Suppressor of variegation 3-9) (Ozawa et al. 2016; Masuko et al. 2018). Regarding the former, endogenous Wge and Mute are components of the nuclear body called histone locus body (HLB) (Ozawa et al. 2016), which is involved in gene expression and pre-mRNA metabolism at the histone gene cluster (Duronio & Marzluff 2017). However, it is unclear whether HLB is involved in Wge-mediated TD, because overexpressed Wge protein associates with many chromosomal loci, not only with HLB (Katsuyama et al. 2005). As for the latter of these associated proteins, Su(var)3-9 is a histone methyltransferase specific to histone H3 at lysine 9 (H3K9), and is important for regulation of heterochromatin organization (Elgin & Reuter 2013). Overexpression of Wge enhances H3K9 methylation, and Su(var)3-9 is required for Wge-mediated H3K9 methylation and TD (Masuko et al. 2018). In the present study, we identified three other WiPs (Lam, His1 and Modulo), which are involved in heterochromatin regulations. The his1 and modulo genes, as well as the wge gene, are classified as “suppressors of PEV” (Perrin et al. 1998; Lu et al. 2009), which are generally involved in heterochromatin organization (Elgin & Reuter 2013); and His1 plays a key role in gene silencing and recruitment of Su(var)3-9 to chromatin (Lu et al. 2013). Further analysis is needed, however, to investigate how these WiPs contribute to Wge-mediated TD.
We showed here that Lam is one of the WiPs acting in TD. Since Lam null mutant flies have been reported to be normal-looking adults (Osouda et al. 2005), our observations suggest that Lam might be involved in switching of disc identity rather than disc identity per se. How does Lam contribute to Wge-mediated TD? Wge-overexpression caused eya to be transcriptionally silenced, and changed the position of the eya locus such that it was closer to the NP. Wge and Lam were localized to the NP in cells. Overexpression of Lam∆CAAX attenuated those localizations and the Wge-mediated TD. From
these results, we hypothesize that Lam might govern the spatial genome organization of Wge-binding chromatin domains, and those genome dynamics might shift the gene expression mode so as to induce TD. Although Lam KD dampened Wge-mediated TD, it did not visibly attenuate Wge localization at the NP. Regarding this discrepancy, we found that Lam KD significantly impaired the Wge-mediated increase of H3K9me2 (Fig. S2D and S2E). This suggests that Lam might be involved in the heterochromatin-related histone modifications, as well as in genome dynamics, under Wge-overexpression. Further analysis will be required to understand the relationships of genome dynamics and chromatin modifications in the TD process. Lam is known to
contribute to genome dynamics during Drosophila neuroblast development (Kohwi et al. 2013). Although we attempted to investigate the involvement of Lam in repositioning of the eya locus in Wge-overexpressing clones, it was unfortunately difficult to examine it, because Wge-overexpressing clone in the absence of Lam activity were rarely detected in the eye disc, or even if detected, showed abnormal cell architecture (Fig. S4). This might suggest that Lam is also required for maintaining cell integrity under Wge-overexpression.
There is increasing evidence that NP-mediated spatial genome organization seems to be linked to the process of cell fate decision (Wen et al. 2009; Kohwi et al. 2013; Solovei et al. 2013; Poleshko et al. 2017). Moreover, C. elegans perinuclear protein CEC-4 anchors methylated H3K9 at the NP, and this chromatin anchoring ensures full commitment in the process of transdifferentiation via forced expression of HLH-1 (MyoD orthologue, a master regulator of muscle differentiation), although how CEC-4 contributes to the transdifferentiation process is not known (Gonzalez-Sandoval et al. 2015). We speculate that spatial genome reorganization of large-scale domains via heterochromatin regulation is an important step of Wge-mediated TD. To address how chromatin-NP interaction organizes the process of TD, DamID-seq (DNA adenine methyltransferase identification-followed by high throughput sequencing; Pickersgill et al. 2006; Southall et al. 2013) would be a powerful method, which would enable us to characterize the Lamin-chromatin interaction domains at the genome scale in the TD context. Further analysis will shed light on how chromatin organization functions in in vivo cellular plasticity in the future.
Experimental procedures
Fly stocks and genetics
Flies were raised on a standard cornmeal medium. The following alleles and transgenic lines were used in this study. ey-GAL4 (Kurata et al. 2000), UAS-HA-wge (Katsuyama et al. 2005), UAS-Lam KD (VDRC #107419), UAS-LamC KD (Bloomington #31621), UAS-Flag-Lam∆CAAX (generated in
this work), Ay-GAL4, hs-flp. Unless otherwise indicated, the fly lines were obtained from the Bloomington Drosophila Stock Center, the Vienna Drosophila RNAi Center (VDRC), or the Kyoto Stock Center (DGRC). Wge-expressing clones were induced by heat-shock treatment (37˚C for 1 hr) at 48 hr after egg laying (AEL) of the following flies: yw hs-flp; Ay-GAL4 UAS-GFP/+; UAS-wge/+. To generate the UAS-Lam∆CAAX transgenic fly, Lam∆CAAX cDNA was made by standard
PCR from a female adult fly cDNA, and cloned into pUAST vector. The following primers were used to generate Lam∆CAAX cDNA:
Lam fw; cggaattcatgtcgagcaaatcccgacg,
Lam∆CAAX rv; ataagaatgcgccggcgttacttctcgtttgactgctg
Cell culture and cloning
S2 cells were cultured in Schneider’s medium (GIBCO) with 10% fetal calf serum (MP Biomedicals, Fisher Scientific) at 25˚C.
To generate Flag-wge/Flag-wge∆BAH -inducible cell lines, wge/wge∆BAH sequences (Katsuyama et al. 2005) were cloned into pMT-Flag-SBP vector, which was generated from pMT/V5-HisA vector (Thermo Fisher Scientific) by adding Flag and SBP (streptavidin-binding peptide)-tag. To investigate the Wge-Modulo protein interactions, we constructed a V5-Modulo/V5-GFP expression plasmid. Modulo/GFP cDNAs were made by PCR from the female adult fly cDNA and pACGFP1-Hyg-N1 vector (Clontech), respectively, and were cloned into pMT/V5-HisA vector (Thermo Fisher Scientific).
The following primers were used to generate modulo and GFP cDNA: Modulo fw;ggactagtaaatggcccaaaagaaagccgtg,
Modulo rv; gccgctcgagaaatcttgcccttttaacaaac GFP fw; ggactagtatggtgagcaagggcgccga GFP rv; gccgctcgagcttgtacagctcatccatgc
Immunoprecipitation
Affinity purification of Wge-interacting proteins was performed as previously described (Ozawa et al. 2016). 1x106 cells/mL (pMT-Flag-wge/S2 or pMT-Flag-wge∆BAH/S2 stable cell lines) were
inoculated in 30 mL flasks, and after a 24 hr incubation, CuSO4 was added at a final concentration of
100 µM. Nuclear extracts of pMT-Flag-wge/S2 or pMT-Flag-wge∆BAH/S2 (CuSO4 or mock-treated
for 12 hr) were used as the starting material for immunoprecipitation. When we prepared nuclear extracts, 10 units of DNase (Promega) were added to lyse insoluble fractions. Dialyzed nuclear extracts were incubated with antibodies and ethidium bromide (Lai & Herr 1992) for 2 hr at 4˚C, and subsequently incubated with pre-cleared Dynabeads M280 (Life Technologies) for 1 hr at 4˚C. Captured protein complexes were eluted by incubation with elution buffer (300 µg/mL Flag peptides; 1 x Reducing agent, Life Technologies) at 70˚C twice. The antibodies used were as follows: mouse anti-Flag mAb (M2, 1 µg, Sigma); mouse anti-Lam mAb (ADL84.12, 1 µg, DSHB); rabbit anti-His1 pAb (0.3 µL, a gift from Dr. James Kadonaga; University of California San Diego). Immuno-precipitates were analyzed by SDS-PAGE and subsequent silver-staining, CBB-staining, and immuno-blotting. WiPs were characterized by LC-MS/MS performed in APROSCIENCE Co., Ltd.
Histochemistry
Preparation of polytene chromosome squashes and FISH-combined with immunostaining of imaginal disc and salivary glands were performed as previously described (Grimaud et al. 2006; Masuko et al. 2018). The primary antibodies used were as follows: mouse anti-Flag mAb (M2, 1:200, Sigma); rat anti-HA mAb (3F10, 1:500, Roche); rabbit anti-GFP pAb (596, 1:1,000, MBL); mouse anti-Lam mAb (ADL84.12, 1:100, DSHB); rabbit anti-Wge pAb (1:100, Ozawa et al. 2016). Fluorescently labeled DNA probes for the His-C locus and eya locus were synthesized using a FISH Tag DNA Multicolor kit (Thermo Fisher Scientific). Genomic DNA was extracted from a female adult, and used as template DNA. We made DNA fragments of the His-C and eya loci by standard PCR, and synthesized amine-modified DNA fragments using a nick-translation system. We then labeled the amine-modified DNA with fluorescent dye, and purified labeled DNA by ethanol precipitation. The eya locus probe used consisted of a mixture of 4 types of probes that covered the eya coding region.
The following primers were used to generate FISH probes: His-C fw; cacttcaagcaaacttcgac His-C rv; ccgattatagagtacgctag eya1 fw; ttctacgatctggccgttct eya1 rv; atgcgcataggaaaaccaac eya2 fw; cctgactgtccggaataagc eya2 rv; ccctcgctgagaaaactcac
eya3 fw; gttcgatggatccgtaacgc eya3 rv; gacatcttgagaataggtcaac eya4 fw; gaagcgataatgccaggtgt eya4 rv; tgcccatgaaagggttaaag
Image analysis for DNA-immuno FISH
All imaging was performed using a Lieca Confocal Microscopy system equipped with LAS AF software (Leica Microsystems) and a digital color camera (Leica DFC310 FX, Leica Microsystems). FISH imaginal discs were imaged using an ACS APO 63x/1.30 OIL CS objective lens with 2.5x optical zoom. We used independent imaginal discs for analyzing eya signal and His-C signal. Pinholes were adjusted to equalize the resolution of all wavelengths. Therefore, rim-like Lam signals and FISH dot signals in an optical cross section would be within the same focal plane. To measure gene position within the nucleus, we selected the analyzable cells, which had a clear FISH signal, rim-like Lam signal and appropriate nuclear size. Then, we measured the shortest distance from the center of the FISH signal to the edge of the nucleus using ImageJ. The statistical differences of NP-FISH signal distances were tested by the chi-square test and the Wilcoxon-Mann-Whitney rank sum test on the R platform.
Western blotting
To obtain salivary gland tissue lysates, 10 pairs of salivary glands from wandering 3rd instar larvae were dissected and homogenized in 100 µL of lysate buffer (1xLDS and 1xReducing agent, Life Technologies), and incubated at 70˚C for 10 min. Western blotting was performed using the NuPAGE system (Life Technologies). The protein-transferred polyvinylidene difluoride membranes were incubated with blocking buffer (PBS, 0.05% Tween 20, 3% skim milk, overnight, 4˚C), primary antibodies (as indicated below, 2 hr, RT), secondary antibodies (conjugated with HRP, GE Healthcare, 1 hr, RT), and immuno-labelled proteins were detected using Luminata Forte Western HRP substrate (Millipore). Images were acquired using a LAS-4000UVmini (GE Healthcare). The primary antibodies used were as follows: mouse anti-Flag monoclonal Ab (1E6; 1:5,000; Wako), mouse anti-Lam monoclonal Ab (ADL67.10; 1:5,000; DSHB), mouse anti-LamC monoclonal Ab (LC28.26; 1:1000; DSHB), rabbit anti-His3 polyclonal Ab (ab1791; 1:10,000; abcam), rabbit anti-His1 polyclonal Ab (a gift from Dr. James Kadonaga; University of California San Diego), and mouse anti-V5 monoclonal Ab (R960-25; 1:1,000; Invitrogen).
Observation of adult eye phenotype
the morphology of compound eyes using a stereoscopic microscope (Stemi 2000, Zeiss). The eye phenotypes were categorized as described in a previous report (Masuko et al. 2018).
Acknowledgements
We thank Dr. James Kadonaga for α-His1 antibodies, and Drs. Susumu Hirose and Kiyoe Ura for helpful suggestions and discussions. We also thank the Bloomington Drosophila Stock Center at Indiana University, Kyoto Stock Center (DGRC), and the Vienna Drosophila RNAi Center for fly stocks. We also thank the S.K. laboratory members for discussions and helpful comments on the manuscript. The authors declare that there are no conflicts of interest. This work was supported by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (MEXT 25670019, 20052004, 18055003, 18016001, 14380343, 13202004, 12202009, 12026202, 11152201, 11142203, 10480207, and 10171206); the Japan Society for the Promotion of Science (JSPS); Japan Science and Technology Agency (JST); the Mitsubishi Foundation; and Sumitomo Foundation.
References
Bonini, N.M., Leiserson, W.M., & Benzer, S. (1993). The eyes absent gene: genetic control of cell survival and differentiation in the developing Drosophila eye. Cell 72, 379–395.
Burke, B., & Stewart, C.L. (2013). The nuclear lamins: flexibility in function. Nat. Rev. Mol. Cell Biol. 14, 13–24.
Duronio, R.J., & Marzluff, W.F. (2017). Coordinating cell cycle-regulated histone gene expression through assembly and function of the Histone Locus Body. RNA Biol 14, 726–738.
Eguchi, G., & Kodama, R. (1993). Transdifferentiation. Curr. Opin. Cell Biol. 5, 1023–1028. Elgin, S.C., & Reuter, G. (2013). Position-effect variegation, heterochromatin formation, and gene silencing in Drosophila. Cold Spring Harb Perspect Biol 5, a017780.
Filion, G.J., van Bemmel, J.G., Braunschweig, U., Talhout, W., Kind, J., Ward, L.D., ... van Steensel, B. (2010). Systematic protein location mapping reveals five principal chromatin types in Drosophila cells.Cell 143, 212–224.
Gabut, M., Dejardin, J., Tazi, J., & Soret, J. (2007). The SR family proteins B52 and dASF/SF2 modulate development of the Drosophila visual system by regulating specific RNA targets. Mol. Cell. Biol. 27, 3087–3097.
Garzino, V., Pereira, A., Laurenti, P., Graba, Y., Levis, R.W., Le Parco, Y., & Pradel, J. (1992). Cell lineage-specific expression of modulo, a dose-dependent modifier of variegation in
Drosophila. EMBO J. 11, 4471–4479.
Gonzalez-Sandoval, A., Towbin, B.D., Kalck, V., Cabianca, D.S., Gaidatzis, D., Hauer, M.H., ... Gasser, S.M. (2015). Perinuclear Anchoring of H3K9-Methylated Chromatin Stabilizes Induced Cell Fate in C. elegans Embryos. Cell 163, 1333–1347.
Grimaud, C., Bantignies, F., Pal-Bhadra, M., Ghana, P., Bhadra, U., & Cavalli, G. (2006). RNAi components are required for nuclear clustering of Polycomb group response elements. Cell 124, 957–971.
Guelen, L., Pagie, L., Brasset, E., Meuleman, W., Faza, M.B., Talhout, W., ... van Steensel, B. (2008). Domain organization of human chromosomes revealed by mapping of nuclear lamina interactions. Nature 453, 948–951.
Hadorn, E. (1968). Transdetermination in cells. Sci. Am.219, 110–4 passim.
Harmon, B., & Sedat, J. (2005). Cell-by-cell dissection of gene expression and chromosomal interactions reveals consequences of nuclear reorganization. PLoS Biol. 3, e67.
Isaac, C., Yang, Y., & Meier, U.T. (1998). Nopp140 functions as a molecular link between the nucleolus and the coiled bodies. J. Cell Biol. 142, 319–329.
Katsuyama, T., Sugawara, T., Tatsumi, M., Oshima, Y., Gehring, W.J., Aigaki, T., & Kurata, S. (2005). Involvement of winged eye encoding a chromatin-associated bromo-adjacent homology domain protein in disc specification. Proc. Natl. Acad. Sci. U.S.A. 102, 15918–15923.
Kitten, G.T., & Nigg, E.A. (1991). The CaaX motif is required for isoprenylation, carboxyl methylation, and nuclear membrane association of lamin B2. J. Cell Biol.113, 13–23. Kohwi, M., Lupton, J.R., Lai, S.-L.L., Miller, M.R., & Doe, C.Q. (2013). Developmentally regulated subnuclear genome reorganization restricts neural progenitor competence in Drosophila. Cell 152, 97–108.
Kurata, S., Go, M.J., Artavanis-Tsakonas, S., & Gehring, W.J. (2000). Notch signaling and the determination of appendage identity. Proc. Natl. Acad. Sci. U.S.A. 97, 2117–2122.
Lai, J.S., & Herr, W. (1992). Ethidium bromide provides a simple tool for identifying genuine DNA-independent protein associations. Proc. Natl. Acad. Sci. U.S.A. 89, 6958–6962.
Lu, X., Wontakal, S.N., Emelyanov, A.V., Morcillo, P., Konev, A.Y., Fyodorov, D.V., & Skoultchi, A.I. (2009). Linker histone H1 is essential for Drosophila development, the establishment of
pericentric heterochromatin, and a normal polytene chromosome structure. Genes Dev. 23, 452– 465.
Lu, X., Wontakal, S.N., Kavi, H., Kim, B.J., Guzzardo, P.M., Emelyanov, A.V., ... Skoultchi, A.I. (2013). Drosophila H1 regulates the genetic activity of heterochromatin by recruitment of
Su(var)3-9. Science 340, 78–81.
Masuko, K., Fuse, N., Komaba, K., Katsuyama, T., Nakajima, R., Furuhashi, H., & Kurata, S. (2018). winged eye Induces Transdetermination of Drosophila Imaginal Disc by Acting in Concert with a Histone Methyltransferase, Su(var)3-9. Cell Reports 22, 206–217
Meier, U.T. (2005). The many facets of H/ACA ribonucleoproteins. Chromosoma 114, 1–14. Ni, J.-Q.Q., Liu, L.-P.P., Hess, D., Rietdorf, J., & Sun, F.-L.L. (2006). Drosophila ribosomal proteins are associated with linker histone H1 and suppress gene transcription. Genes Dev. 20, 1959–1973.
Osouda, S., Nakamura, Y., de Saint Phalle, B., McConnell, M., Horigome, T., Sugiyama, S., ... Furukawa, K. (2005). Null mutants of Drosophila B-type lamin Dm(0) show aberrant tissue differentiation rather than obvious nuclear shape distortion or specific defects during cell proliferation. Dev. Biol. 284, 219–232.
Ozawa, N., Furuhashi, H., Masuko, K., Numao, E., Makino, T., Yano, T., & Kurata, S. (2016). Organ identity specification factor WGE localizes to the histone locus body and regulates histone expression to ensure genomic stability in Drosophila. Genes Cells 21, 442–456.
functions of Klarsicht and nuclear lamin in developmentally regulated nuclear migrations of photoreceptor cells in the Drosophila eye. Mol. Biol. Cell 15, 600–610.
Perrin, L., Demakova, O., Fanti, L., Kallenbach, S., Saingery, S., Mal’ceva, N.I., ... Pradel, J. (1998). Dynamics of the sub-nuclear distribution of Modulo and the regulation of position-effect variegation by nucleolus in Drosophila. J. Cell. Sci.111 ( Pt 18), 2753–2761.
Pickersgill, H., Kalverda, B., de Wit, E., Talhout, W., Fornerod, M., & van Steensel, B. (2006). Characterization of the Drosophila melanogaster genome at the nuclear lamina. Nat. Genet. 38, 1005–1014.
Poleshko, A., Shah, P.P., Gupta, M., Babu, A., Morley, M.P., Manderfield, L.J., ... Jain, R. (2017). Genome-Nuclear Lamina Interactions Regulate Cardiac Stem Cell Lineage Restriction. Cell 171, 573–587.e14.
Reddy, K.L., Zullo, J.M., Bertolino, E., & Singh, H. (2008). Transcriptional repression mediated by repositioning of genes to the nuclear lamina. Nature 452, 243–247.
Repiso, A., Bergantiños, C., Corominas, M., & Serras, F. (2011). Tissue repair and regeneration in Drosophila imaginal discs. Dev. Growth Differ. 53, 177–185.
Riemer, D., Stuurman, N., Berrios, M., Hunter, C., Fisher, P.A., & Weber, K. (1995). Expression of Drosophila lamin C is developmentally regulated: analogies with vertebrate A-type lamins. J. Cell. Sci. 108 ( Pt 10), 3189–3198.
Solovei, I., Wang, A.S., Thanisch, K., Schmidt, C.S., Krebs, S., Zwerger, M., ... Joffe, B. (2013). LBR and lamin A/C sequentially tether peripheral heterochromatin and inversely regulate
differentiation. Cell 152, 584–598.
Southall, T.D., Gold, K.S., Egger, B., Davidson, C.M., Caygill, E.E., Marshall, O.J., & Brand, A.H. (2013). Cell-type-specific profiling of gene expression and chromatin binding without cell isolation: assaying RNA Pol II occupancy in neural stem cells. Dev. Cell 26, 101–112.
Thowfeequ, S., Myatt, E.-J.J., & Tosh, D. (2007). Transdifferentiation in developmental biology, disease, and in therapy. Dev. Dyn. 236, 3208–3217.
Uchino, R., Sugiyama, S., Katagiri, M., Chuman, Y., & Furukawa, K. (2017). Non-farnesylated B-type lamin can tether chromatin inside the nucleus and its chromatin interaction requires the Ig-fold region. Chromosoma 126, 125–144.
Van Steensel, B., & Belmont, A.S. (2017). Lamina-Associated Domains: Links with Chromosome Architecture, Heterochromatin, and Gene Repression. Cell 169, 780–791.
Wang, C.I., Alekseyenko, A.A., LeRoy, G., Elia, A.E., Gorchakov, A.A., Britton, L.-M.P.M., ... Kuroda, M.I. (2013). Chromatin proteins captured by ChIP-mass spectrometry are linked to dosage compensation in Drosophila. Nat. Struct. Mol. Biol. 20, 202–209.
Wen, B., Wu, H., Shinkai, Y., Irizarry, R.A., & Feinberg, A.P. (2009). Large histone H3 lysine 9 dimethylated chromatin blocks distinguish differentiated from embryonic stem cells. Nat.
Genet. 41, 246–250.
Worley, M.I., Setiawan, L., & Hariharan, I.K. (2012). Regeneration and transdetermination in Drosophila imaginal discs. Annu. Rev. Genet. 46, 289–310.
Zhu, J., Ordway, A., Weber, L., Buddika, K., & Kumar, J.P. (2018). Polycomb group (Pc-G) proteins and Pax6 cooperate to inhibit in vivo reprogramming of the developing Drosophila eye. Development, doi: 10.1242/dev.160754,
Figure legends
Figure 1. Wge interacts physically with Lam in living cells.
(A) Flag-Wge was induced in S2 cultured cells in the presence of CuSO4. Nuclear extracts (Input)
were immunoprecipitated using anti-Flag antibody (Flag-IP) and analyzed by silver-staining. The specific bands (marked 1-6*) were analyzed by mass spectrometry. (B and C) Western blots of Flag-IP fractions (B) and Lam-IP fractions (C) showed Wge-Lam interaction in cultured cells. (D) Full-length Wge or BAH domain-deleted Wge (∆BAH) was expressed in S2 cells, and western blotting of Flag-IP fractions was performed.
Figure 2. Lam is required for Wge-mediated TD.
(A to C) Representative eye phenotype of eye-specific Lam-KD or Lam∆CAAX overexpression flies.
These flies did not show obvious defects in eye formation. (D to F) Representative eye phenotype of the flies indicated in (G and H). Arrow indicates ectopic wing structure. ey>wge, Lam-KD flies and ey>wge, Lam∆CAAX flies mostly showed small eyes. (G and H) Eye-to-wing TD frequency in adult
flies. Ratios of the number of flies with an ectopic wing in the eye field relative to the total number of flies are indicated. UAS-GFP-KD and UAS-GFP were used as control crosses. *P<0.05 by chi-square test.
Figure 3. Wge-overexpression affects spatial genome organization of eya locus in eye disc.
(A and B) FISH signal on polytene chromosomes showed correct targeting of the probes against the His-C locus (A) and eya locus (B). DAPI (cyan) and FISH signals (red) are merged. Inset boxes show higher-magnification views. (C) An example of mosaic analysis of Wge-overexpression in eye-antennal disc. GFP signals indicate Wge-overexpressing clones. (D to G) Immuno-FISH of eye-antennal disc in wild-type flies (Oregon-R). DAPI (blue), Lam (green) and FISH signals (red; His-C locus in (D and F); eya locus in (E and G)) were merged. Eye disc cells (D and E) and antennal disc cells (F and G) are shown. eya dots were more frequently located close to the NP in antennal disc cells (G, white arrow). (H) Percentile plots of distances between FISH signals and the NP are shown. Y-axis shows cumulative frequency of FISH signals. Statistical test results are shown in Table S2. (I to L) Immuno-FISH of eye disc in Wge-overexpression mosaic flies (hs-Flp; Ay>GFP, wge). DAPI (blue), Lam (green) and FISH signals (red; His-C locus in (I and K); eya locus in (J and L)) were merged. Clone-outside cells (control cells; I and J) and clone-inside cells (Wge-overexpression; K and L) are shown. eya dots were more frequently located close to the NP in clone-inside cells (L, white arrow). (M) Percentile plots of distances between FISH signals and the
M
290 240 160 116 97 66 55 40-+
-+
Input Flag-IP Flag-Wge InductionA
B
-+
Input Flag-IP+
-Flag-Wge LamC
Input Lam-IP IgG-IP
Flag-Wge Lam
D
Lam Flag-Wge ∆BAHFull Full∆BAH
Input Flag-IP 5* 6* 3* 2* 4* 1* Flag-Wge Induction
eye to wing transformation (%) 5 10 15 0 16.1% (41/254) 20 6.56% (16/244) Lam-KD * ey>wge GFP-KD
eye to wing transformation (%)
5 10 15 0 16.3% (17/104) 20 2.80% (4/143) UAS-Lam ΔCAAX * ey>wge UAS-GFP
A
B
C
D
E
F
G
H
ey -GAL4 ey -GAL4; UAS-wgeLam KD UAS-Lam∆CAAX
+
Lam KD UAS-Lam∆CAAX
0 1.0 2.0 distance from nuclear periphery [µm] 20
40 60 80 100
cumulative cell number [%]
eya probe in antennal disc eya probe in eye disc His-C probe in antennal disc His-C probe in eye disc
0 1.0 2.0 100 80 60 40 20
distance from nuclear periphery [µm]
cumulative cell number [%]
n = 607 n = 1007 n = 342 n = 552
His-C probe inside Wge clone His-C probe outside Wge clone eya probe inside Wge clone eya probe outside Wge clone
FISH / Lam / DAPI
eye disc eya locus probe His-C probe antennal disc clone-outside clone-inside eya locus probe His-C probe eye disc antennal disc FISH / DAPI eya locus probe His-C probe hs-Flp, Ay>GFP, wge GFP/ DAPI n = 370 n = 772 n = 542 n = 448
A
B
C
D
E
F
G
H
I
J
K
L
M
Supplemental Figure 1.
Figure S1. Western blot analyses of Wge-His1 and Wge-Modulo interactions.
(A) Flag-IP fractions from pMT-Flag-Wge/S2 cells were analyzed by CBB-staining. Four bands in the Flag-IP fraction are characteristic of core histone proteins. +/- indicates Flag-Wge induction or lack of induction. (B and C) Western blots of immunoprecipitates of Flag-IP fractions (B) and His1-IP fractions (C) showed Wge-His1 interaction in cultured cells. (C) Western blots of Flag-IP fraction from S2 cells expressing Flag-Wge/V5-Modulo, Flag-Wge/V5-GFP, and V5-Modulo. V5-GFP was used as a negative control for immunoprecipitation.
Input
IgG-IP
His1-IP
Flag-Wge
His1
Flag-Wge
V5-Modulo
V5-GFP
Input
Flag-IP
Input
Flag-IP
Input
Flag-IP
Flag-Wge
V5-Modulo/S2
Flag-Wge
V5-GFP/S2V5-Modulo/S2
A
B
C
20
15
10
M
+
-
+
-Input Flag-IP
His3
His1
-+
-+
Input
Flag-IP
D
Flag-Wge Induction Flag-Wge InductionSupplemental Figure 2.
Figure S2. Overexpressed-Wge is enriched at contact sites between chromatin and nuclear periphery.
(A and A’) Immunostaining of salivary gland cell for ey>wge larvae. (A’) Higher magnification view of white box in (A). DAPI (blue), Wge (green) and Lam (red) signals are merged. Wge signals were mainly detected on polytene chromosomes, but were also enriched at the NP. (B and C) Immunostaining of larval salivary gland tissue for ey>wge (B) and ey>wge, Lam-KD (C). NP localization of Wge was observed even in the case of knockdown of Lam. (D) Western blot of larval salivary gland tissue lysates showed that Lam-KD impaired the Wge-mediated increase of H3K9me2. (E) Quantifications of (D). Fold changes of relative signal intensities, when control signal is taken as 1, are shown. Data indicate mean SD of three independent experiments. *p < 0.05, by Student's t test. DAPI Wge ey > wge ey > wge, Lam KD Lam ey > wge DAPI Wge Lam
A
D
MergeA’
B
C
HA-Wge Lam H3K9me2 pan-H3 ey> GFP KD UAS-wge ey> Lam KD UAS-wge + + 0 4 2 6Relative H3K9me2 level
ey> GFP KD UAS-wge ey> Lam KD UAS-wge + +
E
*
Supplemental Figure 3.
Figure S3. Lam∆CAAX
expression antagonized nuclear periphery localization of Wge.
Immunostaining of salivary gland tissue for ey>Lam∆CAAX
(A), ey>wge, GFP (B) and ey>wge,
Lam∆CAAX
(C). White boxes show higher magnification views. Wge protein and Lam∆CAAX
protein were detected using anti-HA antibody and anti-Flag antibody, respectively.
ey
>
Lam
∆CAAXey
>
wge, GFP
ey
>
wge, Lam
∆CAAXDAPI
DAPI
Wge
Lam
∆CAAXDAPI
DAPI
Wge
Lam
∆CAAXDAPI
DAPI
Wge
Lam
∆CAAXA
B
C
Supplemental Figure 4.
Figure S4. Wge-overexpressing eye disc clones in the absence of Lam activity were rarely observed, or showed abnormal cell architecture.
(A and B) Immunostaining of eye disc for hs-Flp; Ay>GFP, wge, Lam-KD. GFP signals indicate Wge overexpression clones. Imaginal disc is composed of two types of epithelial monolayer: squamous peripodial membrane (PM) and columnar disc proper. Clones with co-expression of Wge and Lam-KD were prone to being dislocated from disc layer abnormally. (B) Cross section of white dotted line in (A’) is shown. DAPI (cyan) and Phalloidin (red) signals are merged. White lines indicate clones with co-expression of Wge and Lam-KD. Confocal section (CS) roughly corresponds to X-Y plane in (A). Scale Bar, 7.5 µm. (C and D) Immunostaining of eye disc for hs-Flp; Ay>GFP,
wge, UAS-GFP(C) and hs-Flp; Ay>GFP, wge, UAS-Lam∆CAAX
(D). We observed very few clones co-expressing Wge and Lam∆CAAX
in eye disc.
Lamin GFP DAPI
MERGE
X-Y plane X-Z plane cross section
GFP : UAS-wge , UAS-lam RNAi PM CS
A
A’
A”
B
B’
B”
B’’’
DAPI GFP LaminC
C’
D
D’
GFP : UAS-wge GFP : UAS-wge, UAS -lam ∆CAAXSupplemental Table 1.
Table S1. Candidates of Wge-interacting proteins (WiPs).
The list of WiP candidates identified from mass spectrometric analyses of Flag-IP fractions. Protein name, signal band# (see Figure 1A), expected molecular weight (kDa), and protein function/domain are shown.
Protein name signal band Expected MW Functions, domain Reference
Nopp140, Isoform B #2 70.6 k nucleolar and cajal body protein, chaperone for snoRNPs Isaac et al. 1998, Meier 2005 CG8939 #2 93.3 k rRNA methyltransferase activity Fly base
Lamin Dm0 #4 71.3 k nuclear lamina component, heterochromatin, gene silencing Burke & Stewart 2013, Steensel & Belmont 2017 Modulo #4 60.3 k localize to nucleolus and chromatin, suppressor of PEV Garzino et al. 1992, Perrin et al. 1998
CG4747 #4 65.3 k PWWP domain, H3K36me3 binding, dosage compensation Wang et al. 2013
Histone H1 #5 26.3 k Linker histone, heterochromatin, gene silencing Lu et al. 2009, Lu et al. 2013 SF2/ASF #6 28.3 k regulation of alternative mRNA splicing Gabut et al. 2007
Copia protein #6 162.7 k Zinc finger (CCHC-type), Ribonuclease H-like Ni et al. 2006 RpS4 #6 29.1 k Ribosomal protein S4 Ni et al. 2006 RpS6, Isoform B #6 28.4 k Ribosomal protein S6 Ni et al. 2006 RpS8e (intronic protein 259) #6 23.8 k Ribosomal protein S8e Ni et al. 2006 CG13096 #3 74.3 k Ribosomal protein L1-like homologue Fly base RpL5, Isoform A #5 34 k Ribosomal protein L5 Fly base RpL6 Isoform B #6 29.7 k Ribosomal protein L6 Ni et al. 2006 RpL7-like #6 29.2 k Ribosomal protein L7-like Ni et al. 2006 RpL7a #6 30.7 k Ribosomal protein L7a Ni et al. 2006 RpL8, isoform A #6 27.9 k Ribosomal protein L8 Ni et al. 2006
Supplemental Table 2.
Table S2. Statistical analyses of the immuno-FISH experiments.
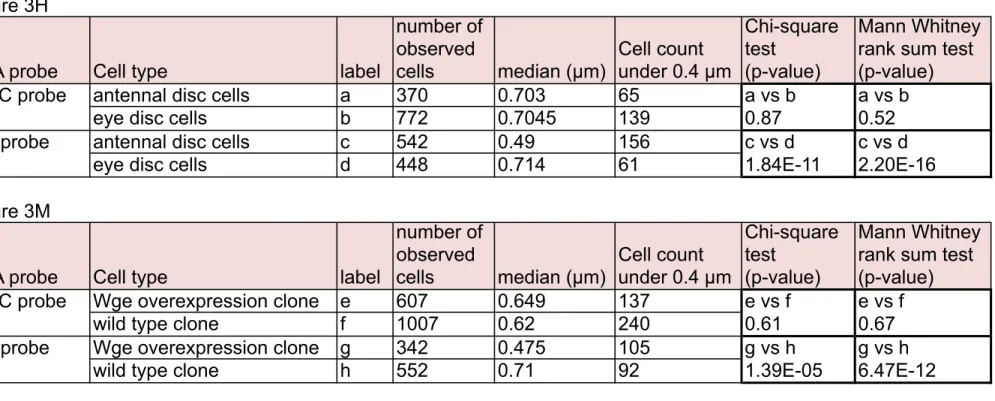
The details of experiments of Figure 3H and 3M (number of observed cells and p-value for statistical analyses) are shown. We performed statistical analyses using two approaches. We analyzed frequencies of FISH foci that were proximate to the NP (within 0.4 µm) (Kohwi et al. 2013), and compared values by the chi-square test. Also, we applied the Wilcoxon-Mann-Whitney rank sum test to compare the median value of FISH dots-NP distances between two independent groups by nonparametric statistics following a previous report (Harmon & Sedat 2005).
Figure 3H
DNA probe Cell type label
number of observed cells median (µm) Cell count under 0.4 µm Chi-square test (p-value) Mann Whitney rank sum test (p-value) His-C probe antennal disc cells a 370 0.703 65 a vs b a vs b
eye disc cells b 772 0.7045 139 0.87 0.52
eya probe antennal disc cells c 542 0.49 156 c vs d c vs d eye disc cells d 448 0.714 61 1.84E-11 2.20E-16 Figure 3M
DNA probe Cell type label
number of observed cells median (µm) Cell count under 0.4 µm Chi-square test (p-value) Mann Whitney rank sum test (p-value) His-C probe Wge overexpression clone e 607 0.649 137 e vs f e vs f
wild type clone f 1007 0.62 240 0.61 0.67
eya probe Wge overexpression clone g 342 0.475 105 g vs h g vs h wild type clone h 552 0.71 92 1.39E-05 6.47E-12