熊本大学学位論文
SPF 環境はトランスサイレチンアミロイドーシスの 発症を防止する
井上聖也
Specific pathogen free conditions prevent transthyretin amyloidosis in mouse models
Seiya Inoue
Transthyretin (TTR) associated amyloidosis is an autosomal dominant disorder characterized by peripheral and autonomic neuropathy. Both genetic and environmental factors are thought to be involved in development of TTR associated amyloidosis. Previously, we demonstrated that amyloid deposition was observed in various tissues of transgenic mouse lines carrying a human mutant TTR (Met30) gene. To analyze the influence of environmental factors on TTR amyloidosis, these amyloidogenic transgenic mouse models were kept under conventional (CV) or specific pathogen free (SPF) conditions. Although the serum levels of Met30 for mice housed in the CV and SPF conditions were similar, amyloid deposition was observed in CV conditions, but not in SPF conditions. In addition, the extent of amyloid deposition in transgenic mice was dependent on duration kept under CV conditions. There were significant differences in proportion of amyloid deposition in several tissues between CV and SPF conditions. Maintenance of these mice at 30o C did not induce amyloid deposition in SPF conditions. These results suggest that the SPF conditions can completely prevent amyloid deposition, and that environmental factors can affect the onset and progression even in a single gene disorder.本論文で使用した略語一覧表 AD : Alzheimer’s Disease
ATTR : transthyretin-associated amyloidosis cDNA:complementary DNA
CD4+:Cluster of Differentiation 4+
CV : Conventional
FAP : Familial Amyloidotic Polyneuropathy hTTR : human transthyretin
IL4 : Interleukin 4 LPS : Lipopolysaccharide Met : Methionine
MT : Metallothionein
PCR : Polymerase Chain Reaction RBP : retinol binding protein SAM : Senescence-Accelerated Mouse SAP : Serum Amyloid P component SOD : Super Oxide Dismutase SPF : Specific Pathogen Free Tg : Transgenic mouse
TTR : Transthyretin Val : Valine
発表論文
本論文は、学術雑誌に掲載された次の論文を基礎とするものである。
(1) Specific pathogen free conditions prevent transthyretin amiloidosis in mouse models
Transgenic Research Accepted: 6 March 2008
Inoue S, Ohta M, Li Z, Zhao G , Takaoka Y , Sakashita N , Miyakawa k , Takeda K , Tei H , Suzuki M , Masuoka M , Sakaki Y ,
Takahashi K ,Yamamura K.
目次
Abstract ……… 2
略語一覧 ……… 3
発表論文 ……… 4
目次 ……… 5
要旨 ……… 7
序論 1)TTR 遺伝子とその機能 ……… 9
2) TTR が関与するアミロイドーシス ……… 9
3) モデルマウスの作製 ……… 11
4) 環境要因 ……… 12
材料と実験方法 1) トランスジェニックマウス ……… 17
2) 飼育条件 ……… 17
3) Tg マウスの同定 ……… 18
4) 組織化学的解析 ……… 18
5) ウエスタンブロット法 ……… 19
6)統計学的解析 ……… 19
結果 1)アミロイド沈着は SPF 環境下では起こらなかった ……… 20
2)CV 環境とアミロイド沈着の関係 ……… 21
3)30℃飼育下でアミロイド沈着は起こらなかった ……… 22
考察 ……… 30
謝辞 ……… 35
参考文献 ……… 36
要旨
「SPF 環境はトランスサイレチンアミロイドーシスの発症を防止する」
生命薬科学専攻 臓器形成分野所属 井上聖也 トランスサイレチン(TTR)遺伝子が関与するアミロイドーシスは、常染色体の 優性遺伝病で、主に末梢神経や自律神経に障害を来す疾患であるが、その発症に は遺伝的要因と環境要因が深く関わっていると考えられる。
これまでに臓器形成分野の研究室では、ヒト変異 TTR 遺伝子(Met30)を導入した トランスジェニック(Tg)マウスを用いることにより、様々な組織にアミロイド が沈着することが確認されている。そこで、TTR アミロイドーシスにおける環境 因子の影響を解析するために、本実験では 4 種類の Tg マウスを作製し、これらを 異なる環境下において長期飼育し、アミロイド沈着の有無を検討した。
4 種類の Tg マウスとは、①メタロチオネイン(MT)プロモーターを接続し、さま ざまな組織に hMet30 を発現する Tg(MT-Met30)マウス、②hTTR 遺伝子の上流 0.6kb 領域を含む hMet30 遺伝子を導入した Tg(0.6h-Met30)マウス、③hTTR 遺伝子の 上流 6.0kb 領域を含む hMet30 遺伝子を導入した Tg(6.0h-Met30)マウス、そして
④hTTR 遺伝子の上流 7.2kb 領域に hMet30cDNA を接続した Tg(7.2h-Met30)マウス である。これらの Tg マウスをコンベンショナル(CV)な環境下と特定な病原菌が いない清浄な(SPF)環境下において長期飼育し、アミロイド沈着の比較を行なっ た。以下に本研究で得られた知見を要約する。
1)Tg(0.6-Met30)マウス、Tg(6.0-Met30)マウス、Tg(MT-Met30)マウスを用いて、
24 ヶ月齢まで飼育してアミロイドの沈着を比較した。その結果、まず 6〜9 ヶ 月齢で CV 環境飼育の MT-Met30 マウスのみが 1/8 匹に沈着が認められた。12〜
18 ヶ月齢では CV 環境の Tg(0.6-Met30)マウスが 4/8 匹、Tg(6.0-Met30)マウス が 7/11 匹、Tg(MT-Met30)マウスが 10/13 匹にアミロイド沈着が確認できた。
21〜24 ヶ月齢では、やはり CV 環境飼育マウスの Tg(0.6-Met30)マウスが 4/6 匹、Tg(6.0-Met30)マウスが 6/6 匹、Tg(MT-Met30)マウスが 7/7 匹において沈着 していた。一方、SPF 環境飼育マウスはいずれの系統においても一例も沈着が 認められなかった。しかし CV、SPF 環境で飼育したマウスの血中の Met30 量を ウエスタンブロット法で比較すると、両者に違いはなかった。
アミロイド沈着はさまざまな組織に分布していたが、沈着率は MT-Met30>
6.0-Met30>0.6-Met30 の順であった。また FAP 患者で見られるような自律神経、
末梢神経への沈着はこれまでの報告と同様、認められなかった。
2) Tg(7.2h-Met30)マウスを使用して、飼育環境とその期間を途中から変更する 4 つの群を設定し、アミロイド沈着を比較した。すなわち SPF24(24 ヶ月 SPF で 飼育)、SPF14/CV10(SPF で 14 ヶ月飼育した後、CV で 10 ヶ月飼育)、SPF8/CV16(SPF で 8 ヶ月飼育した後、CV で 16 ヶ月飼育)、CV24(CV で 24 ヶ月飼育)である。ア ミロイド沈着匹数は、順に 0/17、1/14、6/19、8/8 であり、
SPF24 の群と SPF8/CV16、CV24 の群間では明らかな有意差が認められた。
本実験から CV 飼育期間が長くなるほどアミロイド沈着は相関し増大するとい うことが明らかになった。
3)飼育環境の指標の一つである温度による影響を見るため、Tg(0.6-Met30)マウ ス、Tg(6.0-Met30)マウス、Tg(MT-Met30)マウスを用いて SPF 環境下で 30℃の 高温で長期飼育した場合のアミロイド沈着を観察した。6 ヶ月齢、12 ヶ月齢、
18 ヶ月齢、24 ヶ月齢で観察した結果、いずれの系統でも沈着は認められなか った。
以上の結果より、トランスサイレチンアミロイドーシスモデルマウスにおける アミロイドの沈着が、SPF という清浄な環境下で飼育された場合は、起こらない こと、コンベンショナル(CV)な環境で飼育された場合のみ沈着すること、しか もその度合いは CV 環境での飼育期間と密接に関連しているということが判明し た。CV 環境において沈着を誘導する要因が何かということは未だ不明である。し かし、環境因子が単一遺伝病においてさえ、その発症と進行に大きく影響すると いう事が明らかになり、本研究はアミロイド沈着における環境要因の解明に向け た重要なステップであると考えられる。
序論
1) TTR 遺伝子とその機能
トランスサイレチン(TTR)蛋白の分子量は 13761Dalton で同一の単量体が重合 して4量体を形成する(Kaneda et al.1974)。またこの4量体は甲状腺ホルモン
(T4)とレチノール結合蛋白(RBP)に親和性の高い、独立した2つの結合部位を もち、血中および髄液中で甲状腺ホルモンやレチノール(ビタミン A)を運搬す ることが知られている。ちなみにトランスサイレチンという名称はその事に由来 している。サイはサイロイド、レチンはレチノールを意味する。TTR 単量体の高 次構造は 2 層のβシートがあり、それぞれの 1 層は 4 本の逆方向のβ―strand か らなる(図1B)。
ヒトとマウスの TTR 遺伝子はどちらも 18 番染色体上にあり、4 つのエクソンから 構成されている。マウス TTR は全長約 8909 塩基対であり、cDNA は 1053 個塩基、
アミノ酸は 127 個である(図 1A)。
遺伝子は、主として肝臓で発現するが、脳の脈絡叢および網膜でも発現している。
細胞の中で 4 量体を形成し、それが血中に分泌される。血清蛋白の一つである。
2) TTR が関与するアミロイドーシス
TTR が関与するアミロイドーシス(ATTR)は、変異した TTR が安定的かつ不溶 性の繊維を重合し(図 3)、末梢神経をはじめ全身の臓器や組織の細胞外にアミ ロイドを沈着させる常染色体優性遺伝病である(Andrade,1952;Glenner1980a,b)。
これまでに約 110 種類の変異 TTR が確認されているが、そのうち 90 種以上はヒト のアミロイドーシスに関与している(表 1)。また、変異の型では、TTR 蛋白の
30 番目のバリン(Val)がメチオニン(Met)に置換したタイプが最も一般的であ り、この変異型は、日本(Tawara et al.,1983)、ポルトガル(Saraiva et al.,1983)、
スウェーデン(Westermark st al.,1985)それにアメリカ(Benson and Dwulet,1985)
では特に多く、他の置換タイプを圧倒している。
臨床的には、末梢神経や腸、腎臓、心臓など各種組織にアミロイドが沈着し、
機能障害を起こさせる事により、最終的に死に至る疾患である(Glenner,1980a,b)。
興味深い事には、日本、ポルトガル、スウェーデンにおいて、その患者の発症平 均年齢が大きく異なっている。さらには、本疾患は同一家族においてさえ、その 発症年齢がさまざまである。これらの知見は TTR 遺伝子において、その発症には、
遺伝子の変異以外に環境要因が重要である事を示唆している。
3) モデルマウスの作製
環境因子の解明には、モデルマウスを作製することが重要である。臓器形成分 野の研究室では、これまで異なるプロモーターをもつ 4 種類のヒト変異 TTR 遺伝 子を導入したトランスジェニック(Tg)マウスを作製してきた(図 2)。即ち、
第一にメタロチオネイン(MT)遺伝子プロモーターを接続し、さまざまな組織に Met30 を発現する Tg(MT-hTTRMet30)マウス(Wakasugi et al.,1987)である。
第 二 に hTTR 遺 伝 子 の 上 流 0.6kb 領 域 を 含 む hMet30 遺 伝 子 を 導 入 し た Tg(0.6-hTTRMet30)マウスである。上流 0.6kb は肝臓特異的に発現させるのに重要 な領域であるが、脈絡叢では発現するのに不十分である事は確認されていた
(Yamamura et al.,1987)。三番めには、hTTR 遺伝子の上流 6.0kb 領域を含む hMet30 遺伝子を導入した Tg(6.0-hTTRMet30)マウスである。この場合、導入遺伝子は肝
臓だけでなく、脈絡叢においても発現する(Nagata et al.,1995)。最後に hTTR 遺伝子の上流 7.2kb 領域を含む hMet30cDNA を導入した Tg(7.2-TTRMet30)マウ スである(Takaoka et al.,2004)。
4 種類全ての Tg マウスは種々の組織にアミロイド沈着を起こすことが証明されて いる(Takaoka et al.,2004;Takaoka et al.,1997;Wakasugi et al.,1987;Yi et al.,1991)。したがって、これらの Tg マウスを ATTR アミロイドーシスにおいて、
環境因子を解析するための最適な研究モデルとして使用した。
4)環境要因
環境要因はさらに内因性のものと外因性のものに分けられる。例えば Tg マウ スの胃において、アミロイドの沈着は非腺部に認められるものの腺部には認めら れない(Takaoka et al.,1997)。これはアミロイド沈着が限局した組織構造のよ うな内因性の要因と関係していることを示唆している。また Tg マウスにおいて、
弱病原性の腸内フローラのような外因性の要因が、アミロイドの沈着に影響する
(Noguchi et al.,2002)。
本研究では、外因性の環境要因がアミロイド沈着に影響する事を解析した。最 近の研究では環境要因が DNA やクロマチンの修飾を通して遺伝子発現に影響を及 ぼすという報告もある(Jirtle and Skinner.,2007)。我々はアミロイドの沈着 が SPF という清浄な環境で飼育された場合は起こらないこと、コンベンショナル
(CV)な環境で飼育された場合のみ沈着し、その度合いは CV 環境での飼育期間と 関連している事を確認した。
A
948bp 3392bp 3516bp
EXON 1 EXON 2 EXON 3 EXON4
95bp 131bp 136bp 691bp
8.909kb cDNA:1053,Amino acids:127
B
図 1. A: TTR 遺伝子の構造 B. TTR の単量体の立体構造 :2 層のβシート
(A-H)をもち、それぞれの 1 層は 4 本の逆方向のβ-strand からなる。
*
-GTG- normal 6.0-hMet30 gene
*
*
7.2-hMet30 gene
Poly A
図 2.導入遺伝子の構造。0.6-hMet30 は、上流約 600bp を含む変異トランスサイレ チン遺伝子である。6.0-hMet30 は、上流約 6kb を含む変異トランスサイレチン遺 伝子である。MT-hMet30 は、メタロチオネインプロモーターをつないだ変異トラ ンスサイレチン遺伝子である。7.2-hMet30 は、hTTR 遺伝子の上流約 7.2kb を接続
0.6-hMet30 gene
MT-hMet30 gene(Metallothionein promotor) -ATG- mutant
hMet30 cDNA
表 1.種々の異型 TTR が沈着するアミロイドーシス
アミノ酸 置換部位
正常 異型 症状、沈着組織等
10 Cys Arg 末梢神経障害(PN)、自律神経障害(AN)、眼、心、[遅発型]
12 Leu Pro PN、心、肝、脳軟膜
18 Asp Glu PN
20 Val Ile 心、手根管症候群(CTS)
23 Ser Asn PN、心、眼
24 Pro Ser PN、心、CTS, [遅発型]
30 Val Met PN、AN、眼、第Ⅰ型 FAP 30 Val Leu PN、AN
30 Val Ala PN、AN、心
33 Phe Ile PN、眼
33 Phe Leu PN、心
33 Phe Val PN
34 Arg Thr PN、心
35 Lys Asn PN、AN、心
36 Ala Pro PN、眼
38 Asp Ala 心
42 Glu Gly PN、AN、心
42 Glu Asp 心
44 Phe Ser PN、AN、心
45 Ala Thr AN、心
45 Ala Asp PN、心
45 Ala Ser 心
47 Gly Arg PN、AN
47 Gly Ala PN、心
47 Gly Val CTS、PN、AN、心 49 Thr Ala PN、AN、眼、CTS、心
49 Thr Ile PN、心
50 Ser Arg PN、AN
50 Ser Ile PN、AN、心、眼
51 Glu Gly 心
52 Ser Pro PN、AN、心、腎
54 Glu Gly PN、AN、眼
54 Glu Lys PN、AN、心、眼、[劇症型]
55 Leu Pro PN、AN、心、眼、[劇症型]
58 Leu His CTS、PN、心
58 Leu Arg CTS、AN、眼
59 Thr Lys PN、AN、心
60 Thr Ala PN、AN、CTS、心、[遅発型]
61 Glu Lys PN
64 Phe Leu CTS、PN、心
64 Phe Ser PN、眼、LM
68 Ile Leu 心
69 Thy His 眼
70 Lys Asn CTS、PN、眼
71 Val Ala CTS、PN、AN 73 Ile Val PN、AN
77 Ser Tyr PN、AN、CTS、心、[遅発型]
77 Ser Phe PN
84 Ile Ser CTS、PN、AN、心、第Ⅱ型 FAP
84 Ile Asn 眼、心、CTS
84 Ile Thr PN、心
89 Glu Gln PN、心
89 Glu Lys PN、心
91 Ala Ser PN、心、CTS
97 Ala Gly PN、心、CTS, [遅発型]
107 Ile Val PN、CTS, 心、[遅発型]
111 Leu Met 心
112 Ser Ile PN、心 114 Tyr Cys PN、AN、眼 114 Tyr His CTS 116 Tyr Ser PN
120 Ala Ser 心
122 Val Ala PN、心、眼
122 Val 欠失 PN、心
図3. アミロイド繊維の形成過程
SAP (serum amyloid P component):血清アミロイド P 成分
変異 TTR は不安定なので、4 量体が単量体に解離しやすい。次に単量体の 3 次元 構造がほぐれて可溶性の蛋白質重合体となり、不溶性のアミロイド繊維が形成さ れる。最後にアミロイドタンパクに SAP が付着する事により、アミロイドが分解 されにくくなると考えられている。
4量体 単量体
3次元構造が ほぐれた単量体
変性 単量体
可 溶 性 重合体
アミロイド 線維
SAP付着
解離 修飾 重合 不溶化
材料と実験方法
1)トランスジェニックマウス
実験には 4 種類の Tg マウス、すなわち MT-Met30 ライン、0.6-Met30 ライン、
6.0-Met30 ラインと 7.2-Met30 ラインを使用した(Nagata et al.,1995;Takaoka et al.,2004;Wakasugi et al.,1987;Yamamura et al.,1987)。
表 2 にはそれら 4 つのラインの特徴を示している。MT-Met30 マウスは BDF1 の雌 雄の交配により得られた受精卵を用いる事によって独自に作出した。系統樹立後、
血中の Met30 の発現が最も高い No.5 のラインを選抜した。このマウスは 10 代以 上、C57BL/6 と戻し交配を行なった。その他の系統は C57BL/6 の受精卵を用いた。
同様にそれぞれ 0.6-Met30、6.0-Met30、7.2-Met30 系統の、発現の高い No.61、No.15、
No.2 ラインを使用した。過去の文献では、6 から 9 ヶ月、12 から 18 ヶ月、21 か ら 24 ヶ月ではアミロイドの沈着は同等であったことが示されている(Yi et al.1991;Takaoka et al.1997;Taakaoka et al.2004)。それゆえ本実験では、6 か ら 9 ヶ月、12 から 18 ヶ月、21 から 24 ヶ月齢の3群に分け、アミロイドの沈着を 解析することにした。これらのマウスに関する実験は、ヘルシンキ宣言にのっと り、熊本大学の動物実験倫理委員会の承認を得た上で実施した。
2)飼育条件
トランスジェニックマウスはコンベンショナル(CV)環境を井上実験動物セン ター(下益城郡中央町)の飼育室、清浄(SPF)環境を熊本大学動物資源開発研究 センター(熊本市)の SPF エリアおよび武田薬品工業の研究開発部実験動物室を 用いて飼育した。これらのマウス飼育室は、昼夜 12 時間の照明サイクルで管理さ
れており、室温は 22±2℃であった。飼育温度の影響を見るための実験は、武田 薬品工業の研究開発部実験動物室において、SPF 状態で 30℃の環境を作り、その 中でマウスを飼育した。給餌は一般的な市販のマウス用固形飼料を与え、水は自 由給水とした。
3)Tg マウスの同定
Tg マウスは Polymerase chain reaction(PCR)とウエスタンブロット法により、
遺伝子型を同定した(図 4)。マウスの尾から DNA を抽出し、2 組の hTTR プライマ ーセットを用いて PCR を行なった。ウエスタンブロット法は引用論文にしたがっ て実施した(Nagata et al.,1995)。血液はエーテル麻酔下で、眼底動脈から採取 した。マウスの血清は 17%SDS ポリアクリルアミドゲルに電気泳動し、トランス ファーメンブレン(Millipore,USA)に転写した。メンブレンは抗 hTTR ウサギ抗 体(MBL,JAPAN)と反応させた後、抗ウサギ IgG ペルオキシダーゼ標識ヤギ抗体と 反応させた。TTR 蛋白は、ケミルミネッセンス検出システム(ECL,Amersham,USA)
を用いてプロトコールにしたがって検出した。
4) 組織化学的解析
全ての Tg マウスは頸椎脱臼により安楽死させた後、心臓、腎臓、脾臓、肝臓、
肺、膵臓、胃、小腸、大腸、膀胱、甲状腺、リンパ節、骨髄、座骨神経、自律神 経、脳を採取し、10%中性ホルマリン液で固定し、パラフィン包埋
を行なった(Yi et al.,1991)。その後、パラフィン標本は HE 染色し、観察した。
アミロイドの組織化学的観察のためにパラフィン切片は文献にしたがって過マン ガン酸カリウム(KMnO4)で固定した後、コンゴレッドで染色を行なった(Yi et
al.,1991)。コンゴレッド染色を行ない、偏光顕微鏡下で観察するとアミロイド沈 着が存在したところが、黄緑色の複屈折光を発するため検出できる。また免疫組 織学的解析のためには、パラフィン切片を間接免疫ペルオキシダーゼ法で染色し た。その場合の抗体は、抗ヒトプレアルブミン(Beringwerke,Marburg,Germany)
と抗マウス SAP(BehringDiagnostics,LaJolla,USA)を用いた。
5) ウエスタンブロット法
血中 hTTR の検出に、ウエスタンブロット法を用いた。まず、0.9%NaCl で 1:
50 に希釈したマウス血清 6μℓを 12%ポリアクリルアミドゲルの各レーンにアプ ライし、電気泳動を行なった。次に immobilon polyvinylidene difluode filter
(Millipore,Billerica,MA,USA)に転写した。
一次抗体はウサギ抗ヒト TTR 抗体を指示通り 1:1000 に希釈して使用した
(MBL,Nagoya,Japan)。検出用抗体は HRP を標識した抗ウサギ IgG 抗体を使用した
(Amersham Japan,Tokyo)。 6)統計学的解析
表 5 と 表 4 に 示 す デ ー タ を 比 較 す る の は 、 StatView software ( SAS Institute,Cary,NC)を用いた。表 4 においては、グループ間の統計的差は Kruskal-Wallis test と Mann-Whitney U test を用いて検討した。(P<0.05)で有 意差ありと判断した。
表 5 においては、グループ間の統計的意味は Fisher’s exact test を用いて検 討した。(P<0.05)で有意差ありと判断した。
結果
1)アミロイド沈着は SPF 環境下では起こらなかった
外因性の環境要因がアミロイド沈着に影響するかどうかをまず、MT-Met30、
0.6-Met30、6.0-Met30 の 3 つの Tg マウス系統を用いて検討した。その理由はこ れら 3 系統のマウスは cDNA ではなく、Met30 のヒトゲノム DNA を導入した Tg マ ウスであるからである。つまり、ゲノム DNA と cDNA の構造上の違いから来る影響 をなるべく無くし、プロモーターの違いが影響する環境要因だけを観察すること ができるからである。3 系統の Tg マウスは CV と SPF 環境下で 24 ヶ月齢まで飼育 し、アミロイドの沈着を比較した。その結果、まず CV 環境下の MT-Met30 マウス が 6—9 ヶ月齢で 1 匹沈着が認められた。次に 12—18 ヶ月齢時点で、0.6-Met30 ラ インの 4/8 匹、6.0- Met30 ラインの 7/11 匹、MT-Met30 ラインの 10/13 匹にアミ ロイド沈着が確認された。最後の 21—24 ヶ月齢の時点では、0.6-Met30 ラインの 4/6 匹、6.0- Met30 ラインの 6/6 匹、MT-Met30 ラインの 7/7 匹に沈着が起こった。
一方、SPF 環境飼育群では、どのステージにおいてもアミロイドは沈着しなかっ た。(図 5 と表 2)
表 4 には、CV 環境下で飼育したマウスのアミロイド沈着について、その比率、部 位、相対的量を示しており、0.6-Met30、6.0-Met30、MT-Met30 間で、数種の組織 においては、アミロイド沈着の割合に統計上の有意差があった。しかし、沈着の パ タ ー ン は 過 去 の 文 献 と 同 様 で あ っ た ( Kohno et al.,1997;Takaoka et al.,1997;Yi et al.,1991)。次に CV と SPF 環境が、各 Tg マウスにおいて、血中 のヒト TTR 値に影響しているかどうかをウエスタンブロット法により、検討した。
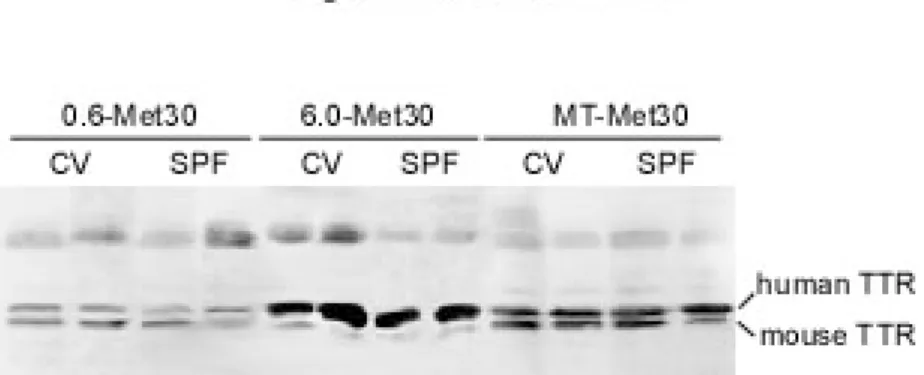
CV と SPF 環境飼育の各 Tg マウスの血中の Met30 値は、これまでの報告通り(Tagoe et al.,2003)、同様であった(図 6)。
これらの結果は、SPF 環境で飼育することが、マウス血中の Met30 のレベルに関 係なく、アミロイド沈着を抑制することを示唆している。
2)CV 環境とアミロイド沈着の関係
CV 環境下でのアミロイド沈着について、我々はマウスを CV 環境で飼育する期 間がアミロイド沈着の程度に関係するかどうかという検討を試みた。本実験には 7.2-hMet30 ラインを使用した。このラインはゲノム Met30 遺伝子の代わりにヒト Met30 の cDNA が導入されているが、他の系統と同じようなアミロイド沈着
(Takaoka et al.,1997;Yi et al.,1991)が起こる事が確認されている(Takaoka et al.,2004)。この Tg マウスは4つのグループに分けられた。すなわち第一に SPF24 というグループであるが、これは SPF 環境下に 24 ヶ月飼育された。第二番 目は、SPF14/CV10 というグループで、これは SPF に最初の 14 ヶ月、その後 CV 環 境で 10 ヶ月飼育した群である。第三番目のグループは、SPF8/CV16 であるが、こ れは出生後 SPF 環境に 8 ヶ月、その後 CV 環境で 16 ヶ月飼育したグループである。
第四グループは、CV24 でこれは CV 環境に 24 ヶ月飼育した。アミロイド沈着は、
SPF24 で 17 匹中 0 匹、SPF14/CV10 で 14 匹中 1 匹、SPF8/CV16 グループで 19 匹中 6 匹、CV24 グループで 8 匹中 8 匹が確認された。SPF24 のグループと SPF8/CV16、
CV24 のグループ間ではアミロイド沈着において、統計学上、明らかな有意差が認 められた(表 5)。またアミロイド沈着は、小腸と腎臓において、最も際立ってお り、心臓、甲状腺、胃の上部が次に多かった。アミロイド沈着のこれらの分布傾
向 は 過 去 の 論 文 と 同 様 な 結 果 で あ っ た ( Takaoka et al.,2004; Takaoka et al.,1997;Yi et al.,1991)。またこれらの結果は、アミロイド沈着が CV 環境で飼 育された期間と大いに関連している事を明確に示唆している。
3)30℃飼育下でアミロイド沈着は起こらなかった
SPF と CV 環境では例えば腸内フローラ、温度、湿度など多くの環境の違いが存在 する。我々は、高温下では血管が拡張するため、4 量体が細胞外に漏出し、それ が結果的にアミロイド沈着を誘導するのではないかということを考えた。加えて 飼育室の温度のコントロールは比較的容易である。この可能性を検討するために、
MT-Met30、0.6-Met-30、6.0-Met-30 のラインのマウスを SPF 環境下で 2 年間飼育 した。各々の系統から 4 匹ずつを 6 ヶ月毎に病理解析したが、アミロイド沈着は どの Tg マウスにおいても確認されなかった。
Tg Tg Wt Tg Tg Wt
図4.PCRを用いた genotyping
2 組のプライマーを作製して、274bp と 231bp のバンドを検出した。
表 2.
各トランスジェニックマウス系統の特徴
Tg ライン プロモーター コード領域 血中濃度
(mg/dl)
0.6-hMet30 hTTR の 0.6kb 上流 ゲノム DNA V30M 1.5-3.0 6.0-hMet30 hTTR の 6.0kb 上流 ゲノム DNA V30M 13.7-14.5
MT-hMet30 マウスメタロチオネインⅠ ゲノム DNA V30M 1.0-4.8 7.2-hMet30 hTTR の 7.2kb 上流 cDNA V30M 4.8-5.7
表 3.
アミロイド沈着における SPF 環境の効果
Tg ライン 月齢
6-9 12-18 21-24 CV
0.6-hMet30 0/2 4/8 4/6 6.0-hMet30 0/12 7/11 6/6 MT-hMet30 1/8 10/13 7/7 SPF
0.6-hMet30 0/8 0/12 0/8 6.0-hMet30 0/8 0/8 0/8 MT-hMet30 0/9 0/12 0/8
図 5. 24 ヶ月齢の各 Tg マウスの小腸と腎臓。コンゴレッド染色をし、偏光顕微 鏡下で観察するとアミロイド沈着部位が緑色に蛍光する。CV 環境で飼育されたマ ウスには沈着が認められたが、SPF 環境で飼育されたマウスには沈着していなか った。 バー;100μm
図 6. 抗ヒト TTR 抗体を使用したウエスタンブロット。血中 Met30 値は SPF 飼育 下でも全ての系統において変わらなかった。
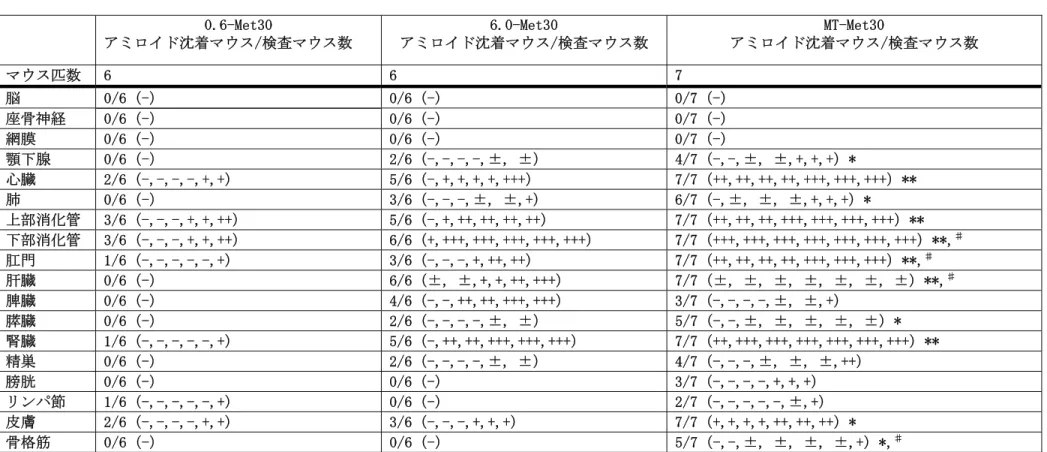
表 4 21〜24 ヶ月齢の各 Tg マウスにおける TTR アミロイド沈着の組織分布 0.6-Met30
アミロイド沈着マウス/検査マウス数
6.0-Met30
アミロイド沈着マウス/検査マウス数
MT-Met30
アミロイド沈着マウス/検査マウス数
マウス匹数 6 6 7
脳 0/6(-) 0/6(-) 0/7(-)
座骨神経 0/6(-) 0/6(-) 0/7(-)
網膜 0/6(-) 0/6(-) 0/7(-)
顎下腺 0/6(-) 2/6(-,-,-,-,±, ±) 4/7(-,-,±, ±,+,+,+)*
心臓 2/6(-,-,-,-,+,+) 5/6(-,+,+,+,+,+++) 7/7(++,++,++,++,+++,+++,+++)**
肺 0/6(-) 3/6(-,-,-,±, ±,+) 6/7(-,±, ±, ±,+,+,+)*
上部消化管 3/6(-,-,-,+,+,++) 5/6(-,+,++,++,++,++) 7/7(++,++,++,+++,+++,+++,+++)**
下部消化管 3/6(-,-,-,+,+,++) 6/6(+,+++,+++,+++,+++,+++) 7/7(+++,+++,+++,+++,+++,+++,+++)**,♯ 肛門 1/6(-,-,-,-,-,+) 3/6(-,-,-,+,++,++) 7/7(++,++,++,++,+++,+++,+++)**,♯
肝臓 0/6(-) 6/6(±, ±,+,+,++,+++) 7/7(±, ±, ±, ±, ±, ±, ±)**,♯
脾臓 0/6(-) 4/6(-,-,++,++,+++,+++) 3/7(-,-,-,-,±, ±,+)
膵臓 0/6(-) 2/6(-,-,-,-,±, ±) 5/7(-,-,±, ±, ±, ±, ±)*
腎臓 1/6(-,-,-,-,-,+) 5/6(-,++,++,+++,+++,+++) 7/7(++,+++,+++,+++,+++,+++,+++)**
精巣 0/6(-) 2/6(-,-,-,-,±, ±) 4/7(-,-,-,±, ±, ±,++)
膀胱 0/6(-) 0/6(-) 3/7(-,-,-,-,+,+,+)
リンパ節 1/6(-,-,-,-,-,+) 0/6(-) 2/7(-,-,-,-,-,±,+)
皮膚 2/6(-,-,-,-,+,+) 3/6(-,-,-,+,+,+) 7/7(+,+,+,+,++,++,++)*
骨格筋 0/6(-) 0/6(-) 5/7(-,-,±, ±, ±, ±,+)*,♯
-,アミロイド沈着無し;±,沈着は血管壁限定;+,沈着は血管壁とその周辺;++,中等度の沈着が間質にあり;+++,顕著な沈着が実質と間質にあり
*P<0.05 対 0.6-Met30. **P<0.01 対 0.6-Met30. ♯P<0.05 対 6.0-Me
表 5.
アミロイド沈着にける CV 環境の効果
*P<0.05 SPF8/CV16 対 SPF24 グループ;**P<0.0001 CV24 対 SPF24 グループ;♯ P<0.0001CV24 対 SPF14/CV10 グループ;+P<0.01 CV24 対 SPF8/CV16 グループ
SPF24 0/17
SPF14/CV10 1/14
SPF8/CV16 6/19*
CV24 8/8** ,♯,+
SPF CV
実験環境 0 2 4 16 18 20 22
飼育期間(月)
アミロイドの 24 沈着匹数
6 8 10 12 14
考察
我々は、Tg マウスにおいて、SPF 環境がアミロイド沈着を完全に抑制し、環境 要因としての CV 下での飼育がアミロイド沈着を促進することを明らかにした。
多くのヒトの疾患において、遺伝的要因と環境要因が関連している事が知られ ている。一般的な疾患の例として、癌や糖尿病は、遺伝子と食生活など環境要因 の複合的な要因の結果、発症するとされている。さらに環境要因は単一遺伝子病 の発症にさえ関与しており、例えばハンチントン病では発症する年齢の違いには、
環境の影響が 63%を占めていると言われている(Wexler et al.,2004)。しかし 環境要因には、ストレス、肉体的、精神的虐待、食事、毒や病原菌、放射線や化 学物質への曝露、気候、その他などを含んでおり、一口に患者への環境要因が何 であるのか、決定するのは困難である。したがって、これまでもヒト疾患におけ る環境要因の解析にはモデル動物が有効なツールとして使用されてきた。
Matsuda ら(Matsuda et al.,1997)は、近交系の NC/Nga というマウスを CV 環 境で飼育すると、IgE の過剰産生を伴うヒトアトピー性皮膚炎に類似した皮膚の 病変を自然発症的に起こすが、SPF 環境では起こらないことを示している。この 皮膚の病変は肥満細胞と IgE 合成に必要な IL-4 を含む CD4+の T 細胞が増加を示 しており、これはすなわち環境要因が IgE の過剰生産の引き金になっている事を 示唆している。Kuhn ら(Kuhn et al.,1993)は、IL-10 欠損マウスにおけるヒト 腸炎(IBD)に似た慢性腸炎は、SPF 環境下では比較的軽症でその発症も遅れるこ とを示している。その後、Kulberg らは(Kulberg et al.,1998)、
Helicobacter
hepaticus
の感染が、IL-12 とガンマインターフェロン依存というメカニズムを通じて、SPF 下での IL-10 欠損マウスの大腸炎への引き金になることを明らかにし た。Yoshitomi ら(Yoshitomi et al.,2005)は、SKG マウスが SPF 下では関節炎 が発症しないこと、しかしながらおそらく IL-4 産生(Kobayashi et al.,2006)
を通して、SPF 環境下の SKG マウスにおいては真菌類のベータグルカンが慢性関 節炎の引き金になることを示した。これらのことから自己免疫疾患のモデル動物 が CV 環境下で飼育されるとその反応として、腸内フローラが炎症を誘発したり、
自己免疫の標的になったりするのかもしれない。
本実験では、アミロイド沈着は SPF 環境では全く観察されないこと、また沈着 の度合いは CV 環境下に飼育された期間に依存している事を示した。
Shino らは、SAM(老化促進マウス)におけるアポリポプロテイン A-Ⅱの沈着に特徴 づけられる老人性アミロイドーシスは、SPF 環境で飼育された場合、軽症である
(Shino et al.,1987)。しかしながら SAM マウスにおいて免疫活性が低下するこ とは老人性アミロイドーシスの発症に直接には影響しなかったし、逆にアミロイ ドーシスが免疫活性に影響する事も無かった(Hosono et al.,1997)。我々は以前 LPS 処置がアミロイドの沈着の始まり、進行、分布には影響しないことを明らか にした(Murakami et al.,1992)。また CV 環境の腸内フローラがアミロイド沈着 を容易にすることを確認したが、それは消化管に限定していた。これらの結果は 腸内細菌を仲介する免疫反応は ATTR には関係していないことを示唆している。
Tagoe らは、過去に TTR アミロイドの沈着の頻度、沈着の範囲や性質は、CV と SPF 環境下では明らかな違いは無いと報告している(Tagoe et al.,2003)。彼ら の実験では、ヒトの野生型の TTR 遺伝子を導入した Tg マウスを 12 ヶ月齢まで SPF
環境に飼育し、その後、半数のマウスを CV 飼育室に移動している。両グループの 24 ヶ月齢における TTR と AA アミロイドの発生率は同様ということであった。
しかし、これは TTR アミロイド沈着が SPF 飼育下では Tg マウスのいかなる組織に も認められなかったという我々のデータとは一致していない。この理由として、
我々の SPF と CV の環境は Tagoe らの環境と違うことが考えられる。我々の CV 環 境では AA アミロイドはよく観察されたが、SPF では見られなかった。さらにアミ ロイド沈着の程度は CV 環境で飼育した期間に依存していた。すなわちこれは我々 の実験では CV 環境が SPF 環境と全く異なっている事を示唆している。一方、Tagoe らのマウス飼育室は CV と SPF 環境がそれほど変わらない事を意味している。また 別の可能性は、Tg マウス作製において使用した遺伝子の違いである。我々が変異 した Met30 遺伝子を用いたのに対し、Tagoe らは正常な Val30 遺伝子を使ってい る。Met30 から作られる 4 量体はより不安定という報告もあるが(Ferrao-Gonzales et ak.,2003;Nettleton et al.,1998;Niraura et al.,2002;Quintas et al.,1997)、 我々の実験では CV 環境下で 4 量体の解離がさらに起こりやすかったのかもしれな い。異なる遺伝子のコンストラクトを導入した 3 種類の Tg マウスラインにおいて アミロイド沈着が観察されなかったのは、それが Met-30 遺伝子の発現調節が変え られたことによるものではない事を示している。実際、血中の Met-30 値は SPF 環境でも CV 環境でも同様であった。
CV と SPF 環境の違いは未だに明らかになっていない。室温は環境要因の一つで あるものの、少なくとも SPF 環境でマウスを 30℃で飼育する事がアミロイド沈着 を誘導することはなかった。
以前に我々は、Cys10 のチオール残基がアミロイド形成に関係していることを 確認している(Takaoka et al.,2004)。10 番目のシステインをセリンに置き換え た時、30 番目にはメチオニンが存在するにも関わらず、Tg マウスにおいてアミロ イド沈着は確認されなかった。このようにジスルフィド形成がアミロイドジェネ シスに関与しているかもしれないし、逆に抗酸化物質がアミロイド沈着を抑制し ているのかもしれない。これは強烈な酸化ストレスが ATTR 患者に確認されるとい うデータと一致している(Ando et al.,1997)。さらには変異型細胞外スーパーオ キシドジスムターゼ(SOD)遺伝子を持つ ATTR 患者の場合には、SOD が野生型の ATTR 患者より、アミロイド沈着がかなり重症である(Sakashita et al.,1998)。 この酸化によるダメージはアルツハイマー病(AD)のような他のアミロイドーシ スにおいては重要な役割を果たすようである。ある特定の血漿蛋白の酸化作用が 増えることが AD 患者に起こり、非 AD 患者には起こらない事が報告されている
(Conrad et al.,2000)。まとめると、アミロイド沈着は、CV 条件下で酸化スト レスに遭遇した時に促進されるようである。
より一般的で可能な治療法を開発するためには、アミロイドジェネシスが関係 するような環境因子を発見する事が重要である。4 量体の単量体への解離はアミ ロイドジェネシスにおいて非常に重要な段階であり、TTR4 量体を安定化させる分 子を発見する事に多くの努力が費やされた。これまでに小さな分子や Cr3+が TTR4 量体に結合する事によって 4 量体の構造の安定性を維持する事が示されてきた
( Almeida et al.,2004;Ando,2005;Jonson et al.,2005a;Jonson et al.,2005b;Nettleton et al.,1998;Sato et al.,2006)。さらに ATTR Y78P を用い
て Met30 遺伝子を導入した Tg マウスへ免疫すると、アミロイド沈着を抑制した
(Terazaki et al.,2006)。
肝臓移植(Stangou and Hawkins,2004)は現在も唯一の信頼性のある治療法で ある。しかし一方で、環境因子の影響や効果を明らかにする事は、手術に頼らな い新しい治療法を確立する事につながるものと思われる。本研究はアミロイド沈 着における環境要因の解明に向けた重要なステップであると考えられる。
謝辞
熊本大学発生医学研究センター器官形成部門・臓器形成分野において、熊本大
学薬学教育部、臓器形成学、山村研一教授のご指導の下、本研究を行いました。
終始ご指導ご鞭撻を賜わりましたことを深く感謝いたします。
臓器形成学、李正花博士には日々の実験手技から論文の指導まで幅広いご指導
を頂きました。深く感謝いたします。高岡裕博士、大田美香博士にはマウスの飼
育管理についてご指導を頂き、深く感謝の意を表します。
また臓器形成学教室の皆様には有形無形の多くのご協力とご支援を頂きました。
心から感謝申し上げます。
参考文献
1. Almeida, M.R., Macedo, B., Cardoso, I., Alves, I., Valencia, G., Arsequell, G., Planas, A., and Saraiva, M.J. (2004). Selective binding to
transthyretin and tetramer stabilization in serum from patients with familial amyloidotic polyneuropathy by an iodinated diflunisal derivative
. Biochem J
381, 351-356.2.Ando, Y. (2005). Liver transplantation and new therapeutic approaches for familial amyloidotic polyneuropathy (FAP).
Med Mol Morphol
38, 142-154.3. Ando, Y., Nyhlin, N., Suhr, O., Holmgren, G., Uchida, K., el Sahly, M., Yamashita, T., Terasaki, H., Nakamura, M., Uchino, M., and Ando, M. (1997).
Oxidative stress is found in amyloid deposits in systemic amyloidosis.
Biochem Biophys Res Commun
232, 497-502.4. Andrade, C. (1952). A peculiar form of peripheral neuropathy; familiar atypical generalized amyloidosis with special involvement of the
peripheral nerves.
Brain
75, 408-427.5. Benson, M.D., and Dwulet, F.E. (1985). Identification of carriers of a variant plasma prealbumin (transthyretin) associated with familial amyloidotic polyneuropathy type I.
J Clin Invest
75, 71-75.6.Conrad, C.C., Marshall, P.L., Talent, J.M., Malakowsky, C.A., Choi, J., and Gracy, R.W. (2000). Oxidized proteins in Alzheimer's plasma.
Biochem
Biophys Res Commun
275, 678-681.7. Ferrao-Gonzales, A.D., Palmieri, L., Valory, M., Silva, J.L., Lashuel, H., Kelly, J.W., and Foguel, D. (2003). Hydration and packing are crucial to amyloidogenesis as revealed by pressure studies on transthyretin variants that either protect or worsen amyloid disease.
J Mol Biol
328, 963-974.8. Glenner, G.G. (1980a). Amyloid deposits and amyloidosis. The beta-fibrilloses (first of two parts).
N Engl J Med
302, 1283-1292.9.Glenner, G.G. (1980b). Amyloid deposits and amyloidosis: the
beta-fibrilloses (second of two parts).
N Engl J Med
302, 1333-1343.10. Hosono, M., Hanada, K., Toichi, E., Naiki, H., Higuchi, K., and Hosokawa, T. (1997). Immune abnormality in relation to nonimmune diseases in SAM mice.
Exp Gerontol
32, 181-195.11.Jirtle, R.L., and Skinner, M.K. (2007). Environmental epigenomics and disease susceptibility.
Nat Rev Genet
8, 253-262.12.Johnson, S.M., Petrassi, H.M., Palaninathan, S.K., Mohamedmohaideen, N.N., Purkey, H.E., Nichols, C., Chiang, K.P., Walkup, T., Sacchettini, J.C., Sharpless, K.B., and Kelly, J.W. (2005a). Bisaryloxime ethers as potent inhibitors of transthyretin amyloid fibril formation.
J Med Chem
48, 1576-1587.13.Johnson, S.M., Wiseman, R.L., Sekijima, Y., Green, N.S., Adamski-Werner, S.L., and Kelly, J.W. (2005b). Native state kinetic stabilization as a
strategy to ameliorate protein misfolding diseases: a focus on the transthyretin amyloidoses.
Acc Chem Res
38, 911-921.14.Kobayashi, K., Suda, T., Nan-Ya, K., Sakaguchi, N., Sakaguchi, S., and Miki, I. (2006). Cytokine production profile of splenocytes derived from zymosan A-treated SKG mice developing arthritis.
Inflamm Res
55, 335-341.15.Kohno, K., Palha, J.A., Miyakawa, K., Saraiva, M.J., Ito, S., Mabuchi, T., Blaner, W.S., Iijima, H., Tsukahara, S., Episkopou, V., Gottesman, M.E., Shimada, K., Takahashi, K., Yamamura, K., and Maeda, S. (1997). Analysis of amyloid deposition in a transgenic mouse model of homozygous familial amyloidotic polyneuropathy.
Am J Pathol
150, 1497-1508.16.Kuhn, R., Lohler, J., Rennick, D., Rajewsky, K., and Muller, W. (1993).
Interleukin-10-deficient mice develop chronic enterocolitis.
Cell
75, 263-274.17.Kullberg, M.C., Ward, J.M., Gorelick, P.L., Caspar, P., Hieny, S., Cheever, A., Jankovic, D., and Sher, A. (1998). Helicobacter hepaticus triggers colitis in specific-pathogen-free interleukin-10 (IL-10)-deficient mice through an IL-12- and gamma interferon-dependent mechanism.
Infect Immun
66, 5157-5166.18.Matsuda, H., Watanabe, N., Geba, G.P., Sperl, J., Tsudzuki, M., Hiroi, J., Matsumoto, M., Ushio, H., Saito, S., Askenase, P.W., and Ra, C. (1997).
Development of atopic dermatitis-like skin lesion with IgE hyperproduction
in NC/Nga mice.
Int Immunol
9, 461-466.19.Murakami, T., Yi, S., Maeda, S., Tashiro, F., Yamamura, K., Takahashi, K., Shimada, K., and Araki, S. (1992). Effect of serum amyloid P component level on transthyretin-derived amyloid deposition in a transgenic mouse model of familial amyloidotic polyneuropathy.
Am J Pathol
141, 451-456.20.Nagata, Y., Tashiro, F., Yi, S., Murakami, T., Maeda, S., Takahashi, K., Shimada, K., Okamura, H., and Yamamura, K. (1995). A 6-kb upstream region of the human transthyretin gene can direct developmental, tissue-specific, and quantitatively normal expression in transgenic mouse.
J Biochem
117, 169-175.21.Nettleton, E.J., Sunde, M., Lai, Z., Kelly, J.W., Dobson, C.M., and Robinson, C.V. (1998). Protein subunit interactions and structural integrity of amyloidogenic transthyretins: evidence from electrospray mass spectrometry.
J Mol Biol
281, 553-564.22.Niraula, T.N., Haraoka, K., Ando, Y., Li, H., Yamada, H., and Akasaka, K.
(2002). Decreased thermodynamic stability as a crucial factor for familial amyloidotic polyneuropathy.
J Mol Biol
320, 333-342.23.Noguchi, H., Ohta, M., Wakasugi, S., Noguchi, K., Nakamura, N., Nakamura, O., Miyakawa, K., Takeya, M., Suzuki, M., Nakagata, N., Urano, T., Ono, T., and Yamamura, K. (2002). Effect of the intestinal flora on amyloid deposition in a transgenic mouse model of familial amyloidotic
polyneuropathy.
Exp Anim
51, 309-316.24.Quintas, A., Saraiva, M.J., and Brito, R.M. (1997). The amyloidogenic potential of transthyretin variants correlates with their tendency to aggregate in solution.
FEBS Lett
418, 297-300.25.Sakashita, N., Ando, Y., Marklund, S.L., Nilsson, P., Tashima, K., Yamashita, T., and Takahashi, K. (1998). Familial amyloidotic polyneuropathy type I with extracellular superoxide dismutase mutation: a case report.
Hum Pathol
29, 1169-1172.26.Saraiva, M.J., Costa, P.P., and Goodman, D.S. (1983). Studies on plasma transthyretin (prealbumin) in familial amyloidotic polyneuropathy, Portuguese type.
J Lab Clin Med
102, 590-603.27.Sato, T., Ando, Y., Susuki, S., Mikami, F., Ikemizu, S., Nakamura, M., Suhr, O., Anraku, M., Kai, T., Suico, M.A., Shuto, T., Mizuguchi, M., Yamagata, Y., and Kai, H. (2006). Chromium(III) ion and thyroxine cooperate to stabilize the transthyretin tetramer and suppress in vitro amyloid fibril formation.
FEBS Lett
580, 491-496.28.Shino, A., Tsukuda, R., Omori, Y., and Matsuo, T. (1987). Histopathologic observations on the senescence-accelerated mice (SAM) reared under specific pathogen free conditions.
Acta Pathol Jpn
37, 1465-1475.29.Stangou, A.J., and Hawkins, P.N. (2004). Liver transplantation in
transthyretin-related familial amyloid polyneuropathy.
Curr Opin Neurol
17,615-620.
30.Tagoe, C.E., Jacobson, D.R., Gallo, G., and Buxbaum, J.N. (2003). Mice transgenic for human TTR have the same frequency of renal TTR deposition whether maintained in conventional or specific pathogen free environments.
Amyloid
10, 262-266.31.Takaoka, Y., Ohta, M., Miyakawa, K., Nakamura, O., Suzuki, M., Takahashi, K., Yamamura, K., and Sakaki, Y. (2004). Cysteine 10 is a key residue in amyloidogenesis of human transthyretin Val30Met.
Am J Pathol
164, 337-345.32.Takaoka, Y., Tashiro, F., Yi, S., Maeda, S., Shimada, K., Takahashi, K., Sakaki, Y., and Yamamura, K. (1997). Comparison of amyloid deposition in two lines of transgenic mouse that model familial amyloidotic
polyneuropathy, type I.
Transgenic Res
6, 261-269.33.Tawara, S., Nakazato, M., Kangawa, K., Matsuo, H., and Araki, S. (1983).
Identification of amyloid prealbumin variant in familial amyloidotic polyneuropathy (Japanese type).
Biochem Biophys Res Commun
116, 880-888.34.Terazaki, H., Ando, Y., Fernandes, R., Yamamura, K., Maeda, S., and Saraiva, M.J. (2006). Immunization in familial amyloidotic polyneuropathy:
counteracting deposition by immunization with a Y78F TTR mutant.
Lab Invest
86, 23-31.35.Wakasugi, S., Inomoto, T., Yi, S., Naito, M., Uehira, M., wanaga, T., Maeda, S., Araki, K., Miyazaki, J., Takahashi, K., Shimada, K., and Yamamura, K.
(1987). A transgenic mouse model of familial amyloidotic polyneuropathy.
Proc Japan Acad
63, 344-347.36.Westermark, P., Pitkanen, P., Benson, L., Vahlquist, A., Olofsson, B.O., and Cornwell, G.G., 3rd (1985). Serum prealbumin and retinol-binding protein in the prealbumin-related senile and familial forms of systemic amyloidosis.
Lab Invest
52, 314-318.37.Wexler, N.S., Lorimer, J., Porter, J., Gomez, F., Moskowitz, C., Shackell, E., Marder, K., Penchaszadeh, G., Roberts, S.A., Gayan, J., Brocklebank, D., Cherny, S.S., Cardon, L.R., Gray, J., Dlouhy, S.R., Wiktorski, S., Hodes, M.E., Conneally, P.M., Penney, J.B., Gusella, J., Cha, J.H., Irizarry, M., Rosas, D., Hersch, S., Hollingsworth, Z., MacDonald, M., Young, A.B., Andresen, J.M., Housman, D.E., De Young, M.M., Bonilla, E., Stillings, T., Negrette, A., Snodgrass, S.R., Martinez-Jaurrieta, M.D., Ramos-Arroyo, M.A., Bickham, J., Ramos, J.S., Marshall, F., Shoulson, I., Rey, G.J., Feigin, A., Arnheim, N., Acevedo-Cruz, A., Acosta, L., Alvir, J., Fischbeck, K., Thompson, L.M., Young, A., Dure, L., O'Brien, C.J., Paulsen, J., Brickman, A., Krch, D., Peery, S., Hogarth, P., Higgins, D.S., Jr., and Landwehrmeyer, B. (2004). Venezuelan kindreds reveal that genetic and environmental factors modulate Huntington's disease age of onset.
Proc Natl Acad Sci U S A
101, 3498-3503.38.Yamamura, K., Wakasugi, S., Maeda, S., Inomoto, T., Iwanaga, T., Uehira,
M., Araki, K., Miyazaki, J., and Shimada, K. (1987). Tissue-specific and developmental expression of human transthyretin gene in transgenic mice.
Dev Genet
8, 195-205.39.Yi, S., Takahashi, K., Naito, M., Tashiro, F., Wakasugi, S., Maeda, S., Shimada, K., Yamamura, K., and Araki, S. (1991). Systemic amyloidosis in transgenic mice carrying the human mutant transthyretin (Met30) gene.
Pathologic similarity to human familial amyloidotic polyneuropathy, type I.
Am J Pathol
138, 403-412.40.Yoshitomi, H., Sakaguchi, N., Kobayashi, K., Brown, G.D., Tagami, T., Sakihama, T., Hirota, K., Tanaka, S., Nomura, T., Miki, I., Gordon, S., Akira, S., Nakamura, T., and Sakaguchi, S. (2005). A role for fungal {beta}-glucans and their receptor Dectin-1 in the induction of autoimmune arthritis in genetically susceptible mice.