Doctoral Thesis
Cantharidin world in nature:
structure and function of a concealed arthropod assemblage with interactions via cantharidin
Kosei Hashimoto
Department of Biology Tokyo Metropolitan University
Tokyo Japan
February in 2016
Contents
Abstract ---1 1. General introduction --- 3 2. Structure and seasonal dynamics of the arthropod community attracted to cantharidin --- 7
2-1. Introduction
2-2. Materials and methods 2-3. Results
2-4. Discussion
3. The arthropod community attracted to cantharidin on islands --- 25
3-1. Introduction
3-2. Materials and methods 3-3. Results
3-4. Discussion
4. Cantharidin as a nuptial gift from males to females in false blister beetles --- 37
4-1. Introduction
4-2. Materials and methods 4-3. Results
4-4. Discussion
5. General discussion --- 59
5-1. Inter-specific interactions via cantharidin
5-2. Intra-specific interactions via cantharidin
6. Acknowledgments --- 70 7. References --- 71 8. Appendices ---83
8-1. Publications
8-2. Title and summary in Japanese
Abstract
Cantharidin is produced by beetles of only two families, the Meloidae and the Oedemeridae. This substance is a defensive terpenoid that is toxic to the enemies of the beetles. Cantharidin also attracts other groups of insects and has been used as a bait to trap them. Cantharidin-baited traps deployed in central Japan captured coleopterans (Anthicidae, Endomychidae, Pyrochroidae, and Scarabaeidae), dipterans (Ceratopogonidae), hymenopterans (Formicidae), and harvestmen (Podoctidae). The seasonal occurrences of these arthropods, their sex ratios, known feeding choices, and mating habits suggested three possible underlying reasons for the attractancy of cantharidin: (i) it is accumulated as a defense against enemies and sometimes for transfer from males to females as a nuptial gift, (ii) it is used as a chemical cue in food searching, and (iii) it is used as an aggregation pheromone by mature arthropod individuals. Thus, the group of canthariphilous arthropods represented a cryptic ecological assemblage with rare chemical networking among apparently unrelated species, and this community was named “cantharidin world”.
Next, to clarify the canthariphilous arthropod community formed on small islands, they were collected by the cantharidin-baited traps and compared among the mainland, peninsula, and 11 small islands of the Izu-Ogasawara Arc, central Japan. Control traps without cantharidin were also used to collect non-canthariphilous arthropods.
Cantharidin-producing beetles were distributed across the study area, but the number of
species was reduced on islands. We identified ten species of five families as
canthariphilous arthropods in this study area; i.e., two species of Anthicidae and
Pyrochroidae (Coleoptera), four species of Ceratopogonidae (Diptera), three species of
Formicidae (Hymenoptera), and one species of Podoctidae (Opiliones). The number of
canthariphilous species was not correlated with island size (area) but negatively
correlated with the distance of islands from the mainland. These tendencies were also
seen in non-canthariphilous fauna. The canthariphilous arthropods increased with the
number of cantharidin-producing beetle species, but the non-canthariphilous fauna did
not. Thus, the cantharidin world on islands seems to be affected not only by the
geographic characteristics of islands, particularly isolatedness from the mainland, but also by the diversity of cantharidin-producing members within it.
Thirdly, to examine the nuptial gift functions of cantharidin, mating behavior and internal reproductive organs were compared among false blister beetle species (Oedemeridae). In several groups of insects, spermatophores that males transfer into the female genital tract at copulation are used as nuptial gifts because the mated females absorb nutrients, defense chemicals, or minerals from them and use to maintain their bodies and reproduction. In the morphological comparisons of female internal reproductive organs of 39 species of false blister beetles, we found that some species have sclerotized spines in the bursa copulatrix (bursal spines) but others have no such spines. The molecular phylogenetic trees inferred from mitochondrial 16S and nuclear 28S rRNA gene sequences suggested multiple origins of bursal-spine acquirement from the ancestor without spines. Males transferred a small amount of ejaculates in the spineless species, but transferred a large spermatophore in the spined species. Deposited spermatophores were finally disappeared in the bursa probably by absorption. To compare the amount of cantharidin contained in eggs laid by the species with and without bursal spines, a new bioassay system was constructed using a small beetle Mecynotarsus tenuipes of the family Anthicidae. This beetle was attracted to a droplet of cantharidin/acetone solution to feed on, and increased in number as increasing cantharidin concentration. This bioassay demonstrated that the eggs of Nacerdes caudata and N. katoi both with conspicuous bursal spines contain more cantharidin than N. waterhousei without spines. In the former species, males transfer large spermatophores to the female, and then the spermatophores are eventually broken and consumed within the female spiny bursa. Thus, females with bursal spines may be able to accumulate more cantharidin in their eggs; that is nuptial gift function of their spermatophores.
Finally, the structure and function of cantharidin world are discussed at the inter-
1. General introduction
Cantharidin (C
10H
12O
4), a monoterpene anhydride, is one of the most famous toxins from insects. This compound has been known for its various physiological activities such as vesicant, diuretic, abortifacient, aphrodisiac, and antitumoral agents in humans (Nickolls & Teare 1954; Pemberton 1999; Prestwich & Blomquist 1987; Sandori 2001;
Wang 1989; Verma & Prasad 2012). Furthermore, cantharidin inhibits protein phosphatase 2A enzymes which are most highly conserved in many different organisms such as invertebrates, vertebrates, yeast, and higher plants (Cohen 1989; Cohen et al.
1989; Li & Casida 1992; Shenolikar 1994; Luan 2003).
In nature, cantharidin is produced only by the beetles of two families Meloidae and Oedemeridae (Carrel & Eisner 1974; Carrel et al. 1986). These beetles hold the toxin in their hemolymph fluid and in their tissues (Carrel et al. 1986, 1993; Frenzel & Dettner 1994; Holz et al. 1994). Adult female meloid beetles contain cantharidin, but they are unable to synthesize it (e.g., Carrel et al. 1993). They acquire the compound from males through frequent copulation and use it subsequently for defending eggs (Nikbakhtzadeh et al. 2007, 2012). In contrast, both sexes of oedemerid beetles produce cantharidin in their bodies (Carrel et al. 1986; Frenzel & Dettner 1994), and there is little or no transfer of the substance from males to females (Holz et al. 1994). Meloid and oedemerid beetles use cantharidin through their entire lives as an antimicrobial agent against entomopathogenic fungi, as an effective feeding deterrent against predatory insects, and as a strong toxin against vertebrate enemies (e.g., Carrel & Eisner 1974;
Graziano et al. 1988; Pinetti & Biggio 1968; Blum 1996).
In contrast, several groups of insects are known to be attracted to cantharidin,
including the coleopteran families Anthicidae, Endomychidae, Staphylinidae,
Pyrochroidae, Cleridae, and Chrysomelidae, the dipteran families Ceratopogonidae,
Anthomyiidae, Cecidomyiidae, Chloropidae, Platystomatidae, and Sciaridae, the
heteropteran families Lygaeidae, Miridae, and Tingidae, and the hymenopteran families
Braconidae and Diapriidae (Hemp & Dettner 2001). Members of these groups are
recognized as canthariphilous insects (Hemp & Dettner 2001). Taxonomists often use
cantharidin to attract these species to collection traps (Young 1984; Hemp et al. 1999).
Why are they attracted to cantharidin? Although the knowledge is fragmentary, three pathways have been suggested how particular groups of insects are attracted to cantharidin. One pathway describes the use of cantharidin for defense against enemies.
Member insects either synthesize the compound or acquire it by consumption (Frenzel
& Dettner 1994; Hemp et al. 1999). Female pyrochroid and anthicid beetles obtain cantharidin by mating with males that have taken up the substance from their food; the females use cantharidin for protection of their offspring (eggs and larvae) (Eisner et al.
1996a, 1996b; Dettner 1997). The underlying premise for this nuptial gifting function is that males are strongly attracted to cantharidin. A second pathway describes the use of cantharidin as a cue in food searching (Dettner 1997). Certain complex chemical compounds are similar to cantharidin in form and function, and diverse insects searching for foods may be coincidentally attracted to cantharidin. Cantharidin has never been found in plants or fungi, but cantharidin-related compounds occur in plants.
Palasonin is one of these compounds; it lacks one of the angular methyl groups of cantharidin and has been isolated from the seeds of Butea frondosa (Leguminosae) (Bochis 1960; Raj & Kurup 1966). Higher plants in the families Meliaceae, Simaroubaceae, Apiaceae, and Aristolochiaceae may also have the potential to synthesize cantharidin itself or its analogs, such as palasonin and coumarin (Frenzel et al. 1992; Dettner 1997). Thus, considerations of the attractancy of cantharidin and its analogs must take into account the attractancy of volatile substances, such as floral scents and the aromas of mushrooms, produced by species that are food or hosts of the insects. The last pathway describes the use of cantharidin or related compounds as chemical signals in insect aggregation responses (Hemp et al. 1999).
Unfortunately, most previous empirical studies have not used controlled
experimental designs; the attraction of canthariphilous species to baited traps has not
been tested through comparisons with simultaneously set control traps without bait.
spatio-temporal patterns of all species of canthariphilous arthropods and to identify three functional groups with different types of cantharidin interactions.
The members of the cantharidin world are common among distant continents in the world (e.g., Young 1984; Hemp et al. 1999). On islands, however, only a few species of the beetle family Anthicidae were recorded as canthariphilous from Cebu, Canary, Mallorca, Cyprus, and Dominica (Hemp & Dettner 2001), and the comparisons of these arthropod communities have never been done. How does the cantharidin world change on islands? Insular fauna and flora are often disharmonious in their taxonomic composition (Carlquist 1965), and in the general island theory species richness is lower on smaller and more distant islands (MacArthur & Wilson 1963, 1967). Therefore, the arthropod communities attracted to cantharidin were compared between the mainland and islands. This is the second purpose of this study.
Males of some insect species that use defense chemicals against predators store defensive compounds as nuptial gifts to their spermatophores; for example, pyrrolizine alkaloids (Dussourd et al. 1988; Eisner & Meinwald 1995) and cyanogenic glycosides (Cardoso & Gilbert 2007). These chemicals are transferred from males to females through the copulation, and allow females to protect their eggs from predation.
Cantharidin may also be nuptial gifts in some beetles in the same context as above. This defensive compound is produced by true blister beetles (Family Meloidae) and false blister beetles (Oedemeridae) (Carrel & Eisner 1974; Carrel et al. 1986), and kept in their hemolymphs and various other tissues at larval and adult stages (Dixon et al. 1963;
Carrel et al. 1986, 1993; Frenzel & Dettner 1994; Holz et al. 1994). In true blister
beetles, however, only males continue to synthesize cantharidin after adult eclosion, and
a newly synthesized cantharidin is moved to their reproductive accessory glands, then
transported to the epididymis and vas deferens, and finally deposited and accumulated
in the testes (Nikbakhtzadeh et al. 2007). Adult females have cantharidin just
synthesized during the larval period (Sierra et al. 1975; Carrel et al. 1993). Thus, adult
females need to receive more cantharidin from male-derived spermatophores to defend
their eggs successfully. After mating, cantharidin is accumulated in the female
spermatophoral receptacle (bursa copulatrix) and allocated to their eggs (Carrel et al.
1993; Nikbakhtzadeh et al. 2007, 2012).
Contrarily, false blister beetles can synthesize cantharidin by not only adult males but also adult females (Carrel et al. 1986; Frenzel & Dettner 1994). In most cases, field-caught females contain more cantharidin than males (Frenzel & Dettner 1994;
Abtahi et al. 2012). Holz et al. (1994) reported that no or only a very small amount of
cantharidin is transferred from males to females at mating and the contribution of it as
nuptial gifts may be negligible in Oedemera femorata. Thus, it is still unclear whether
or not male false blister beetles use cantharidin as nuptial gifts to females. Therefore, in
the third part of this study, the possible function of cantharidin as a nuptial gift from
males to females was examined in false blister beetles. Finally, the structure and
function of the arthropod community with interactions via cantharidin was discussed
from not only interspecific relationships but also intraspecific relationships of such an
intersexual nuptial gifting.
2. Structure and seasonal dynamics of the arthropod community attracted to cantharidin
2-1. Introduction
Insects of many groups use chemical defenses for protection from predators and parasites. These chemicals are either synthesized by insect metabolism or acquired from larval or adult food (Bowers 1992). Cantharidin (C
10H
12O
4), a monoterpene anhydride, is one of the better known toxins produced by insects. In nature, beetles of the families Meloidae and Oedemeridae are the only producers of cantharidin (Carrel & Eisner 1974; Carrel et al. 1986). These beetles hold the toxin in their hemolymph fluid and in their tissues through their entire lives (Carrel et al. 1986, 1993; Frenzel & Dettner 1994;
Holz et al. 1994). Adult female meloid beetles contain cantharidin, but they are unable to synthesize it (e.g., Carrel et al. 1993). They acquire the compound from males through frequent copulation and use it subsequently for defending eggs (Nikbakhtzadeh et al. 2007, 2012). In contrast, both sexes of oedemerid beetles produce cantharidin in their bodies (Carrel et al. 1986; Frenzel & Dettner 1994), and there is little or no transfer of the substance from males to females (Holz et al. 1994). Meloid and oedemerid beetles use cantharidin through their entire lives as an antimicrobial agent against entomopathogenic fungi, as an effective feeding deterrent against predatory insects, and as a strong toxin against vertebrate enemies (e.g., Carrel & Eisner 1974;
Graziano et al. 1988; Pinetti & Biggio 1968; Blum 1996).
Cantharidin also serves as a powerful attractant for several insect groups, including
the coleopteran families Anthicidae, Endomychidae, Staphylinidae, Pyrochroidae,
Cleridae, and Chrysomelidae, the dipteran families Ceratopogonidae, Anthomyiidae,
Cecidomyiidae, Chloropidae, Platystomatidae, and Sciaridae, the heteropteran families
Lygaeidae, Miridae, and Tingidae, and the hymenopteran families Braconidae and
Diapriidae (Hemp & Dettner 2001). Members of these groups are recognized as
canthariphilous insects (Hemp & Dettner 2001). Taxonomists often use cantharidin to
attract these species to collection traps (Young 1984; Hemp et al. 1999).
To date, three pathways have been suggested to describe the ways in which particular groups of insects are attracted to cantharidin, but the underlying knowledge base is fragmentary. One pathway describes the use of cantharidin for defense against enemies. Member insects either synthesize the compound or acquire it by consumption (Frenzel & Dettner 1994; Hemp et al. 1999). Female pyrochroid and anthicid beetles obtain cantharidin by mating with males that have taken up the substance from their food; the females use cantharidin for protection of their offspring (eggs and larvae) (Eisner et al. 1996a, 1996b; Dettner 1997). The underlying premise for this nuptial gifting function is that males are strongly attracted to cantharidin. A second pathway describes the use of cantharidin as a cue in food searching (Dettner 1997). Certain complex chemical compounds are similar to cantharidin in form and function, and diverse insects searching for foods may be coincidentally attracted to cantharidin.
Cantharidin has never been found in plants or fungi, but cantharidin-related compounds occur in plants. Palasonin is one of these compounds; it lacks one of the angular methyl groups of cantharidin and has been isolated from the seeds of Butea frondosa (Leguminoseae) (Bochis 1960; Raj & Kurup 1966). Higher plants in the families Meliaceae, Simaroubaceae, Apiaceae, and Aristolochiaceae may also have the potential to synthesize cantharidin itself or its analogs, such as palasonin and coumarin (Frenzel et al. 1992; Dettner 1997). Thus, considerations of the attractancy of cantharidin and its analogs must take into account the attractancy of volatile substances, such as floral scents and the aromas of mushrooms, produced by species that are food or hosts of the insects. The last pathway describes the use of cantharidin or related compounds as chemical signals in insect aggregation responses (Hemp et al. 1999).
Unfortunately, most previous empirical studies have not used controlled
experimental designs; the attraction of canthariphilous species to baited traps has not
been tested through comparisons with simultaneously set control traps without bait. In
the present study, we examined a canthariphilous arthropod community in a hill forest
different types of cantharidin interactions.
2-2. Materials and methods
Trapping
Trapping was performed in a 4-ha deciduous forest at Matsugi-hinata Hill, Hachioji, Tokyo, central Japan (35°37'N, 139°22'E; altitude 120 m; 100 × 400 m
2) where the three species of cantharidin-producing oedemerid beetles, Nacerdes katoi, N. luteipennis, and N. waterhousei inhabit. In each trap, we inserted a single filter paper disk (55 mm in diameter, #2, Whatman, Little Chalfont, UK), which had been impregnated with 0.5 ml of a 10
-2M solution of cantharidin in acetone, into the base of a cylindrical plastic cup (95 mm in internal diameter, 50 mm in depth). Two square openings were cut in the cover of the plastic cup; the openings were each 10 × 10 mm
2and spaced 60 mm apart (Fig. 2-1a). We used the same procedure in setting up controls, but there was no cantharidin in the acetone used to impregnate the filter paper. On each sampling occasion, we buried five pairs of cantharidin and control traps in the forest floor (Fig.
2-1b). Paired traps were ca. 2 m apart, and the distance between pairs was about 30 m.
After 24 h, all the traps were recovered, and the openings sealed with vinyl tape to prevent loss of trapped animals. Recovered traps were kept at –20°C for later analysis.
Trappings were repeated every two weeks for 1.2 years (April 2011 to June 2012).
Analyses
All animals in the traps were carefully extracted with forceps. They were counted
and identified to the species level when possible. The attractancy of cantharidin for each
taxon was judged by binomial tests on differences in the total number of individuals
between cantharidin and control traps. Assemblage structure of the canthariphilous
fauna was described with the Shannon index H (Shannon & Weaver 1949) and the
evenness index E (Pielou 1975) calculated as below:
H = − Σ p
ilog
2p
iE = H / log
2S,
where S is the number of species trapped, and p
iis the proportion of individuals belonging the i-th species. The dominance status of each species in the assemblage was determined by a logarithmic dominance classification: eudominant when 0.32 < p
i ≤1.00, dominant when 0.10 < p
i≤ 0.32, subdominant when 0.032 < p
i≤ 0.10, recedent when 0.010 < p
i ≤ 0.032, subrecedent when 0.0032 <p
i ≤ 0.010, sporadic whenp
i ≤0.0032, and missing when p
i= 0 (Engelmann 1978).
Morphological observation
We used light microscopy to determine whether specimens attracted to the cantharidin traps were sexually dimorphic in glandular development and structure.
2-3. Results
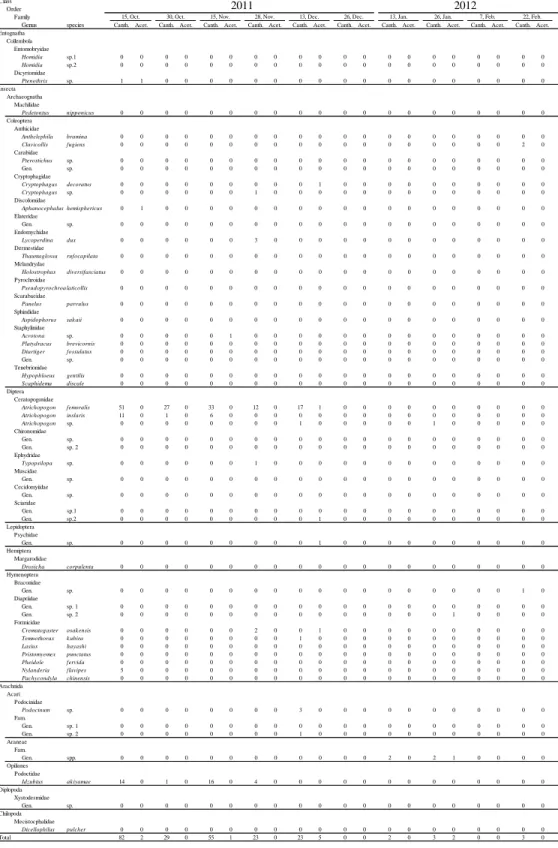
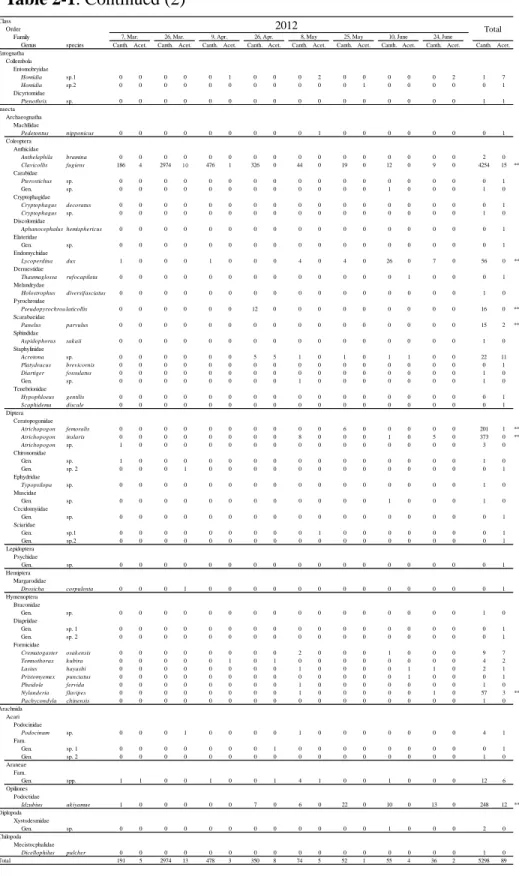
We captured >53 species of arthropods (34 families, 12 orders, five classes) in the traps during the study period (Table 2-1). Among them, eight species were significantly attracted to cantharidin: four species of the coleopteran families Anthicidae, Endomychidae, Pyrochroidae, and Scarabaeidae, two species of the dipteran family Ceratopogonidae, one species of the hymenopteran family Formicidae, and one species of the harvestman family Podoctidae.
The seasonal occurrences and sex ratios of species collected are depicted in Figure
2-2. The pyrochroid beetle Pseudopyrochroa laticollis and the anthicid beetle
Clavicollis fugiens were attracted to cantharidin only in spring, and their sex ratios were
male-biased. We collected more males than females of the endomychid beetle
Lycoperdina dux during the period from spring to summer. Both males and females of
summertime; Atrichopogon femoralis appeared mainly in spring and autumn. Ants belonging to the species Nylanderia flavipes were all workers and were trapped in seasons other than winter, although this includes one exceptional sampling of 46 individuals (Table 2-1). Both sexes of the harvestman Idzubius akiyamae were also trapped in seasons other than winter.
The diversity (H) and evenness (E) indices calculated from pooled frequencies of all canthariphilous species across all months were 1.09 and 0.36, respectively. Values of H in each of the months from April 2011 to June 2012 were 0.26, 0.63, 2.09, 1.86, 1.61, 0.97, 1.30, 1.43, 0, 0, 0, 0.01, 0.12, 1.79, and 1.89, respectively; over the same time period, values of E were 0.26, 0.24, 0.74, 0.80, 0.70, 0.49, 0.65, 0.71, 0, 0, 0, 0.01, 0.08, 0.69, and 0.81, respectively. Thus, diversity tended to increase from spring to summer, decreasing thereafter (Fig. 2-3). Clavicollis fugiens was eudominant, two species of Atrichopogon and Idzubius akiyamae were subdominant, Lycoperdina dux and Nylanderia flavipes were recedent, and Pseudopyrochroa laticollis and Panelus parvulus were sporadic members of the canthariphilous fauna.
Males of Pseudopyrochroa laticollis had cephalic glands, but females did not (Fig.
2-4a, b). Clavicollis fugiens was sexually dimorphic in elytral structure: males had a pair of remarkable glandular notches, but the females did not (Fig. 2-4c, d). Both male and female Idzubius akiyamae had a single pair of exocrine glands on anterolateral margins of the carapace, located approximately between the second and third walking legs (Fig. 2-4e–h).
2-4. Discussion
The canthariphilous assemblage collected in central Japan comprised the coleopteran families Anthicidae, Endomychidae, Pyrochroidae, and Scarabaeidae, the dipteran family Ceratopogonidae, the hymenopteran family Formicidae, and the harvestman family Podoctidae. Common insect groups have been collected by cantharidin traps across different geographic regions and habitats (e.g., Young 1984;
Hemp et al. 1999), indicating that this assemblage is readily extracted from the biota by
using cantharidin traps. The harvestman Idzubius akiyamae was clearly attracted to cantharidin in our study; this is a first record of this group of arthropods in the canthariphilous fauna. Idzubius akiyamae had secretory glands similar to those of other harvestmen (e.g., Duffield et al. 1981), viz., there was a pair of exocrine glands on the anterolateral margins of the carapace located approximately between the second and third pairs of walking legs. Clear droplets exuded slowly from these glands. These secretions may well have functioned as a chemical defense against enemies (e.g., Eisner et al. 1971). However, studies of glandular construction in Trogulus species suggest functions other than defense, such as territorial marking, aggregation, and sexual communication (Schaider & Raspotnig 2009). Harvestmen have aggregation behavior that likely contributes to defense, avoidance of water loss, and/or mating success;
however, the adaptive significance of this behavior remains unknown (Machado &
Macías-Ordóñez 2007). Harvestmen form both multispecies and unispecific aggregations (e.g., Machado & Vasconcelos 1998; Chelini et al. 2012). The multispecies aggregation may increase the defense potential against predators. If the podoctid harvestmen use cantharidin or related compounds similarly, formation of multispecies aggregates would probably occur. The only species of harvestmen that we collected was Idzubius akiyamae, which might use cantharidin as an aggregation pheromone (Fig. 2-5).
Another possibility that they are attracted to cantharidin is their usage as a cue in food searching. However, this may be denied because any other harvestmen were not attracted to cantharidin even though their food habits were similar to those Idzubius akiyamae (Acosta & Machado 2007).
Defensive use of ingested cantharidin occurs in some canthariphilous insects.
Pyrochroid beetles are attracted to and feed on synthetic cantharidin and on meloid
beetles, which contain cantharidin. Since 1827, canthariphily has been reported in 29
pyrochroid species of the genera Anisotoria, Pedilus, Neopyrochroa, Pyrochroa, and
Schizotus (Hemp & Dettner 2001). Holz et al. (1994) first reported that males of
flabellata. Eggs are effectively protected from insectivorous predators by the toxicity of cantharidin (Eisner et al. 1996b). Eisner et al. (1996a) demonstrated that cantharidin in the male functions as a close-range excitatory pheromone for females before copulation occurs. Male Neopyrochroa flabellata have a glandular cleft on the head, which also occurs as in many other pyrochroids (elytral notches in Pedilus species). The males secrete stored cantharidin from the cleft when meeting a potential mate. A female bites the cleft to confirm the presence of cantharidin in the male. Females preferentially mate with males with cantharidin, probably to ensure a higher survival rate in their offspring.
Thus, cantharidin may be used for self-protection but also for offspring protection via nuptial gifting, which may well be a sexually selected male trait. We found that the pyrochroid beetle Pseudopyrochroa laticollis is attracted to cantharidin and that cephalic glands occur only in males. Thus, cantharidin eaten by the male may be used as a nuptial gift, as in Schizotus pectinicornis and Neopyrochroa flabellata (Fig. 2-5).
Strong attraction to cantharidin occurs in anthicid beetles. Görnitz (1937) first demonstrated that the anthicid species Notoxus monoceros is canthariphilous. Since then, canthariphily has been reported in more than 190 species of Anthicidae (Hemp &
Dettner 2001). Males sequester considerable amounts of cantharidin in elytral notches, testes, and accessory glands. Females, however, are little or never attracted to cantharidin (Schütz & Dettner 1992). Paired male elytral notches function in cantharidin secretion in the same way as glandular clefts in pyrochroid beetles other than species of Anthelephila (Schütz & Dettner 1992). We found that Clavicollis fugiens, mostly males, were readily trapped with cantharidin bait. The species was sexually dimorphic in elytral structure; males possessed pairs of remarkable glandular notches, but females did not. Clavicollis fugiens may therefore use cantharidin to defend offspring via nuptial gifting, as the case in pyrochroid beetles (Fig. 2-5).
The cues for food searching are diverse in insects. Some insect groups may be attracted to cantharidin or chemically similar substances if the species use these compounds as food searching cues. The following groups fit this pattern (Fig. 2-5).
Beetles of the family Endomychidae include five canthariphilous species in four genera:
Aphorista, Danae, Lycoperdina, and Xenomycetes (Young 1984, 1989). We found that
Lycoperdina dux was significantly attracted to cantharidin. Insects in this group are mycophagous, and among them, members of the genera Endomychus, Xenomycetes, Mycetina, Lycoperdina, and Aphorista have obligate relationships with lignicolous Basidyomycetes (Tomaszewska 2010). For example, beetles in the genus Lycoperdina feed on puffballs of the genus Lycoperdon (Pakaluk 1984); beetles of the species Xenomycetes laversi graze on the basidiomycete Paxilus atrotomentosus (Johnson 1986).
Lycoperdina dux also feeds on the puffball species Calvatia craniiformis (Taniguchi 1942). Endomychid beetles often appear to aggregate on their host mushrooms, but this is not true aggregation; it is merely a dense conglomeration of individuals on sparse, patchily distributed host fungi (Shockley et al. 2009). Basidiomycete fruiting bodies release volatiles including some of monoterpene/lactone compounds (Breheret et al.
1997). The aroma of the puffball Calvatia gigantea includes some of monoterpene/lactone compounds (Leffingwell & Alford 2011). Cantharidin is one of them. Therefore, endomychids may be attracted to cantharidin, whose odor resembles that of their host mushrooms.
There is no available information on the function of cantharidin as an attractant for dung beetles (family Scarabaeidae). In our study, male and female Panelus parvulus were attracted to the cantharidin bait. This beetle feeds on feces and uses them for brooding. Fresh feces of vertebrates emit volatiles belonging to the monoterpene/lactone group (e.g., Dormont et al. 2010). It is therefore likely that Panelus parvulus is attracted to cantharidin, which has stimulatory activities similar to those of cantharidin analogs emitted from vertebrate feces.
Adult female biting midges of the family Ceratopogonidae suck hemolymph from
their hosts but males do not. We collected two species of Atrichopogon (subfamily
Forcipomyiinae) in our cantharidin traps. To date, cantharidin is known to attract more
than 20 species of biting midges (Hemp & Dettner 2001). Most of them are those
members of the genus Atrichopogon that suck hemolymph from cantharidin-producing
Atrichopogon. On the other hand, Atrichopogon midges sometimes visit flowers of the Apiaceae and Aristolochiaceae (Frenzel et al. 1992). It is difficult to decide at the present time whether flower-visiting behavior in these midges is normal or simply an artifact explainable by the chemical similarities between plant-derived compounds such as pyrano- and furano-coumarins and cantharidin (Frenzel et al. 1992). Nevertheless, some of the forcipomiine biting midges are clearly important pollinators of cocoa (Theobroma cacao) and rubber trees (Hevea brasiliensis) (Wirth 1956; Winder 1977;
Young 1982). Furthermore, the forcipomiine species Forcipomyia opilionivora sucks fluids from an unidentified harvestman species (Phalangidae gen. sp.) (Lane 1947). If harvestmen release aggregation pheromones, including cantharidin or its analogs, as mentioned above, Atrichopogon species may be attracted to cantharidin and suck out the harvestmen hemolymph. Thus, cantharidin and its analogs may be responsible for unpredictable interspecific interactions between pairs of apparently unrelated species, e.g., midge–plant and midge–harvestman relationships.
We demonstrated that workers of the ant species Nylanderia flavipes were attracted to cantharidin. Cantharidin is a potent feeding deterrent for ants (Carrel & Eisner 1974), which are the most common floral visitors contributing to plant pollination (Beattie 2006). The species of Nylanderia are also responsible for pollination in some plants species (e.g., Kawakita & Kato 2002). Animals are attracted to flowers by diverse sets of visual (floral color and shape) and chemical (floral scent and nectar features) cues (de Vega 2009). Cantharidin-related compounds also occur in some plants (Frenzel et al.
1992). Thus, the attractancy of cantharidin for ants may be an artifact of the natural use of plant-produced cantharidin analogs as chemical cues for ants searching for flowers.
Our study revealed a new type of canthariphilous arthropod assemblage that includes harvestmen. Our premise is that some groups of arthropods are attracted to cantharidin because (i) the compound may be accumulated for defense against enemies or transferred as a nuptial gift from males to females; (ii) cantharidin may be used as a chemical cue in food searching; and (iii) it functions as an aggregation pheromone.
Within this arthropod assemblage, there may be further unknown natural ecological
interactions between apparently unrelated species, particularly in species using
cantharidin in food searching, e.g., in midge–plant, midge–harvestman, and ant–plant
relationships (Fig. 2-5). Undetected interactions may include preadaptations leading to
the evolution of new pollination systems and the acquisition of new hosts.
Table 2-1. The number of arthropods collected by the cantharidin and control
traps set semimonthly in a forest floor of the hill. *P < 0.05, **P < 0.01 in the exact binominal tests for the total number of individuals collected. Canth., the number of each species collected by the cantharidin traps. Acet., the number of each species collected by the control traps using only acetone.
Class Order
Family
Genus species Canth. Acet. Canth. Acet. Canth. Acet. Canth. Acet. Canth. Acet. Canth. Acet. Canth. Acet. Canth. Acet. Canth. Acet. Canth. Acet. Canth. Acet.
Entognatha Collembola
Entomobryidae
Homidia sp.1 0 0 0 0 1 0 0 0 0 2 0 0 0 0 0 0 0 0 0 0 0 0
Homidia sp.2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Dicyrtomidae
Ptenothrix sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Insecta Archaeognatha
Machilidae
Pedetontus nipponicus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Coleoptera Anthicidae
Anthelephila bramina 0 0 2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Clavicollis fugiens 79 0 40 0 52 0 18 0 17 0 0 0 0 0 0 0 0 0 0 0 0 0
Carabidae
Pterostichus sp. 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0
Gen. sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cryptophagidae
Cryptophagus decoratus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cryptophagus sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Discolomidae
Aphanocephalus hemisphericus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Elateridae
Gen. sp. 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Endomychidae
Lycoperdina dux 0 0 0 0 1 0 0 0 5 0 4 0 0 0 0 0 0 0 0 0 0 0
Dermestidae
Thaumaglossa rufocapilata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Melandrydae
Holostrophus diversifasciatus 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pyrochroidae
Pseudopyrochroa laticollis 2 0 2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Scarabaeidae
Panelus parvulus 0 0 0 0 0 0 2 1 2 0 0 0 0 0 8 0 3 0 1 1 0 0
Sphindidae
Aspidophorus sakaii 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0
Staphylinidae
Acrotona sp. 0 0 0 0 0 0 1 1 12 3 1 0 0 0 0 0 0 0 0 0 0 0
Platydracus brevicornis 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0
Diartiger fossulatus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0
Gen. sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Tenebrionidae
Hypophloeus gentilis 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Scaphidema discale 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Diptera Ceratopogonidae
Atrichopogon femoralis 0 0 0 0 1 0 11 0 0 0 34 0 2 0 3 0 2 0 0 0 2 0
Atrichopogon inslaris 0 0 0 0 2 0 1 0 0 0 20 0 9 0 142 0 36 0 125 0 6 0
Atrichopogon sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Chironomidae
Gen. sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Gen. sp. 2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Ephydridae
Typopsilopa sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Muscidae
Gen. sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cecidomyiidae
Gen. sp. 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Sciaridae
Gen. sp.1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Gen. sp.2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lepidoptera Psychidae
Gen. sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Hemiptera Margarodidae
Drosicha corpulenta 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Hymenoptera Braconidae
Gen. sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Diapriidae
Gen. sp. 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Gen. sp. 2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Formicidae
Crematogaster osakensis 0 0 0 0 0 5 1 0 0 0 1 0 0 0 0 0 0 0 0 1 2 0
Temnothorax kubira 0 0 0 0 1 0 0 0 0 0 0 0 2 0 0 0 0 0 0 0 0 0
Lasius hayashi 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pristomyemex punctatus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pheidole fervida 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Nylanderia flavipes 0 0 0 0 0 2 2 0 0 0 1 0 1 1 46 0 0 0 0 0 0 0
Pachycondyla chinensis 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0
Arachnida Acari
Podocinidae
Podocinum sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Fam.
Gen. sp. 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Gen. sp. 2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Araneae Fam.
Gen. spp. 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 1
Opiliones Podoctidae
Idzubius akiyamae 1 0 1 0 2 0 4 0 7 0 11 1 13 6 8 0 57 3 37 2 13 0
Diplopoda Xystodesmidae
Gen. sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0
Chilopoda Mecistocphalidae
Dicellophilus pulcher 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Total 83 2 45 1 61 8 40 3 44 5 72 2 27 8 208 1 99 3 163 4 25 1
2011
29, Apr. 6, May 20, May 5, June 23, June 7, July 25, July 7, Aug 29, Aug. 8, Sep. 28, Sep.