Rapid Communication 1
2
SGLT2 inhibitor ipragliflozin attenuates breast cancer cell proliferation.
3 4
Shiho Komatsu
1, Takashi Nomiyama
1,4 *, Tomohiro Numata
2, Takako Kawanami
1, 5
Yuriko Hamaguchi
1, Chikayo Iwaya
3, Tsuyoshi Horikawa
1, Yuki Fujimura-Tanaka
1, 6
Nobuya Hamanoue
1, Ryoko Motonaga
1, Makito Tanabe
1, Ryuji Inoue
2, Toshihiko 7
Yanase
3,4, Daiji Kawanami
18
9
1
Department of Endocrinology and Diabetes Mellitus, School of Medicine, Fukuoka 10
University, Fukuoka, Japan 11
12
2
Department of Physiology, School of Medicine, Fukuoka University, Fukuoka, Japan 13
14
3
Muta Hospital, Fukuoka Japan 15
16
4
Research institute for Islet Biology, Fukuoka University, Fukuoka, Japan 17
18
*Correspondence:
19
Takashi Nomiyama MD, PhD 20
Department of Endocrinology and Diabetes Mellitus 21
School of Medicine, Fukuoka University, 7-45-1 Nanakuma, Jonan-ku, Fukuoka 814- 22
0180, Japan 23
[email protected] 24
25
Running Head: SGLT2 inhibitor on breast cancer cell 26
Key words: SGLT2 inhibitor, breast cancer, membrane potential, mitochondria 27
28
ABSTRACT 29
Currently, cancer is one of the major causes of death in patients with type 2 diabetes 30
mellitus. We have previously reported the anti-prostate and anti-breast cancer effect of 31
glucagon-like peptide-1 receptor agonist exendin-4. In the present study, we examined 32
the anti-cancer effect of SGLT2 (sodium-glucose cotransporter 2) inhibitor ipragliflozin 33
using a breast cancer model. In human breast cancer cell line MCF-7 cell, SGLT2 34
expression was detected using both RT-PCR and immunohistochemistry. 1-50nM 35
ipragliflozin significantly and dose-dependently suppressed the growth curve of MCF-7 36
cells. BrdU assay revealed that ipragliflozin attenuates the proliferation rate of MCF-7 37
cell in a dose dependent manner. Because the anti-breast cancer effect of ipragliflozin 38
was completely canceled by knocking down of SGLT2, this effect could be induced by 39
SGLT2 inhibition by ipragliflozin. We next measured membrane potential and whole cell 40
current using the patch clamp technique. When we treated MCF-7 cell with ipragliflozin 41
or glucose free medium, membrane hyperpolarization was observed. In addition, glucose 42
free medium and knock-down of SGLT2 by siRNA suppressed glucose induced whole 43
cell current of MCF-7 cell, suggesting that ipragliflozin inhibits sodium and glucose 44
cotransport through SGLT2. Further, JC-1 fluorescence was significantly increased, 45
suggesting the change of mitochondrial membrane potential. These data suggest that
46
SGLT2 inhibitor ipragliflozin attenuates breast cancer cell proliferation via membrane 47
hyperpolarization and mitochondrial membrane instability.
48
49
INTRODUCTION 50
Recently, cancer is emerging as a major cause of death in patients with diabetes mellitus 51
[1]. Especially in Japan, cancer is the leading cause of death in patients with T2DM (type 52
2 diabetes mellitus). Accordingly, the Japan Diabetes Society and Japan Cancer 53
Association have therefore issued a warning regarding increased cancer risk in patients 54
with diabetes mellitus [2]. Notably, T2DM and metabolic syndrome which are caused by 55
obesity have been suggested to be associated with a higher risk of many cancers [3].
56
Based on these evidences, therapeutic strategy for T2DM which could decrease not only 57
glucose level but also cancer risk and progression should be required. In our previous 58
reports, we have investigated anti-cancer effect of GLP-1 (glucagon-like peptide-1) 59
receptor agonist using prostate cancer [4] and breast cancer models [5]. In addition, 60
further reduction of prostate cancer growth was observed by combined therapy with 61
metformin and GLP-1 receptor agonist [6].
62
On the other hand, SGLT2 (sodium-glucose cotransporter 2) inhibitor is an anti-diabetic 63
agent currently permitted in clinical application. Because of its unique glucose lowering 64
mechanism and cardiovascular protective effect, much attention have been focused on 65
SGLT2 inhibitor, recently. SGLT2 inhibitor ipragliflozin is made in Japan [7] and was 66
the first SGLT2 inhibitor enter clinical use in Japan. We have previously reported that
67
ipragliflozin increased adiponectin and HDL-cholesterol, and decreased HbA1c, body 68
mass index, serum C-peptide level and blood pressure in Japanese patients with T2DM 69
[8]. In the present study, we examined anti-cancer effect of SGLT2 inhibitor ipragliflozin 70
using breast cancer cell.
71 72
MATERIALS AND METHODS 73
Cell culture and cell proliferation assays 74
The MCF-7 and MDA-MB-231 human breast cancer cell lines were purchased from the 75
American Type Culture Collection (Manassas, VA, USA). The KPL-1 human breast 76
cancer cell line was kindly provided by Dr. Junichi Kurebayashi, Kawasaki Medical 77
School [9]. All breast cancer cells were maintained in DMEM (Dulbecco’s Modified 78
Eagle’s Medium), and all media were supplemented with 10% fetal bovine serum (FBS) 79
and 1% penicillin/streptomycin. Cell proliferation assays were performed as described 80
previously [4, 5, 6] with minor modifications. Briefly, cells were seeded in 12-well 81
tissue culture plates and maintained in complete media with or without 1–50 M 82
ipragliflozin (kindly provided by Astellas Pharma Inc., Tokyo, Japan). Cell proliferation 83
was analyzed after 0–4 days after by cell counting using a hemocytometer.
84
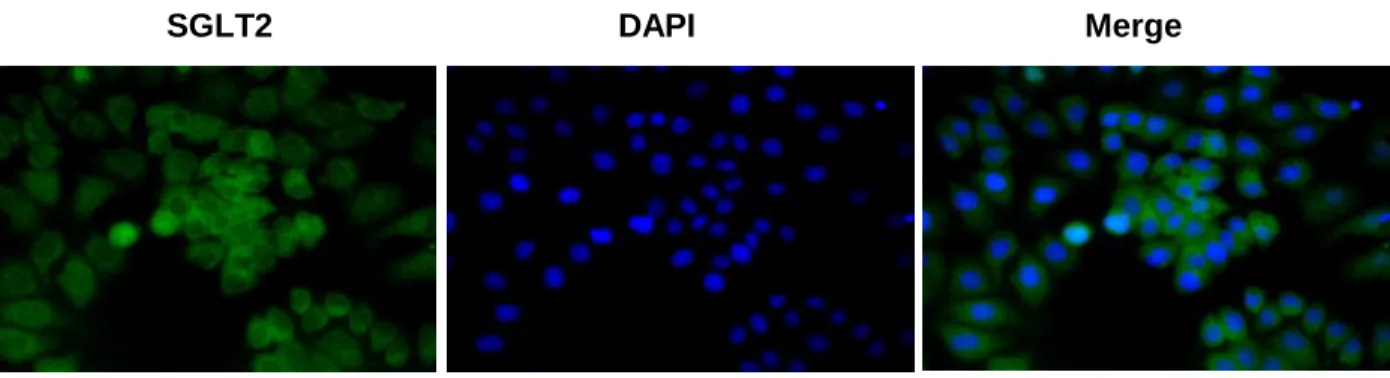
Immunohistochemistry
85
Paraffin sections were incubated with anti-SGLT2 (ab37296; Abcam, Cambridge, UK) 86
and subsequently incubated with Alexa Fluor 488 goat anti-rabbit IgG (A-11008, 87
Thermo Fisher Scientific, Rockford, IL, USA). Sections were counterstained with 4',6- 88
diamidino-2-phenylindole (DAPI) and visualized with confocal microscopy.
89
RT and quantitative real-time PCR 90
RT (Reverse Transcription) and quantitative real-time PCR (Polymerase Chain Reaction) 91
were performed as described previously [4, 6]. Total mRNA from breast cancer cells was 92
isolated using RNeasy Mini Kits (Qiagen, Venlo, the Netherlands) and reverse- 93
transcribed into cDNA. PCR reactions were performed using a Light Cycler 2.0 (Roche, 94
Basel, Switzerland) and SYBR Premix Ex Taq™ II (Takara, Otsu, Japan). Each sample 95
was analyzed in triplicate and normalized against TATA-binding protein (TBP) mRNA 96
expression. The primer sequences used were as follows: human TBP, 5′- 97
TGCTGCGGTAATCATGAGGATA-3′ (forward), 5′-
98
TGAAGTCCAAGAACTTAGCTGGAA-3′ (reverse); human SGLT2, 5′- 99
TGCATCTGATTGGCAGTCAC-3′ (forward), 5′- TTTTTGGACAGGGGAAAGGC -3′
100
(reverse). PCR products were separated by agarose gel electrophoresis and visualized 101
with ethidium bromide staining.
102
BrdU (Bromodeoxyuridine) assays
103
To evaluate cell proliferation in breast cancer cells, the BrdU incorporation assay was 104
performed using Cell Proliferation ELISA kits (1647229; Roche Applied Science, 105
Mannheim, Germany). Briefly, MCF-7 cells were plated at 5000 cells/well in 96-well 106
culture plates in complete media. After attaining 60%–70% confluence, cells were 107
treated with media containing 10% FBS with or without ipragliflozin (1–100 M) for 24 108
h. BrdU solution (10 μM) was added during the last 2 h of stimulation. Next, the cells 109
were dried and fixed, and the cellular DNA was denatured with FixDenat solution 110
(Roche Applied Science) for 30 min at room temperature. A peroxidase-conjugated 111
mouse anti-BrdU monoclonal antibody (Roche Applied Science) was added to the 112
culture plates and the cells were incubated for 90 min at room temperature. Finally, 113
tetramethylbenzidine substrate was added and the plates were incubated for 15 min at 114
room temperature, and the absorbance of the samples was measured using a microplate 115
reader at 450–620 nm. Mean data are expressed as a ratio to control (non-treated) cell 116
proliferation.
117
Small interfering (si)RNA knockdown of SGLT2 expression and cell proliferation 118
assay 119
To knockdown SGLT2, we used SGLT-2 siRNA (sc-106547; Santa Cruz Biotechnology, 120
CA, USA), which was designed to target human SGLT2, and control siRNA sc-37007
121
(sc-106547; Santa Cruz Biotechnology, CA, USA) were used as a negative control. For 122
transfection, MCF-7 cells were plated at 2 × 10
5cells/well in 6-well plates and transfected 123
with 10 nmol/l of siRNA targeting SGLT2 or negative control siRNA using MISSION 124
siRNA Transfection Reagent (Sigma-Aldrich). Seventy-two hours after transfection, cells 125
were subjected to the cell proliferation assay. Briefly, cells were detached and re-plated 126
in 24-well tissue culture plates in complete media with or without 10 M ipragliflozin. 0- 127
4 days after treatment, cells were collected and counted using a hemocytometer. The 128
siRNA knockdown efficiency was confirmed by RT-PCR of SGLT2.
129
Patch clamp measurements 130
Whole cell patch recording for current and voltage clamps were recorded using the 131
nystatin-performed patch technique in MCF-7 cell at room temperature (22-25℃) with 132
an Axopatch 200B (Molecular Devices, Sunnyvale, CA) patch-clamp amplifier as 133
describe previously [10]. For whole cell recordings, the Na
+-based bath solutions 134
contained (in mM) 140 NaCl, 5 KCl, 2 CaCl
2, 1 MgCl
2, 10 HEPES, and 10 D-glucose 135
(pH adjusted to 7.4 with NaOH and osmolality adjusted to 320 mosmol/kg-H
2O with D- 136
mannitol). The pipette solution contained (in mM) 55K
2SO
4, 20 KCl, 5 MgCl
2, 0.2 EGTA, 137
and 5 HEPES (pH adjusted to 7.4 with KOH and osmolality adjusted to 300 mosmol/kg- 138
H
2O with D-mannitol). For Figure 3B, ramp pulses (-80mV- +60mV, 0.28V/s) were
139
applied every 10 s from a holding potential of +40mV.
140
Mitochondrial permeability potential 141
Mitochondrial membrane (m) was examined using JC-1 mitochondrial membrane 142
potential detection kit (#10009172; Cayman Chemicals, Ann Arbor, MI) according to the 143
company’s instructions. MCF-7 cells treated with or without 10M ipragliflozin were 144
stained with the cationic dye JC-1, which exhibits potential-dependent accumulation in 145
mitochondria. At low membrane potential, JC-1 continues to exist as a monomer and 146
produces a green fluorescence (emission at 527 nm). At high membrane potential and 147
polarization, JC-1 forms J aggregates (emission at 590nm) and produces a red 148
fluorescence.
149
Statistical analysis 150
Unpaired t-tests and two-way ANOVA for repeated measures were performed for 151
statistical analysis as appropriate. P-values less than 0.05 were considered statistically 152
significant. Results are expressed as mean ± SEM.
153 154
RESULTS 155
SGLT2 is expressed in human breast cancer cells
156
To detect the SGLT2 expression on human breast cancer cell, we demonstrated 157
immunohistochemistry. As depicted in Figure 1A, SGLT2 was expressed on the breast 158
cancer cell membrane. Using three types of breast cancer cell lines, such as highly 159
estrogen-sensitive MCF-7 cell, estrogen-independent MDA-MB-231 cell and low 160
estrogen-sensitive KPL-1 cell, quantitative PCR confirmed SGLT2 gene expression 161
(Figure 1B). Compared with KPL-1 cell, significantly higher SGLT2 gene expression was 162
detected in MCF-7 cell. Then, we performed following experiments using MCF-7 cell, 163
which is one of the most popular human breast cancer cell line.
164
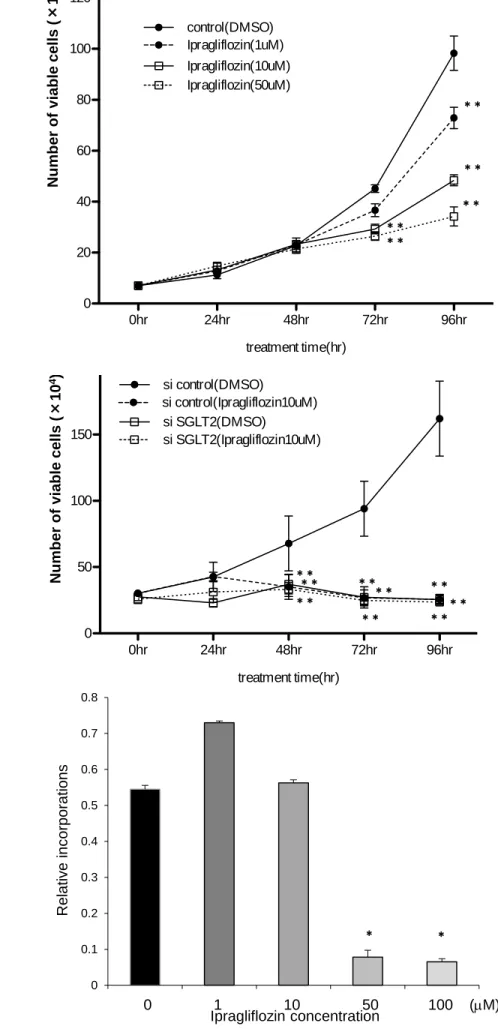
SGLT2 inhibitor attenuated breast cancer cell proliferation 165
We next treated MCF-7 cell with 0-50M ipragliflozin and drew growth curve. As shown 166
in Figure 2A, ipragliflozin decreased cell number of MCF-7 cell in growth curve in a dose 167
dependent manner. If we knocked down SGLT2 expression using siRNA, the attenuation 168
of cell proliferation induced by ipragliflozin was completely cancelled (Figure 2B), 169
suggesting that ipragliflozin attenuated breast cancer cell proliferation through SGLT2 170
inhibition. Further, BrdU assay revealed that high dose ipragliflozin inhibited DNA 171
synthesis of MCF-7 cell significantly (Figure 2C). The efficacy of knock down by siRNA 172
was confirmed by RT-PCR (data not shown).
173
SGLT2 inhibitor ipragliflozin induced hyperpolarization of MCF-7 cell membrane
174
Because SGLT2 uptake not only glucose but also sodium into cytoplasm, we next 175
examined membrane potential using patch clamp technique. As shown in Figure 3A, 176
10mM ipragliflozin induced hyperpolarization of MCF-7 cell membrane similar to 177
treatment with glucose free medium. The measurement of V revealed the significant 178
reduction of membrane potential by glucose free medium and ipragliflozin (Figure 3A), 179
suggesting that the inhibition of glucose uptake through SGLT2 induced 180
hyperpolarization of MCF-7 cell membrane potential. To confirm the pivotal role of 181
SGLT2 in ipragliflozin-induced membrane hyperpolarization, we knocked down SGLT2 182
using siRNA, and we treated MCF-7 cell with medium with or without glucose. As shown 183
in Figure 3B, the difference of membrane potential induced by deletion of glucose in 184
cultured medium, was abolished by siSGLT2.
185
GLT2 inhibitor ipragliflozin induced mitochondrial membrane instability.
186
We next examined the effect of ipragliflozin on mitochondrial membrane potential, 187
because mitochondria are one of the most important intracellular organelle deciding cell 188
death and proliferation. JC-1 dye is an indicator of cell viability, measuring mitochondrial 189
potential. Red fluorescence indicates healthy and intact mitochondria, and green 190
fluorescence indicates poor healthy mitochondria and cells going die, necrosis or 191
apoptosis. As depicted in Figure 4A, much more green fluorescence was observed in
192
MCF-7 cell treated with ipragliflozin. Further, the plotting of area rate of JC-1 193
fluorescence, red or green, revealed that mitochondrial membrane instability was induced 194
at early phase of ipragliflozin treatment, Figure 4B.
195 196
DISCUSSION 197
In the present study, we investigated anti-breast cancer effect of SGLT2 inhibitor 198
ipragliflozin through cell membrane hyperpolarization and mitochondrial membrane 199
instability. SGLT2 inhibitor is newly identified anti-diabetic agent taking much attention 200
for its glucose lowering effect without body weight gain and hypoglycemia and 201
cardiovascular protective effect. On the other hand, current basic experimental reports 202
suggested that anti-cancer effect of SGLT2 inhibitor, such as pancreatic, prostate [11], 203
liver [12] and colon cancers [13]. However, there is no report which examined the effect 204
of SGLT2 inhibitor on breast cancer. Breast cancer is one of the most critical cancer 205
related to T2DM and obesity. Further, recently published data based on National Health 206
Interview Survey in USA, suggested that 65.3% decline of mortality was observed in 207
breast cancer patients with diabetes compared with patients with no diabetes [14].
208
Accordingly, glycemic control inhibiting breast cancer progression is important for 209
female patients with DM.
210
In the present study, we investigated SGLT2 expression in human breast cancer cell, and 211
SGLT2 inhibitor ipragliflozin attenuated breast cancer cell proliferation and DNA 212
synthesis (Figure 1). The dose of ipragliflozin which attenuated breast cancer cell 213
proliferation, 1-10M, was almost similar to pharmacological serum concentrations [14], 214
suggesting that our data are not so far from clinical conditions. In BrdU assay, high dose, 215
50-100M, ipragliflozin reduced DNA synthesis (Figure 2D), however, growth curve was 216
suppressed with lower dose of ipragliflozin (Figure 2A). These data suggested that 217
ipragliflozin attenuated breast cancer cell proliferation through not only inhibiting DNA 218
synthesis but also other mechanisms, such as cell death including apoptosis. Further 219
experiments are required. We focused on sodium transport of SGLT2, because sodium 220
uptake is emerging mechanism of cancer biology including breast cancer [15]. As 221
expected, ipragliflozin shut down sodium up take through SGLT2 and subsequently 222
induced membrane hyperpolarization of MCF-7 cell. In addition, we investigated that 223
ipragliflozin induced instability of mitochondrial membrane potential which may lead to 224
apoptosis and necrosis of host cell. Hopefully, further experiments may reveal other 225
effects of SGLT2 inhibitor on cancer cells.
226
In conclusion, we investigated that SGLT2 inhibitor ipragliflozin attenuates breast 227
cancer cell proliferation via membrane hyperpolarization and mitochondrial membrane 228
instability.
229 230
ACKNOWLEDGEMENTS 231
S.K. and T.K. performed experiments and data analysis. T.No. wrote the manuscript and 232
conceived the research hypothesis and design. T.Nu. performed patch clam measurement 233
and wrote manuscript. Y.H., C.I., T.H., Y.FT., N.H. R.M., M.T., R.I. and D.K. reviewed 234
the manuscript. T.Y. conceived the research design and reviewed the manuscript.
235 236
DISCLOSURE 237
This study was supported by a research grant from Astella Pharma. T.N. received lecture 238
fees from Eli Lilly Japan, MDS, Nippon Boehringer Ingelheim, Novartis Pharma, 239
Sumitomo Dainippon Pharma, Takeda Pharmaceutical, Mitsubishi Tanabe Pharma and 240
Ono Pharmaceutical and research grants from Astellas Pharma, Eli Lilly Japan, 241
Sumitomo Dainippon Pharma, LifeScan Japan and Terumo. D.S. received lecture fees 242
from MSD and AstraZeneca. T.Y. received research grants from Sumitomo Dainippon 243
Pharma, Astellas Pharma, Eli Lilly Japan, Ono Pharmaceutical and Mitsubishi Tanabe
244
Pharma and an endowed chair with MSD, Takeda Pharmaceutical and Nippon Boehringer 245
Ingelheim. D.K. received lecture fees from Takeda Pharmaceutical.
246 247
REFERENCES 248
1. Emerging Risk Factors Collaboration, Seshasai SR, Kaptoqe S, Thompson A, Di 249
Angelantonio E, Gao P, et al. (2011) Diabetes mellitus, fasting glucose, and risk of 250
cause-specific death. N Engl J Med 364:829-841.
251
2. Kasuga M, Ueki K, Tajima N, Noda M, Ohashi K, et al. (2013) Report of the 252
JDS/JCA joint committee on diabetes and cancer. Diabetol Int 4:81-96.
253
3. Esposito K, Chiodini P, Colao A, Lenzi A, Giugliano D. (2012) Metabolic syndrome 254
and risk of cancer: a systemic review and meta-analysis. Diabetes Care 35:2402- 255
2411.
256
4. Nomiyama T, Kawanami T, Irie S, Hamaguchi Y, Terawaki Y, et al. (2014) Exendin- 257
4, a glicagon-like peptide-1 receptor agonist, attenuates prostate cancer growth.
258
Diabetes 63:3891-3905.
259
5. Iwaya C, Nomiyama T, Komatsu S, Kawanami T, Tsutsumi Y, et al. (2017) Exendin- 260
4, a glucagonlike peptide-1 receptor agonist, attenuates breast cancer growth by 261
inhibiting NF-kB activation. Endocrinology 158:4218-4232.
262
6. Tsutsumi Y, Nomiyama T, Kawanami T, Hamagichi Y, Terawaki Y, et al. (2015) 263
Combined treatment with Exendin-4 and metformin attenuates prostate cancer 264
growth. PLoS ONE 10:e0139709 265
7. Tahara A, Kurosaki E, Yokono M, Yamajuku D, Kihara R, et al. (2012) 266
Pharmacological profile of ipragliflozin (ASP1941), a novel selective SGLT2 267
inhibitor, in vitro and in vivo. Naunyn Schmiedebergs Arch Pharmacol 385:423-436.
268
8. Nomiyama T, Shimono D, Horikawa T, Fujimura Y, Ohsako T, et al. (2018) Efficacy 269
and safety of sodium-glucose cotransporter 2 inhibitor ipragliflozin on glycemic 270
control and cardiovascular parameters in Japanese patients with type 2 diabetes 271
mellitus; Fukuoka Study of Ipragliflozin (FUSION). Endocr J 65:859-867.
272
9. Kurebayashi J, Kurosumi M, Sonoo H. (1995) A new human breast cancer cell line, 273
KPL-1 secretes tumor-associated antigens and grows rapidly in female athymic nude 274
mice. Br J Cancer 71:845-853.
275
10. Numata T, Murakami T, Kawashima F, Moroue N, Heuser JF, et al. (2012) 276
Utilization of photoinduced charge-separated state of donor-acceptor-linked 277
molecules for regulation of cell membrane potential and ion transport. J Am Chem 278
Soc 134:6092-6095.
279
11. Scafoglio C, Hirayama BA, Kepe V, Liu J, Ghezzi C, et al. (2015) Functional 280
expression of sodium-glucose transporters in cancer. Proc Natl Acad Sci USA 281
112:E4111-4119.
282
12. Shiba K, Tsuchiya K, Komiya C, Miyachi Y, Mori K, et al. (2018) Canagliflozin, 283
an SGLT2 inhibitor, attenuates the development of hepatocellular carcinoma in a 284
mouse model of human NASH. Sci Rep 8:2362 285
13. Saito T, Okada S, Yamada E, Shimoda Y, Osaki A, et al. (2015) Effect of 286
dapagliflozin on colon cancer cell. Endocr J 62:1133-1137.
287
14. Kadokura T, Saito M, Utsuno A, Kazuta K, Yoshida S, et al. (2011) Ipragliflozin 288
(ASP1941), a selective sodium-dependent glucose cotransporter 2 inhibitor, safely 289
stimulates urinaly glucose excretion without inducing hypoglycemia in healthy 290
Japanese Subjects. Diabetol Int 2:172-182.
291
15. Mao W, Zhang J, Komer H, Jiang Y, Ying S. (2019) The emerging role of voltage- 292
gated sodium channels in tumor biology. Front Oncol 9:124 293
294
FIGURE LEGENDS 295
Fig. 1. SGLT2 is expressed in human breast cancer cells 296
(A) Immunohistochemistry was performed to examine GLP-1R expression in breast 297
cancer cell lines. All samples were counterstained with DAPI (magnification, 400×). (B) 298
Quantitative RT-PCR was performed using a set of primers targeting 94bp coding reqion 299
of SGLT2. TBP expression was used for normalization. Unpaired t-tests were performed 300
to calculate statistical significance (
*P < 0.05 vs. KPL-1 cell) (n = 3).
301
Fig. 2. SGLT2 inhibitor attenuated breast cancer cell proliferation
302
(A) MCF-7 cells were maintained in media supplemented with 10% fetal bovine serum 303
(FBS) with saline or ipragliflozin (1–50 M). After 0, 24, 48, 72 and 96 h, the cells were 304
harvested and cell proliferation was analyzed by cell counting using a hemocytometer.
305
Black circles with solid line, control (non-treated; DMSO); black cicle with dotted line, 306
ipragliflozin (1 M); white squares with solid line, ipragliflozin (10 M); white squares 307
with dotted line, ipragliflozin (50 M). Two-way ANOVA were performed to calculate 308
statistical significance (
**P < 0.01 vs. control) (n = 3) (B) MCF-7 cells were transfected 309
with either negative control duplexes or small interfering siRNA targeting SGLT2 and 310
maintained in media with 10% FBS with DMSO or 10 M Ex-4. After 0, 24, 48, 72 and 311
96 h, the cells were harvested, and cell proliferation was analyzed by cell counting using 312
a hemocytometer. Black circles with solid line, siControl and DMSO; black cicle with 313
dotted line, siControl and ipragliflozin (10 M); white squares with solid line, siSGLT2 314
and ipragliflozin (10 M); white squares with dotted line,siSGLT2 and ipragliflozin (10 315
M). Two-way ANOVA were performed to calculate statistical significance (
**P < 0.01 316
vs. control) (n = 3) (C) MCF-7 cells were plated at a density of 5000 cells/well in 96-well 317
plates in media supplemented with 10% FBS, and incubated with ipragliflozin (0–100 318
M) for 24 h. BrdU solution was added during the last 2 h, and cells were harvested for 319
measurement of DNA synthesis using a microplate reader at 450–620 nm. Mean data are
320
expressed as a ratio of control (non-treated) cell proliferation. Unpaired t-tests were 321
performed to calculate statistical significance (n = 3).
322
Fig. 3. SGLT2 inhibitor ipragliflozin induced hyperpolarization of MCF-7 cell 323
membrane 324
(A) Whole cell patch recording for current and voltage clamps were recorded using the 325
nystatin-performed patch technique in MCF-7 cell at room temperature (22-25℃) with 326
an Axopatch 200B (Molecular Devices, Sunnyvale, CA) patch-clamp amplifier. (B) 327
Ramp pulses (-80mV- +60mV, 0.28V/s) were applied every 10 s from a holding potential 328
of +40mV.
329
Fig. 4. GLT2 inhibitor ipragliflozin induced mitochondrial membrane instability 330
(A) Mitochondrial membrane potential was assessed by JC-1 staining in MCF-7 cells 331
treated with DMSO or 10 M ipragliplozin (Ipra) for 48 h. Photos at 0, 24 and 48 h 332
treatment are depicted. (B) Area rate of JC-1 fluoresence was plotted whole time of 333
experiments.
334 335
336
337
338
Figure 1
SGLT2 DAPI Merge
SGLT2(94bp)
TBP(170bp)
MCF-7 MDA-MB231 KPL-1 A
B
Relative expression of SGLT2 mRNA
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2
MCF-7 MDA-MB231 MDAMB231 KPL-1
*
100bp
100bp 200bp
Figure 2 A
B
0hr 24hr 48hr 72hr 96hr
0 50 100 150 200
treatment time(hr) si control(DMSO)
si control(Ipragliflozin10uM) si SGLT2(DMSO)
si SGLT2(Ipragliflozin10uM)
Number of viable cells (×104)
0hr 24hr 48hr 72hr 96hr
0 20 40 60 80 100 120
treatment time(hr) control(DMSO)
Ipragliflozin(1uM) Ipragliflozin(10uM) Ipragliflozin(50uM)
Number of viable cells (×104)
**
**
**
**
**
**
**
**
****
**
****
**
C
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8
0 1 10 50 100 (mM) Ipragliflozin concentration
Relative incorporations
* *
Figure 3 A
B
*
*
* Glucose(+)
Control (normal medium)
*
*
*
*
C D
Figure 4
A
B
Area rate of JC-1 fluoresence (DMSO)
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 0
20 40 60 80
time-lapse (hr)
JC-1 fluoresence Area (%)
J-aggregates J-monomers
Area rate of JC-1 fluoresence (Ipra)
time-lapse (hr)
JC-1 fluoresence Area (%)
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 0
20 40 60
80 J-aggregates
J-monomers Control
DMSO
Ipra 10μM