海 馬
シナ プ
ス機能
を 下
位
領 域
間
で
比
較
す
る
一
非 侵
襲
的 測 定 研 究
の.
可能 性
一
岡
田
隆
*上 智 大 学*
Comparison
of
synaptic
function
in
hippocampal
subfields
一
Prospects
for
noninvasive
measurement
of
human
hipPocampal
activity
一
Takashi
OKADA
*SoPhia
University
*
In
variQus neuroscientific studies thehipPocampus
is
reported tQ play crucial rolesin
dec
}.
ara−
tive memory
.
The hippocampus is composed of several synaptic fie!d subdivisjons,
such as thedentate
gyru.
s,
CA3
andCA1 .
Synaptic
plasticity in these areas is thought to be a candidate for thebio!ogicaL basis of memory function
,
In
the 上astdecade
therehave
been
efforts to elucidate the memoryfunctions
of each synaptic subfield by using gene−
manipulated animals and recombinant viral vcctor−infected
animals.
The
underlying circu圭t
mechanismshave
been
studiedby
electro−
physiologicai recordings
from
thehippocampus
of the animals.
High
spatial resolutionfunctional
MRI
mustbe
used to clear!y define the activated areas within the hippocampusof the living humanbrain,
and studies using ahigh
resolutionirnaging
systemfor
that
purposehave
recently begun toappear
.
The prospects and challengesfor
future
integration
Qf animal andhuman
studies arediscussed,
Key
words :hippocampus ,
dentate
gyrus,
CA3 ,
CAI,
synaptic plasticity,
functlonal MRI記 憶の生 理 心 理 学 的 研 究の課 題は
,
記 憶 機 能を担 う脳 部 位 (「記 憶 の座」)の同定と,
記憶に対 応する脳 内 変 化 (「記 憶 痕 跡 」)の同 定で あ る とい え る。 この双方に お け る 研 究が最 も進んで い る脳 部 位は海 馬で あ る。
海 馬 摘 出に よ る健忘 症 患者の 事例(Scoville
& Milner,
1957)が古 く か ら知 ら れてお り,
ま た,
シ ナプス前 線 維に対す る高 頻 度 刺 激に よ り海 馬シ ナ プス 応 答が増 加しそ れ が長 期 的 に維持さ れ る現象 (長 期増 強)(Bliss
&L
¢mo,
1973
)を は じめ と する シ ナプス 可塑性は,
記 憶に重要な役割を 果 た す海 馬に お ける伝達効 率の長 期 的 変 化で あ ること か ら,
記 憶 痕 跡にか か わ る もの で ある との期 待の も とに数 多くの研 究がな されて きた。* Department of PsychQlogy
,
Faculty of HumanSciences
,
Sophia University,
7−
1 Kioi・
cho,
Chiyoda−
ku,
Tokyo IO2−
8554E
−
mai1 :0kadat @ sophia.
ac,
jp
動 物の脳を用い た長 期 増 強の典 型 的な実験は
,
まず 海 馬シナプ ス前 線 維に配 置した刺 激 電 極を通 じ て比 較 的 弱 い電 気刺 激を低頻度で (た と え ば10
秒に 1度の 頻度で) 与え.
各 刺 激 時の シナプス後 細 胞の応 答を,
シナ プス 部 に配 置 した記録電極を用い て検 出しモ ニ ター
する こと か ら始ま る。 毎回の シナ プス応答の大き さ が ほ ぼ安 定し た こと を確 認 し た後,
長 期 増 強 誘 導の た めの刺 激を与え る。 多 くの場 合に は高頻度 電気刺激が その誘導の た め に 川い ら れ,
高頻度の 電気刺激を 短時 間 (た と え ば 100Hz の頻度の刺激 を 1秒 間,
そ れ を 1〜
3回 ) 与え る とい う もの である。 そ の後,
ま たシ ナプス応 答の モニ ター
を続 け,
高 頻 度 刺 激 前の シ ナ プ ス応 答と高 頻 度 刺 激 後の シナ プ ス応答との問で大きさ の比 較を行う。
海 馬ス ラ イ ス標 本 を用いた実 験の場 合,
高 頻 度 刺 激か ら1
時 間 後におい て もシナプ ス応 答が増 大し た ま まであ れ ば,
長 期 増 強の 誘 導が な さ れ た とみな さ れ ることが一
般 的である (よ り18
基礎心 理学研 究 第 28 巻 第1
号 詳しくは,
岡田(2005 )参 照 )。
海馬シナ プス応答の長 期増 強が記憶 機能の 生物学 的基 礎といえる の か どうか を知る 目的で,
長 期 増 強 誘 導に対 する阻害 操作ま た は促 進操作後の学 習 成績につ い て検討 し た動 物 実 験が報 告さ れて い る。 長 期 増 強と課 題 成 績と の間に正の相 関 関 係の あ る こ とが示さ れ(Morris,
An −
derson,
Lynch,
&Baudry ,
1986;Tsien,
Huerta,
&Tonegawa
,
1996;Tang,
Shimizu , Dube , Ram pon,
Ker−
chner,
Zhuo ,
Liu,
&Tsien,1999
),
い っ た ん発現 し た長期 増 強を約 1H 後に元の 水 準の シ ナ プス応 答に戻 す 薬 理 的 操 作によっ て その動 物に逆 向 健 忘が生じ ること も明
ら かに なっ た (
Pastalkova,
Serrano,
Pinkhasova,
Wa1 −
lace,
Fenton
,&Sacktor
,2006
)。
海 馬の長 期 増 強 が 記 憶と関 連が あ る とい う考えにつ いて は
,
そ れに合致す る知 見 が,
神 経 科 学 的 手 法による諸 研究 か ら得 られて い る と いえ る。
し か し 近年,
海 馬シナ プス可 塑 性の意 義につ い て,
海 馬 全 体として で はなくその下 位 区 分ご とに理 解 しよ う と する研 究が行わ れ始め て い る。
海馬は神経 細胞同士の結 合か らな る神 経 回 路 網であ りt 海 馬 外の神 経 細 胞との結 合,
およ び 海 馬 内 部にお ける神 経 細 胞 同 士の結 合 が あ る。 つ ま り,
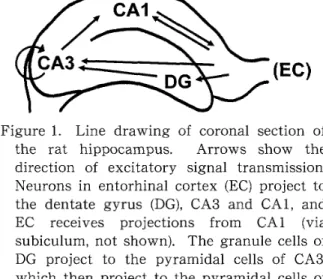
海 馬に は複数の下 位 領 域が存 在す る。 これ ら下 位 領 域の う ち主 要 な もの に,
歯 状 回,CA3 ,
CA1
が 挙げ ら れ る。 興 奮 性シナ プス伝達 を 生 じ さ せ る経 路と し て,
嗅 内 皮 質→
歯 状 回→
CA3→CA1 →
海 馬 台→
嗅 内 皮 質 とい う一
方向性の 経路と,
嗅 内 皮 質か らCAI
やCA3
へ の直 接 的 入 力 経 路,CA3
錐 体 細 胞か らCA3
錐 体 細 胞へ の連 合 性 結 合が知 られて い る (Figure1
)。 部 分 的に は戻 りの 経 路も あ る が,
全 般 的には 海馬は一一
方 向 性の信 号 伝 達を行 っ て おり,
これ は双方向 性の信 号 伝 達を行う大 脳 皮 質と は著し く異なる特 徴で あ る。
な お,
石 塚 (2002 )は一
連の 解剖 学的研 究に よ り, 海馬の 下 位領域間の結 合を 別々 の皮 質 領 域 間の結 合と して と ら え るの で は なく,
同一
皮 質 領域 内の層間 結合と して解 析すべ きで あ る と提 唱 して い る。
シ ナプス応 答の長 期 増 強は.
お もに齧歯類を用い た実 験に よ り,
どの海 馬 下 位 領 域で も生 じる こ と が示さ れて い る。 すなわち,
歯 状 同 領 域の シ ナプス,CA3
領 域の シ ナ プス (歯 状 回 顆 粒 細 胞か ら の苔 状 線 維との シ ナ プ ス と CA3 錐 体 細 胞 同 士の シナプ ス),
CA1 領 域の シ ナ プ スの いずれの下 位 領 域に おいても,
シ ナ プス前 線 維へ の 高 頻 度 刺激後 に長 期 増 強が 生 じ る(Bliss,
Co]lingridge,
&Morris,2007
)。 シナ プス可 塑 性の機 能的 意義にっ い て下 位 領 域 間で違い はあ る のか,
また違いが ある と す れ ばそ れ を 生ん で い る生 理 的 基 盤 は何か,
とい う問題 が 近年の{
EC
)
Figure
1.
Lirle drawing of coronal secti 〔)n ofthe rat hippoca皿 pus
.
Arrows show the
direction
of excitatory signal trarlsmissiQn.
Neurons
in
entorhinal cortex (EC
)project tothe dentate
gyrus
(DG ),
CA3
andCA1 ,
andEC receives projections from CA 1 〔via
subiculum
,
not shown }.
Thc granule cells of
DG
pr〔〕ject
to the pyramidal ce]ls
ofCA3 ,
which then
project
to thepyramidal
cells ofCA1
.
Strong associational connections exist withinthe
CA3
area,
課 題と なっ て い る。 こ の テ
ー
マ に関 して は,
現 時 点で は,
さ まざま な実 験 操作が 可能な勤物実 験によ る知見に限ら れてお り,
な か で も齧 歯 類 を 用い た研 究がほ と んど を 占め る。 本 稿で は,
齧 歯 類 動 物 を対 象として これ まで用 い られて き た研 究法につ い ての概 略を 述べ た あ と,
シナ プス 可塑性の機 能 的 意 義 を 下 位 領 域 別に調べ る研 究 が,
人 間 を対象と し た非 侵 襲 的 測 定 法によっ て行わ れる可 能 性にっ い て考え る。
動物実
験に よ る アプロー
チ例1980
年 代 後 半 に遺 伝 子 欠 損 動 物の作 製 法が確 立 さ れ,
あ る特 定の タン パ ク質を欠い たマ ウス の 行 動 変 化を 調べ るこ とにより, その タン パ ク質の機能を明らか に す る とい うア プ ロー
チ が と ら れ始めた。
1990 年 代に は,
遺 伝 子発 現の ため のプ ロ モー
ター
を適 切に 選ぶ こ とで遺 伝 子 発 現に部 位 特 異 性を持た せ,
あ る特 定の脳 部 位の細 胞に お いて の みタン パ ク質の操作 を行うと い っ た こと が 可能と なっ た。
こ の方 法を 用 いる と,
長 期 増 強に 必要な タンパ ク質を,
あ る海 馬シ ナ プス領 域 特 異 的に欠 損させ ることに より,
その シナ プス領域だ け長 期増 強を阻害す ること がで き る (通常の シ ナ プス 応 答その もの は阻害さ れな い点が実 験上 重要で ある)。 さ ま ざ ま な学習 課 題に おけ る そ の遺 伝 子 改 変 動 物の成 績を調べ る ことに よ り,
各シ ナ プ ス領 域の可 塑 性が記 憶 機 能におい て果たす 役 割 を推 測し ようと い うア プ ロー
チ である。 海馬の 興奮 性伝述 を担う主要な神 経 伝 達 物 質はグル タ ミン酸で あ る。 グル タ ミン酸受容体のサ ブ タ イ プの一
っ NMDA 受容体 を 遺 伝 子 工 学 的 手 法に よ り 欠 損 させ ると
,
NMDA 受 容 体を介 し たCa2
+ 流 入 が長 期 増 強の誘 導 刺 激 とな る領 域 (歯 状 回シナプス,
CA3
連 合 性シナプ ス,CAI
シナプス)での 長期 増強が阻 害さ れ る。 適 切な プ ロ モー
ター
を用い ることに より,
シナプス領 域の いず れ か一
つ で こ のNMDA
受 容 体 欠 損 を 生 じ さ せ れ ば,
領 域 特 異 的に長 期 増 強を阻 害す ることに な る。 こ の被 験 体 が成績 低下 を 示 す 記憶課 題の性質から,
シナ プス可 塑 性 の機 能が下 位領域 別に示唆 さ れ る。NMDA
受 容 体 欠 損 につ いて は,
これ までCAI
錐 体 細 胞の み 〔Tsien
et a1.
,1996 )
,CA3
錐体 細 胞のみ (Nakazawa ,
Quirk
,
Chit・
wood,
Watanabe,
Yeckel,
Sun,
Kato,
Carr,
Johnston,
Wilson,
& Tonegawa,2002
),
歯 状 回顆 粒細 胞のみ (McHugh,
Jones,
Quinn,
Balthasar.
Coppari,
Elmqui9.
t,
LowelL,
Fanselow ,
Wilson,
&Tonegawa ,2007
)でNMDA
受容 体を欠 損さ せ た場合の,
マ ウス の学 習成績 低 下の性 質 を 調べ た研 究が報 告 さ れて い る。ま た
,
後天 的に遺伝子 導入 を 行 う方 法 も ある。Okada ,
Yamada
,
Tsuzuki,
Horikawa,
Tanaka,
&Ozawa
(
2003
)は,
組 換えウィ ル ス ベ クター
を海 馬 下 位 領 域に局 所 的に感染 さ せ,
あ るド位領域に限定 して長 期増強の強 さ を 変 化 させ た際の学 習 成 續 を 比 較 し た。Ca21
透 過 型AMPA
受容 体の サ ブユ ニ ッ1・
を 過剰発 現 さ せ,
局所 的 にCa2
+ 流 入 量 を増 加さ せ るこ とによっ
て長 期 増強誘導 閾 値 を 下 げた。
その結 果,
CAI
へ の こ の操 作は水 迷 路 学 習 成 績を 上昇さ せ た が,
歯 状 回へ の操作は逆に成 績を低 ドさせる こと が示された。 ウ ィル ス ベ ク ター
の 種 類に よっ て は細胞 死を誘 導す る 可能 性が あ る ため短期で終わ る学 習 課 題の選 定 等の工 夫が必 要にな る場 合 も ある が,
ラ ッ トを被 験 体と し て 用い ること が で き る点,
時 期 特異 性・
部 位 特異性を持たせ るこ と が容易で あ る点で, 下位 領 域 別の解 析をするうえで の有 望な方法と言え る。 な お,
長 期 増強と学 習課 題 成 績との因 果関 係を明 確に 示 し た動物 実験 は ほ と ん ど ない が, 近 年 報 告 さ れ たWhitlock
,
Heynen,
Shuler,
& Bear (2006 }の研 究で は,
即 座に習得が可能な受動 的回避学習を用い, 複数の電極 を用い た CA1 か らの記 録を行うこ とに よ っ て
,
学 習 誘 導 性の長 期 増 強の生 起が示されて い る。 こ の実 験に お い て は,
ラ ッ トCA1
か ら同 時記録 し た別の電 極か ら は学 習 後に長 期 増 強で は な く 長期 抑 圧 (シナプス 応答 が 長期 的に減 じ ら れ た ま まにな る現 象 )が記録 さ れ た もの も あっ た。
つ ま り,
伝 達 効 率 変 化の方 向性が 領 域 内におい て 異 なっ てお り,
海 馬 ド位 領域を さ らに細か く 解析す る 必 要 性を示 唆して い る。 以上の よ う に,
あ る特 定の シ ナプ ス領 域の長 期 増 強を 操 作し た際の効 巣は,
操 作さ れ る領 域に よ っ て異な る こ と が示さ れて い る。 し か しそ の一
方で,
歯 状 回・
CA3・
CA
1い ず れの領 域の シナプ ス も,
入 力 線 維へ の高 頻 度 刺激に よっ て長 期増強が誘導さ れ る とい う共 通性があ る。 どの よ うな神 経 回 路レベ ル の特 徴の違いが シナ プス 可 塑 性の機 能差 を 生 む 可 能 性が あ るの か,
とい う生 理 心 理学 的 研 究が今 後 必 要で あ り,
そ の領 域 間比較を 適切に 行 う に は,
同一
の実 験 者 が 同一
の実 験 系 を 用いて全 下 位 領 域の解 柝を行うこと が 望 ま しいが,
その よ う な 研 究 は 希で ある。 その少 ない例 と して,
シナ プス伝 達に対 する 調 節物 質の作用が領域ごとに異な るこ とを,
同一
の実験 条 件 下で示 し た研 究が あ る。 Suzuki & Okada (2007,
2009 )はカ リウムチャ ネル 阻 害に よっ て誘 導さ れ る長 期 増 強に対 し,GABA
(γ・
ア ミノ酪 酸 )入 力の効 果が,
歯 状 回 (貫通 線 維一
歯 状 回 顆 粒 細 胞間シナプ ス) とCA3
領 域 (苔 状線 維一
CA3
錐 体 細 胞 間 シ ナ プス) とで は 正 反 対 と な ること を,
ラッ ト海 馬ス ライス標 本 を 用い た電 気 生 理 学 的測定 に よ り明 ら かにした。 こ れ は,
海 馬 全 体のGABA
濃 度の高 低に よっ
て,
長 期 増 強が生じ や す い シ ナ プ ス領 域 が 変 化 する こ とを 示 唆 した研 究で ある。 以 上の よ う な,
動 物お よ び その 脳標 本を対 象と し た実 験 手 法により,
海 馬 下 位 領 域の可 塑 性の持っ機 能 的 意 義 の違いが 調べ ら れ て き た。
動 物 実 験は遺 伝 子工学 的な操 作や電 気 生理学 的 測定が可能であ る点で 強力なアブm一
チ法で あり,
人 間 を 対 象とした研 究 だ けで は知 りえない 脳機能を私た ちに教えて くれ る。 し か し な が ら,
心 理学 の 目標が人 間の心の解 明で ある な らば,
これ らの動 物 実 験によ る知見 や そ れに基づ くモ デル が, 人間 に どの 程 度.
一
般 化 可 能なのか とい う視 点 を 忘 れるべ きで は ない。 動 物 実験 か ら 示唆さ れ た 脳機 能が 人 間 に も当て は ま るの か どうか を, 人間を対象と し た非 侵襲 的脳機能測定 法によ り,
部 分 的に で も検 証 すること は可 能で あ ろうか。
非
侵襲
的脳機能
測 定 法に よ るア ブ囗一
チの可 能 性 人 間の海 馬は側 頭 葉 内 側 部に位 置して い る ため,
海 馬 の非侵襲 的機 能測定を試み る場合,
た と え ば大脳皮質の 活 動を記 録 する の に適 した EEG やNIRS 等の方 法を 用 い るこ と は で きず, fMRI を 用い るこ と に な る。 fMRI は 神 経 細 胞の 活 動が引き金と なっ て生じ る血 流 変 化に基づ く信 号で あ り,
その 意 味では間 接 的な測度で あ る が,
そ の際の血管 収 縮や拡張を司るの は グ リア細 胞 (ア X トロ サ イト)の酸 素 依 存 性 代 謝 活 動で あ るこ と が 近年 明ら か にな っ て お り(Gordon,
Mulligan,
& MacNicar,
2007 ;Gordon ,
Choi,
Rungta.
Ellis−
DaVies,
& MacVicar ,2008
),
脳 機 能 測 定の根 拠 と なるカ ス ケー
ドも明 確に なっ て き た。
これ ま で,
人間を研 究対 象と し た非 侵 襲 的20
基 礎 心理学 研 究 第 28 巻 第 1号測定に おい て 海馬 の活 性 を 調べ た 研 究 は あ る が (
Eldri−
dge,
Knowlton,
Furmanski,
Bookheimer ,
& Engel,
2000 ;
Gais,
Albouy ,
Bo!y,
Dang −Vu ,
Darsaud ,
Des −
seilles,
Rauchs,
Schabus ,
Sterpenich,
Vandewa !lc,
Maquet
,
& Peigneux,
2007 ;ShDhamy & Wagner,
2008
),
海 馬 下位 領 域ご との活性に言及 し た もの はほ と ん ど ない。 その理 由と して,
海 馬 下 位 領 域 別の解 析の必 要 性が 現時 点で は ま だ広く 認 識 さ れ てい ない とい う 可能 性,
お よ び機 能 画 像 法にお け る空 間 解 像 度の問 題が挙げ られよう。
しか し,
3 テ ス ラ の 磁 場 を 用い て,
人 間の偶 発学 習 時の 海 馬 活 性を下 位 領 域 別に解 析し た研 究が近 年 発 表さ れ た (Bakker,
Kirwan,
Miller,
& Stark,
2008 )。
歯 状回 (
dentate
gyrus,
DG
)とCA3
に関 して は,
そ れ ら の活 性 化を空 間 的に分 離で きた とは主 張せず,
その領 域 の活 性 化 部 位を“
DG
/CA3
”
と 記載す る に と ど めて い る が,
こ の 研 究で は,CAI
や海 馬 台な どの ド位 領 域の 活 性 化 を 区 別 して示 して い る。
より 高 磁 場 を 用い た機 能 画 像 法が可能と な れ ば歯状 同とCA3
と を区 別して 活性 化を 測 定で き る よ うにな る と思わ れ る し,
動 物 実 験にお ける 海 馬 下 位 領 域ごと の解 析が さ らに進みモデル が確 立さ れ れ ば, その ような解 析に着 目する認 知神経 科 学 者の増 加 も期 待さ れ る。
な お,
形態研究に おいて は,
高 磁 場を 用 い たMRI
に より, 海馬の下 位領域の 形 態を従 来よりも 明 確に示 し た例がある。
4 テス ラMRI を 用いて,
海 馬の 各下位領域の加 齢変化を解析し た例(Mueller,
Stables,
Du
,
Schuff,
Truran,
Cashdollar,
& Weiner,
2007 )や,
7テ ス ラ MRI に よ りさ らに明 瞭な画 像 化に成 功 し た例 (
Thomas ,
Welch ,
Niederhauser,
Whetsell,
Anderson ,
Gore
,
Avison,
& Creasy,
20Q8 ; Theysohn,
Kraff,
Maderwald
,
Schlamann,
de Greiff,
Forsting,
Ladd,
Ladd ,
&Gizewski,2009
)が近 年 示さ れて い る。
空 間 解像度の問 題と は別に,
シナプ ス可 塑 性に関 する 海 馬 下 位 領 域 別の 機能の研究 を入 聞で試み る場 合.
海 馬 シ ナ プ ス伝 達の程 度を断 続 的に記 録 する た めの手 法の確 立が必 要 とな るの も困 難さの一
っ である。 動 物や動 物 脳 標 本 を 用い た実 験に おい て は,
シ ナ プス伝 達の 大 きさ は,
上 述の よ うに短い電気刺 激 (0.
1 ミ リ秒な ど)を 入力 線 維に…
定の低頻 度 (10
秒お き な ど)で与え,
その と き の シ ナプス 応 答 を毎回記録するこ とによっ て シナ プス伝 達効 率の 変化 を 知 るe 入間を対象 と し た 研 究 の場合,
可 能 性が あ る と す れ ば,
経 頭蓋磁 気 刺 激 法によ る海 馬へ の 求 心 路 刺 激と機 能 画 像と を組み合わ せた実 験で ある と考 え ら れ る。 ま た,
空間的 記 憶を顕 著に利用する タ ク シー
運 転 手に おい て,
海馬の一
部の サ イズが経 験 年数に従って増大 して い る とい う 研 究 (
Maguire ,
Gadian,
John一
srude
,
Good ,
Ashburner,
Frackowiak
& Frith,
2000 )から示 唆され るよ うに
,
形 態 的な可 塑 的 変 化が,
繰 り返 しの学習経 験によっ て生 じ る ならば,
学 習が引き起こす 脳 構 造の変 化とい う意 味で の 可塑 性が海 馬 下位 領 域 問で 異な る か否か を検 討 する研 究 も可 能かもし れ ない。
こ の 目的のため に有 用と 思 わ れ る手法 と して,
客 観的・
自動 的な体 積 評 価 を口∫能にするボ クセ ル ベー
ス の形態 解 析 (voxel−based
morphometry ;VBM
),
神 経線維の方 向を 画 像 化 する ための 拡 散テ ン ソル に よる評 価 (成 瀬・
田 中,2005
)が挙 げられるe 今 後の展望 海 馬の切片の搆造 はラ ッ トも人 間もほぼ同 様であ り,
両 動 物 種の脳の大 き さの違い に応 じたサ イ ズ差 しかない よ う に 見 え る た め,
動 物 種 を 越 えて海 馬 機 能を一
般 化す ることは比 較 的 容 易だ とい う考え方 もある。 しか し,
よ り細 かい神 経 解 剖 学 的 所 見に関 して は,
たとえば 人問の 海 馬CAI
錐 体 細 胞 層はラ ッ トより も相 対 的に厚 く,
ま た左 右の海 馬 を 結ぶ交 連 性の結 合は人 問の ほ うがラ ッ ト よ り も は る かに少ない とい っ た違い も 指摘さ れて い る {Amaral & Lavenex,
2007 )。 人 間の海 馬の神 経 解 剖 学 的 解 析が さ ら に進めば,
両種 間の違い が よ り 多 く指 摘 さ れ る吋能 性 もあ る。 シナ プス可 塑 性に関 する研 究は実 験 手 法 上の制 約 か ら 今 後 も動 物 実 験 を 中 心に進 む と考え ら れ る が,
動物や動 物 脳ス ライス を用いた解 板に基づ いて 立て ら れ た各シナ プ ス領 域の情 報 処輝 にっ いて の仮 説 を,
非 侵 襲 的 脳 機 能 測 定 法に よる人 闇 対 象の研究におい て部 分 的に で も確 認 することに は大き な意 味が あ る。 た とえ ば,
類 似の刺 激を弁 別 すべ き状況下で,
NMDA 受 容体を歯状回特異的に欠損させ たマ ウ ス の場 合にパ ター
ン分 離 (pattern separation )機肯旨が損な わ れ た とい う研 究 (McHugh et aL,
2007)の後,
人間を対象 と した非 侵 襲 的 脳 機能測 定 法によ る研 究か ら も そ れに合 致す る結 果 が得 ら れ た (Bakker et al.
,
2008 )とい うの は,
両アプ ロー
チ の連 携が見 ら れた好 例と言えよ う。 な お本 稿で は fMRI の可 能 性につ い て 論 じたが,
動 物 実 験で 詳 細に判 明 して い る受 容 体 レ ベル で の シナプス可塑 性メ カニ ズ ム (た と え ばCAI
領 域に お け るAMPA
受 容 体の シ ナ プ ス 後 膜へ の トラフ ィ ッ キ ングな ど)に関 する知 見 を 用い,
た と え ば あ る特定の 学 習の 成 績 と グル タミン酸受容体密 度との相 関を調べ る分 子イメー
ジ ン グ をPET
によっ て 行う とい っ た方 法も,
動 物 実 験と人 間の実 験と を結びつ け る口∫能 性を含ん で い る。
ま た,
動 物か ら人間へ の一
般 化 だけ で な く,
人 間 独 自 の 認 知 行 動 と海馬 との関 連 を 知 る う えで も,
認 知神 経 科学 的 研 究は期 待さ れ る。 特に海 馬は 官言的 記 憶にお け る 役 割 が 唱えら れて おり
,
他の 脳 部 位に比べ,
言 語 が 関 与 す る 課題を 用い た 研究の意義が よ り 大 きい と考え ら れ る。 比較心 理学 的 観 点か らも,
非 侵 襲 的 脳 機 能 測 走 法に よ る知 見の蓄 積が望ま れ る。
勤 物 実 験の知 見 を人 間の非 侵襲 的機 能 測定で確認 する とい う方 向に と ど ま らず,
人 聞を対 象に し た実 験で提 唱 された仮 説を動 物 実 験に戻し て詳細に調べ る と い う.
方 向の提携も可能に なるこ とで あ ろ う。 引 用文
献Amara
!,
D.
,
&Lavenex ,
P .
(2007 ).
Hippocampal
Neu−
roanatomy
.
In P.
Andersen , R.
Morris, D,
Amaral,
T.
Bliss
,
&J.
OKeefe (Eds.
),
The Uippocampal Book.
Oxford
:Oxford
University
Press.
pp.
37−
l l4.
Bakker ,
A .
,
Kirwan ,
C.
B.
,
Mtller,
M .
,
&Stark,
C .
E,
L .
(
2008
),
.
Pattern
separationin
thehurnan
hippocam −
pal CA3 and derltate gyrus
.
Science,
319,
1640−
1642
.
Bliss
,
T.
,
Collingridge,
G .
,
&Morris,
R.
(2007 ).
Synap ・
tic plasticity
irl
thehippoca
皿pus,
In
P.
Andersen,
R
.
Morris,
D .
Amaral,
T.
Bliss,
&J.0
’
Keefe (Eds、
),
The ffipPocamPal β oo 舟
.
Oxford ;Oxford University
Press.
pp,
343−474 .
Bliss
,
T .
V .
,
&Lemo ,
T ,
(1973
),
Long −lasting
potentia−
tion of synaptic transmission
in
thedentate
area ofthe anaesthetized rabbit following stimulation of
the perforant path
.
ノbu77iatOf
Physiotogy (1}ondon ),
232 ,331−356.
E工dridge
,
L.
L.
,
Knowlton,
B.
J
.
Furmanski,
C .
S.
,
Bookheimer ,
S.
Y .
,
&Enge1,
S.
A .
(2000
).
Remem −
bering
episodes :Aselective
rolefor
thehippocarn−
pus
dur
工ng retrieval.
勲 敵 Neuroscience,
3,
1149−
1152.
Gais,
S.
,
Albouy ,
G,
,
Boly,
M 、
,
Dang −Vu ,
T ,
T.
,
Darsaud ,
A .
,
Desseilles,
M .
,
Rauchs,
G.
,
Schabus ,
M,
Sterpenich,
V.
,
Vandewalle,
G .
,
Maquet,
P.
,
& Peig−
neux
.
P.
(20Q7),
Sleep transforms the cerebral traceof declarative memories
.
Proceedings of theNa −
tionag
Acadenzy
〔’f
Sciences
of theUnited
States
of胤η2¢%oα
,
104,
18778−18783.
Gordon ,
G .
R.
,
Mul .
ligan,
S、
J
,
,
&MacVicar ,
B .
A .
(2007 )
.
Astrocyte control of the cerebrovascula−
ture
.
Gtia,55,12
】4−
1221.
Gordon ,
G ,
R ,
,
ChoL
H .
B .
,
Rungta ,
R .
L.
,
EUis−Davies,
G .
C .
R.
,
&MacVicar,
B.
A,
〔2008 ).
Brain
metaboli.
smdictates the polarity of astrocyte control over arte
−
rioles
.
Nature,456 ,
745−
749.
石 塚典生 (
2002
).
大脳.
辺縁 系の細胞構築と神 経 結 合 松本 元
・
小野 武 年 (編 ) 情と 意 の 脳 科 学 培 風 館 pp.
26−−44.
Maguire ,
E.
A ,
,
Gadian,
D ,
G .
Johnsrude
,
1、
S.
,
Good ,
C .
D ,
,Ashburner ,
J
,
,
Frackowiak ,
R .
S.
,
&Frith,
C.
D 、
(
2000
).
Navigation−
related structural changein
thehipPocampi of taxi drivers
.
Proec・edings of the ATa−
tional
Academ
ツ ofScienc2S
of thθbTniteal
States
〔ゾ
Ame
γi
’
ca
,
97,4398−4403.
McHugh
.
T.
J,
,
Jones,
M,
W.
,
Quinn ,
J。
J.
,
Balthasar,
N.
,
Coppari
,
R.
,
Elmquist,
∫.
K.
,
Lowel1,
B.
B.
,
Fanselow,
M .
S.
,
Wi
!son,
M .
A .
,
&Tonegawa ,
S.
(2007
}.
Dentate
gyrus
NMDA
receptors mediate rapid patternseparation in the hipPocampal network
,
Science,
317,94・
凵
一
99.
Morris,
R .
G .
M ,
Anderson,
E.
,
Lynch,
G .
S.
,
&Baudry ,
M
.
〔1986).
Selectiveimpairment
of learning and
blockade
of !ong−
term potentiationby
anN −
methyl
−
D−
aspartate receptor antagonist,
AP5.
.
Vatzare,
319,
774−
776.
Mueller,
S.
G .
,
Stables,
L.
,
Du ,
A .
T .
,
Schui
『。
N .
,
Truran ,
D
.
,
Cashdollar,
N.
,
&Weiner,
M.
W .
(2007 ).
Measure−
ment of hippocampal subfields and age
−
relatedchanges with high resolution
MRI
at4T .
Neurobiol一
α91y of
Aging ,
28,719−726 .
Nakazawa
,
K,
Quirk,
M、
C.
,
Chitwood,
R.
A.
,
Wata−
nabe
,
M .
,
YeckeL
M,
E ,
Sun ,
L .
D .
,
Kato,
A .
,
Carr,
C .
A .
,
Johnston
,
D .
,
Wilson,
M .
A .
,
&Tonegawa ,
S.
(2002 )
.
Requirement fQr hippocampalCA3
NMDAreceptors in associative memory recall
.
Science,
297,211−218.
成 瀬 昭二・
田 巾 忠 蔵 (2005 ).
拡 散 強 調 画 像の可 能 性 最 新 医学, 60,
952−
964.
岡田 隆(2005 }.
脳と記憶 岡田 隆・
廣中 直 行・
宮 森 孝 史 (共 著 ) 生 理 心 理 学 サ イエ ン ス社 pp.
61−
74.
Okada ,
T .
,
Yamada,
N .
,
Tsuzuki,
K.
,
Horikawa ,
H.
P.
,
Tanaka
,
K.
,
&Ozawa ,
S.
(2003 ),
Long−
term poten−
tiation in thc hippocampa !CAl area and dentate
gyrus plays
different
rolesin
sPatiallea
「nin9’
Eu
・
r〔)pean
/bumaal
of.
Veu
γoscience,17,
341−349.
Pastalkova
,
E.
,
Serrano,
P.
, Pj.
nkhasova,
D.
,
Wa 工Lace,
E
.
,
Fenton,
A .
A.
,
&Sacktor,
T .
C.
(2006
).
Storage
ofspatial
information
by
the maintenance mecha−
nism of LTP
.
Science,
313,
1141−
1144.
Scoville
,
W .
B.
,
& Milncr,
B.
(1957
).
Loss of recentmemory after
bilateral
hippocampal
lesions
.
ノbur−
nα
t
ρプ八Jeurology,
Ndeuroszarge
勃y and Ps)・
chiatr /i,
20,
11−
21.
Shohamy
,
D.
,
& Wagner,
A .
D.
(2008
).
Integratingmemories
in
thehuman .
brain
:Hlppocampal −
midbrain encoding of overlapping events
.
.
Neuron,
60,
378−
389,
Suzuki,
E,
,
&Okada ,
T,
(20e7 ).
Regiona 上differences inGABAergic nlodulation for TEA
−
induced synapticplasticity in rat hippocampal CAl
,
CA3 and den−
tate gyrus
.
Neuro∫cience Research,
59,
183−
190.
Suzuki,
E.
,
&Okada .
T .
(2009
>.
TEA −induced
long−
term potentiation at hippocampal rnossy
fiber
−CA3
synapses :Characteristics of its i]iduction and ex
−
Pression
.
Brain
Research,1247,21−27.
22
geeitee#Myeg
28ts
ag
1Il} Kerchner,G.
A.,
Zhuo,M.
Liu,G,
&
Tsien,
J.Z.
(1999>.
Genetic
enhancement of learning andmem-ory
in
mice. IVtiture,401,63-69.
Theysohn,
J.
M.,Kraff,O.,
Maderwald. S.Schlamann,
M.
U.,
de
Greiff,
A.Forsting,
M.,
tadd,
S.
C.
Ladd,
M. E.&
GizewskL
E.R.(2e09).
Thehuman
hippo-campus at 7T-z'nvivo
MRI,
HippocamPus, 19,1-7.Thomas, B.P.,Welch, E.B,Niederhauser, B.D., Whet-sel],
W.
O.
Jr.
Anderson,
A.
W.,
Gore,
J.
C.
Avison,
M.
J.
&
Creasy,
J.
L.
(2008),
High-resolution
7TMRI
of
the
humanhippocarnpus
invivo.fournal
of
Mag-netic
Resonance
imaging,
28,
1266-1272.Tsien,
J.
Z,,Huerta, P.T. & Tonegawa,S.
(1996),
Theessential role of hippocampal
CA1
NMDA
receptor-dependent synaptic plasticityinspatial memory.
Cett,
87,
1327-1338.Whitlock,
J.
R.,I-Ieynen,A,J,
Shuler,
M.G.,
&
Bear,M.
F.(2006).
Learninginduces
long-term potentiationinthe hippocampus. Science,313, 1093-1097.