講 演
平成 20 年日本大学医学会秋季学術大会 (2008 年 10 月 25 日) 同窓会学術奨励賞受賞記念講演要旨平成
20
年日本大学医学会秋季学術大会 同窓会学術奨励賞受賞講演
補体系の活性化と腎糸球体障害
藤 田 宜 是
日本大学医学部内科学系腎臓高血圧内分泌内科学分野緒 言
糸球体腎炎などの腎疾患では腎糸球体の障害機序に免 疫反応が関与するとされています1).馬杉復三らのネフ ロトキシンによる実験的糸球体腎炎2)やF.J. Dixon, C.B.
Wilson
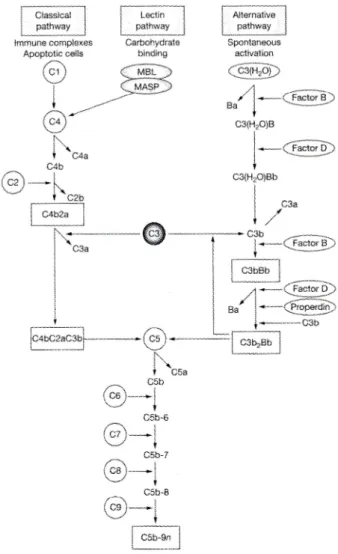
ら3)の実験的免疫複合体腎炎により実験動物での 糸球体障害機序が明らかにされました.ヒトの糸球体腎 炎においても障害された糸球体に抗体や補体が沈着し, 同様の機序が成立していると考えられています. 補体系には現在3
系統の活性化経路が発見されていま す4, 5).第1
は古典経路(classical pathway)
で,抗原に結 合した2
分子の免疫グロブリンに補体のC1q
が結合し, これが次々に補体成分を結合させて組織障害を惹起しま す.この経路は従来から知られている抗原抗体複合体に よって活性化される補体経路です.第2
経路は変更経路(alternative pathway)
とも呼ばれ,抗原や抗体によるini-tiation
が無くてもC3
以降の後期経路の活性化が起こり ます.第2
経路では血中のC3
が加水分解されて流血中 でアクテイブなC3 (H
2O)
を形成するため6),外来抗原の 侵入に際し即座に対応できます.平常時には血中に存在 する制御蛋白によって過剰な反応が抑制されています. 第3
はレクチン経路(lectin pathway)
で外来抗原の糖鎖 部分を構造的に認識して活性化される原始的な経路で す.いづれの経路を介してもC3
以降は同様の後期経路 を通って活性化が進行し,標的組織の障害へと進みま す5, 7)(Fig. 1)
.このような補体活性系の多様化は補体に よる組織障害が抗体を介さず,免疫反応以外でも引き起 こされることを示唆しています.I. 糸球体腎炎における補体系の潜在的活性化
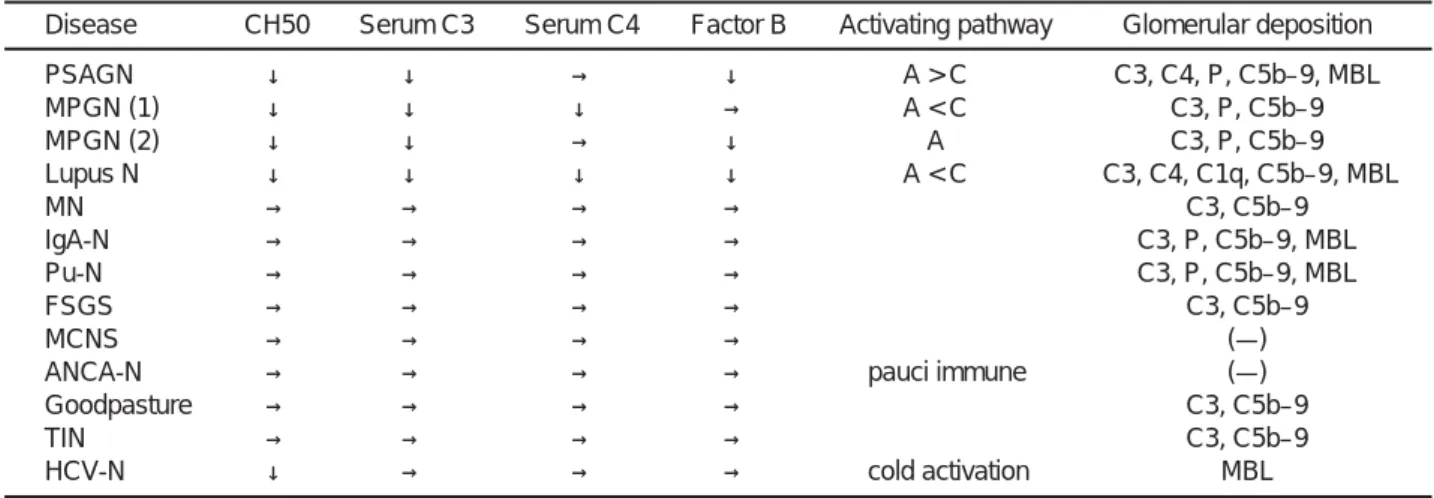
糸球体腎炎では経過中に低補体血症が認められます. これは血中の補体成分が消耗性に低下したためと説明さ れています.私達の検討でも腎炎の活動性に一致した補 体成分の低下を認めています7, 8)(Table 1)
.しかし低補体 血症を示さない腎炎も存在し,これらの病態では補体系 の活性化が起っていないと考えられてきました.補体成 分の血中濃度は補体活性による消費と肝細胞による産生 のバランスの上に成り立っていると考えられます.生体 ではC3
は活性化されてC3b
となった後にH
因子やI
因 子によって分解されてC3c
やC3d
の状態で存在してい るはずです.そこで私達はC3d
の糸球体局在や血中での 残存濃度を検討しました.C3d
の血中濃度は溶連菌感染 後急性糸球体腎炎(PSAGN)
では有意に高値を示し,補 体系の急激な活性化が裏付けられました.また,膜性増 殖性糸球体腎炎(MPGN)
やループス腎炎などの持続性低補体血症を示す症例では
native C3
からC3d
への活性化 が亢進しているもののnative C3
の産生が間に合わず, 慢性の低補体血症がさらなる補体活性を抑制しているこ とが判明しました9).蛍光抗体法による腎組織の検討で はC3c
の沈着を認めない糸球体にC3d
が沈着している ことを報告しました10).また,私たちは糸球体に認めら れるC3
が血中から沈着したものだけでなくメサンギウ ム細胞が産生している可能性もin situ hybridization
によ り明らかにしました11).こうしてC3
が沈着しないだけ で補体系の関与を否定してはいけないと警鐘をならすこ とができました6).II. 血中補体活性化因子による補体系の活性化
PSAGN
では急激で顕著な補体系の活性化が知られて います.外来抗原の暴露により体内で抗体が産生され, 抗原抗体反応が惹起されて免疫複合体型腎障害が起こる ものとされていました.しかし腎糸球体では補体のみが 沈着し抗体が認められない症例についての説明がつきま せんでした.抗体を介さない補体経路を直接活性化する 因子の存在が強く疑われましたが特定されていませんで した.私達は補体活性化物質の同定を試み,溶連菌以外 の抗原により惹起されたAGN
においてその性状を明ら かにしました12).PSAGN
はRodoriguez-Iturbe
らによってneuraminidase
を産生する溶連菌のみで発症することが報告されまし た.一方同じ溶連菌感染でもリウマチ熱では
neuramini-dase
が産生されず,補体系も活性化されません.私達 は溶連菌の産生するneuraminidase
が補体系を活性化す るのではという仮説をたてて検討をおこないました.PSAGN
では低補体血症の推移がneuraminidase
活性や血 中遊離シアル酸濃度と相関し,強い関連性が認められま した.遊離シアル酸の変動は急性相反応蛋白の増加とは 一致せず,in vitro
における補体の活性化実験により補体 第2
経路を活性化することを明らかにしました13).III. 血中補体制御因子と補体活性の持続
MPGN
ではWest
らによって持続性低補体血症が報告 され,その原因として補体活性化中間体C3bBb
に対す る自己抗体であるC3 nephritic factor (C3Nef)
が発見され ました.私達はC3Nef
がC3bBb
に結合しC3bBb
の作 用を持続させる性質を利用して,国内の多くの施設から 届けられた血清を用いて検討をおこないましたがMPGN
の一部にしかC3Nef
は検出されず,その動向も腎組織障 害の程度とは一致していませんでした14).しかし血清C5
値の低下した症例では強く糸球体障害を認めることがわ FSGS → → → → C3, C5b–9 MCNS → → → → (—)ANCA-N → → → → pauci immune (—)
Goodpasture → → → → C3, C5b–9
TIN → → → → C3, C5b–9
HCV-N ↓ → → → cold activation MBL
PSAGN: postatreptcoccal acute glomerulonephritis, MPGN: membranoproliferative glomerulonephritis,
LUPUS N: lupus nephritis, MN: membranous nephropathy, IgA-N: IgA nephropathy, Pu-N: purpura nephropathy, FSGS: focal segmental glomerulosclerosis, MCNS: minimal change nephrotic syndrome, TIN: tublo-interstitial nephritis, ANCA-N: anti-neutrophil cytosomal antibody related nephropathy, HCV-N: heatitis C virus related nephropathy, MBL: mannose binding lectin, A: alternative complement pathway, C: classical complement pathway
Table 2 % inhibition of C3Nef activity by additional
Factor H16)
C3bBb stabilizing activity
Patient EDTA-GVB H I H + I Per cent
inhibition Non-hypo 1 76 56 77 35 55 2 86 56 86 50 43 3 75 57 76 49 35 Hypo 6 68 95 99 86 12 7 96 86 92 79 17
The C3bBb stabilizing activities of three non-hypo patients and two hypo patients were further investigated following addition of factor I. Even when factors H and I were added simultaneously, the C3NeF of the non-hypo patients was inhibited, but that of the hypo patients was not.
Fig. 2 Competitive deposition between C5b-9 (MAC) and Factor H on glomerular epithlium during complement activation18).
かり15),後期経路の活性化を引き起こす引き金は
C3Nef
だけでなく
Factor H
の機能低下が存在することを報告し ました16)(Table 2)
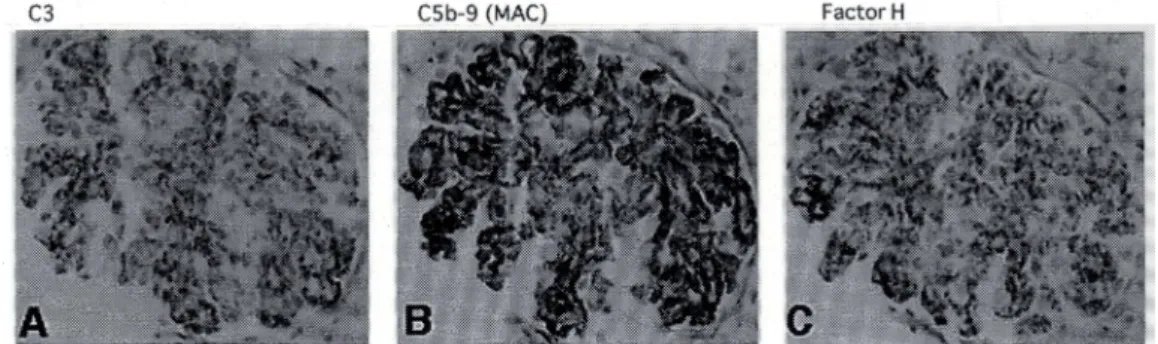
.糸球体においても後期経路の活性化により形成される
membrane attack complex (MAC)
とこ れを制御するFactor H
が競合していることを明らかにし ました17∼19)(Fig. 2)
.AGN
では前述のように低補体血症が一過性で遊離シ アル酸の有意な増加を認めます.一方でMPGN
は低補 体血症が持続し,血中の遊離シアル酸は低値を示しま す.そこで私達は遊離シアル酸が低補体血症の持続性に 関連しているのではないかという仮説をたてて,ループ ス腎炎を対象に遊離シアル酸を測定した.遊離シアル酸 が高値群では低補体血症が改善しましたが,遊離シアル 酸が低値群では低補体血症が持続しました20).さらにin
vitro
において遊離シアル酸の補体活性への影響を検討す ると,ノイラミン酸は補体第2
経路の活性化を抑制する ことが明らかになりました20)(Fig. 3)
.IV. 補体レクチン経路の活性化
レクチン経路は補体経路の中では比較的最近になって 発見された経路で,この経路は抗体を介さないだけでな く第2
経路の構成成分にも関与しないため私達が日頃研 究を続けてきた過程でどうしても説明のつかない現象を 説明することができました.私達はAGN
・IgA
腎症・紫 斑病性腎炎・HCV
関連腎症など種々の腎疾患において レクチン経路を介した腎糸球体障害の存在を報告しまし た21∼25)(Fig. 4)
.V. 代謝異常による補体系の活性化
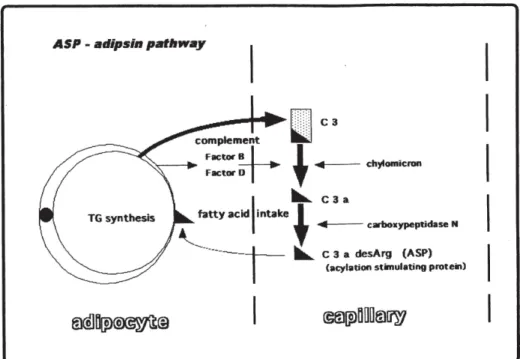
補体成分の多くは肝細胞で生成されます.しかし,脂 肪細胞においても産生されることが知られています.肥 満により活性化された脂肪細胞では補体第2
経路の成分 が多く産生されます.産生されたC3, Factor B, Factor D
などは食後のChylomicron
の存在下で活性化されてASP
(acylation stimulating protein, C3a desArg)
となりますInhibition of C3 breakdown (A) and Factor B breakdown (B) by NANA. The activation of C3 and Factor B was estimated by enzyme immunoassay in incubation mixtures consisting of NHS with various concentra-tions of NANA and 10 mg/mL of LPS as an activator of the alternative complement pathway. C3 or Factor B in NHS was cleaved into the break-down products after incubation witbout NANA, but was significantly linle cleaved in the presence of NANA respectively. These data suggest that NANA might inhibit C3 and Factor B cleavage via complement activation.
(Fig. 5)
.私達はASP
形成の過程でChylomicron
によりC3 tick-over
が亢進し,Factor H
の作用が抑制されるこ とで補体第2
経路がゆっくりと活性化されることを報告 しました26)(Fig. 6)
.この活性化により糖尿病では腎合併 症が誘発され27),肥満関連腎症ではアラキドン酸代謝酵 素を巻き込んだ糸球体硬化を惹起している可能性を報告 しました28)(Fig. 7)
.糖尿病では血管合併症は蛋白質の糖 化や糸球体過剰濾過などが主因とされ,免疫反応は一部 の糖尿病でおこる稀な病態と考えられてきました.私達 は補体活性が免疫反応の枠を超えて引き起こされる点に 着目し,糖尿病モデル動物に補体活性阻止薬を投与して 腎障害の進行を抑制させました29).また,2
型糖尿病患 者で補体活性化産物が増加していることを報告し30),合 併症の増悪因子として補体系の活性化にも注意が必要で あると喚起を促す事ができました(Fig. 8)
.結 語
私達は腎傷害の発症機序の解明と進展阻止という遠大 な計画のもとに色々な検討を行って参りました.特に補 体系の活性化による糸球体障害という点に着目し一定の Fig. 4 Complement activation through the lectin pathway on glomerulus23).Fig. 5 Complement components are produced from adipocyte and activated by chylomicron via the ASP-adipsin pathway7).
Fig. 6 Chylomicron-rich serum accelerates C3 tick-over by regu-lating the role of Factor H26).
Fig. 7 ASP and MAC were significantly increased in diabetic patients but were not elevated in patients with minimal change nephrotic syndrome27).
Fig. 8 Complement inhibitor (K-76 COONa) regulates glomerular injury in diabetic rats (OLETF)30).
成果をあげることが出来ました.ひとえに諸先輩方の御 助力のおかげだと感謝しています.此の場を借りて御礼 申し上げます.補体活性は上述のように外来抗原の暴露 によるもの,自己抗体の産生や免疫複合体によるもの, 代謝異常による非特異的な活性化など多彩で,これから も目標到達への道程は遠いかも知れません.此の度の受 賞を励みにさらなる努力と精進に勤めたいと存じます. ありがとうございました. 文 献
1) Couser WG. Mediation of immune glomerular injury. Journal
of Clinical Investigation 1993; 71: 808–811.
2) Masugi M. Die Allergie und Ihre Pathologische Bedeutung.
Transactiones Societatis Pathologiae Japonicae 1939; 29:
603–631.
3) Dixon FJ, Wilson CB, Marquardt H. Experimental Immuno-logic Glomerulonephritis. In: Hamburger J, Crosnier J, Max-well M, editors. Advances in Nephrology from the Necker
Hos-pital. Chicago: Year Book Medical Publishers Inc, 1971: 1–10.
4) Walport MJ. Complement: First of two parts. New England
Journal of Medicine 2001; 344: 1058–1066.
5) Brown KM, Sacks SH, Sheerin NS. Mechanisms of disease: the complement system in renal injury-new ways of looking at an old foe. Nature Clinical Practice Nephrology 2007; 3: 277– 286.
6) Pangburn MK, Meler-Eberhard HJ. Relation of a putative thioester bond in C3 to activation of the alternative pathway and the binding of C3b to biological targets of complement.
Journal of Experimental Medicine 1980; 152: 1102–1114.
7)藤田宜是,福家吉伸,松本紘一.補体系の活性化と腎組 織障害―慢性腎臓病における最近の展開―.日大医学雑 誌,2008; 67: 272–280. 8)大井洋之,関 正人,藤田宜是,小島弘之,波多野道 信.補体系と腎炎.腎と透析,1984; 17:::::785-791. 9)藤田宜是.腎疾患に於ける C3 分解産物測定の臨床的意義 とその経時的変動からみた補体活性の検討.日本腎臓学 会雑誌,1982; 24: 13–21.
10) Kojima H, Ohi H, Miyaji H, Seki M, Fujita T, Hatano M. Lo-calization of C3d in renal tissues of patients with membranous nephropathy and IgA nephropathy. The Japanese Journal of
Nephrology 1987; 29: 1161–1165.
11) Ohsawa I, Ohi H, Endo M, Fujita T, Kanmatsuse K, Nonaka M. Novel estimate of histologic activity in human glomerulone-phritis by detection of complement components C3 messenger RNA. Clinical and Experimental Nephrology 1998; 2: 50–57.
Patients with C3 Nephritic Factor. Nephron 1987; 47: 217–218. 15) Ohi H, Watanabe S, Fujita T, Seki M, Hatano M. Detection of C3bBb-stabilizing activity (C3 nephritic factor) in the serum from patients with membranoproliferative glomerulonephritis.
Journal of Immunological Methods 1990; 31: 71–76.
16) Ohi H, Watanabe S, Fujita T, Yasugi T. Significance of C3 ne-phritic factor (C3NeF) in non-hypocomplementemic serum with membranoproliferative glomerulonephritis (MPGN).
Clinical and Experimental Immunology 1992; 89: 479–484.
17) Tamano M, Fuke Y, Endo M, Ohsawa I, Fujita T, Ohi H. Uri-nary complement factor H in renal disease, Nephron 2002; 92: 705–707.
18) Endo M, Fuke Y, Tamano M, Hidaka M, Ohsawa I, Fujita T, Ohi H. Glomerular Deposition and Urinary Excretion of Complement Factor H in Idiopathic Membranous Nephropa-thy, Nephron Clinical Practice 2004; 97: c147–c153. 19) Fuke Y, Fujita T, Satomura A, Endo M, Matsumoto M. The
role of complement activation, detected by urinary C5b-9 and urinary factor H, in the excretion of urinary albumin in cisplatin nephropathy. Clinical Nephrology (in press).
20) Fujita T, Satomura S, Hidaka M, Ohsawa I, Endo M, Ohi H. Inhibitory Effect of Free Sialic Acid on Complement Activa-tion and its Significannce in Hypocomplementemic Glomeru-lonephritis, Journal of Clinical Laboratory Analysis 1999; 13: 173–179.
21) Endo M, Ohi H, Ohsawa I, Fujita T, Matsushita M, Fujita T. Glomerular deposition of mannnose-binding lectin (MBL) in-dicates a novel mechanism of complement activation in IgA nephropathy. Nephrology Dialysis Transplantation 1998; 13: 1984–1990.
22) Ohsawa I, Ohi H, Endo M, Fujita T, Matsushita M, Fujita T. Evidence of lectin complement pathway activation in post-streptococcal glomerulonephritis. Kidney International 56: 1158–1160.
23) Endo M, Ohi H, Ohsawa I, Fujita T, Matsushita M, Fujita T. Complement activation through the lectin pathway in patients with Henoch-Schlein purpura nephritis. American Journal of
Kidney Diseases 2000; 35: 401–407.
24) Endo M, Ohsawa I, Ohi H, Fujita T, Matsushita M, Fujita T. Mannose-binding lectin contributes to glomerulonephritis
in-27)藤田宜是,和田由紀,高橋佐智子,福家吉伸,里村厚 司,松本紘一.糖尿病性腎症におけるアシル化刺激蛋白 の生成と補体活性化. Therapeutic Research 2007; 28: 1912– 1914. 28) 藤田宜是,和田由紀,中根千枝,福家吉伸,里村厚司, 松本紘一.肥満関連腎症における脂質代謝異常や慢性炎 症の病態の腎障害への関与.日本腎臓学会誌,2008; 50: 251–251. 29)藤田宜是,大井洋之,八杉忠男.インスリン非依存性糖 尿病患者における補体の変動と腎における補体の活性 化.糖尿病,1992; 35: 977–983.
30) Fujita T, Ohi H, Komatsu K, Endo M, Ohsawa I, Kanmatsuse K. Complement Activation Accelerates Glomerular Injury in Diabetic Rats. Nephron 1999; 81: 208–214.
氏名:藤田 宜是 (ふじた たかゆき) 所属:日本大学医学部内科学系腎臓高血圧内分泌内科 学分野 主な研究領域:腎臓病学 (糸球体腎炎,糖尿病性腎炎, ループス腎炎の発症機序の解明と進展阻止). 抱負:今後も病気で苦しむ人々のために日夜,診療・ 研究・後進の指導に励む所存です.