1〒 606–8502 京都市左京区北白川追分町 京都大学大学院理学研究科生物科学専攻 2〒 819–0395 福岡市西区元岡 744 九州大学大学院工学研究院附属循環型社会システム工学研究センター 3〒 702–8021 岡山県岡山市南区福田 194 岡山淡水魚研究会 4〒 606–8501 京都市左京区吉田下阿達町 46 京都大学大学院アジア・アフリカ地域研究研究科 東南アジア地域研究専攻 生態環境論講座 (2014 年 3 月 20 日受付 ; 2014 年 7 月 25 日改訂 ; 2014 年 7 月 28 日受理) キーワード:アユモドキ,氾濫原,水田生態系,保全,PVA,京都府亀岡市 Japanese Journal of Ichthyology © The Ichthyological Society of Japan 2014
Katsutoshi Watanabe*, Hidetaka Ichiyanagi, Tsukasa Abe and Akihisa Iwata. 2014. Population viability analysis for the endangered loach Parabotia curtus in the Lake Biwa– Yodo River system, central Japan. Japan. J. Ichthyol., 61 (2): 69–83.
Abstract The population of the botiid Parabotia curtus in the Katsura River, Lake Biwa–
Yodo River system, Kameoka, Kyoto Prefecture, Japan, the only one recorded in the Kinki region since the year 2000, was subjected to a population viability analysis (PVA) based on demographic data between 2006 and 2013 (8 years). Count-based and age-structured models with several conditions for density-dependence, carrying capacity, migration rate, and bias in population estimation (48 settings in total) were used for calculating the quasi-extinction probability within 50 years (extinction threshold <10 individuals), using computer simulations. The estimated population for the 8 year period fluctuated from 200 to 2,300 individuals (average ca. 935, with a coefficient of variation of 66%). PVA demonstrated a significant extinction risk for this population (23–82% extinction in 45 of 48 settings). Although further population monitoring is necessary for a more precise evaluation, we conclude that this population faces a real extinction risk. Sensitivity analysis suggested several requirements for effectively enhancing population viability, i.e., restraining large biotic and abiotic environmental perturbations to avoid extreme depression of the recruitment of young, enhancing winter survival, and promoting upstream migration into the spawning site.
*Corresponding author: Division of Biological Sciences, Graduate School of Science, Kyoto University, Kitashirakawa-Oiwakecho, Sakyo, Kyoto 606–8502, Japan
(e-mail: [email protected])
自
然環境の変動や人為的な影響により個体群 の縮小や絶滅が危惧される生物に対して, その個体群変動機構を明らかにし,動態予測を行 うことは,適切な保全策を立案,実施するうえで も,また今後の人為的影響の程度を評価するうえ でも重要である.野外個体群の中・長期データに 基づく個体群動態モデルと,それをもとにした個 体数変動シミュレーションは,個体群存続可能性 分 析(population viability analysis; PVA) と し て 多くの希少生物や資源生物に対して適用され,その 結果は定量的あるいは相対的な評価基準として 用いられてきた(Beissinger and McCullough, 2002; Morris and Doak, 2002 等参照).対象生物に関する 知見の蓄積の程度により,個体群動態モデルの適 合性や予測性は異なる.しかし,それぞれの時点 での最善の情報とモデルにより分析を実現するこ とは,保全対策の機を逸さないために重要であ るばかりではなく,さらに必要なデータ項目を
琵琶湖・淀川水系のアユモドキ個体群の存続可能性分析
渡辺勝敏
1・一柳英隆
2・阿部 司
3・岩田明久
4抽出し,モデルを改善するなど,保全策を適応 的に推し進める過程としても重要である(例え ば,Akçakaya et al., 1999; Beissinger and McCullough, 2002; Frankham et al., 2002). コイ目アユモドキ科アユモドキ Parabotia curtus は,日本固有の純淡水魚であり,琵琶湖・淀川水 系と山陽地方にのみ分布が局限される希少魚である (前畑,2003;岩田,2006a;阿部・岩田,2007). アユモドキは雨季に河川周辺の氾濫原に形成され る一 時 的 水 域 に遡 上し, 産 卵を行う(Abe et al., 2007a, b).人為的な水文管理や河川整備が進んだ 今日,アユモドキが遡上し,繁殖,初期発育を行 える環境はほとんど消失してしまい,現在本種の繁 殖がまとまって見られるのは,琵琶湖・淀川水系で 1 ヶ所, 山 陽 地 方 で 2 ヶ所 の み で ある( 片 野, 1997;阿部・岩田,2007).アユモドキは種として 国の天然記念物に指定されるとともに,環境省の レッドリストでは絶滅危惧 IA 類とされ,「絶滅のお それのある野生動植物の種の保存に関する法律 (種の保存法)」で国内希少野生動植物種に指定さ れている.また「京都府絶滅のおそれのある野生 生物の保全に関する条例」においても指定希少野 生生物として選定されている.本種は日本の魚類 の中でも最も保全優先度の高い種の一つである. アユモドキの残存する 3 つの個体群のうち近畿 地方唯一の個体群は,京都府亀岡市を流れる桂川 の一支流で繁殖していることが知られている(岩 田,2006a).この生息地は,桂川が保津峡で狭窄 される直上流部に位置する亀岡盆地に存在し,洪 水時に遊水機能を果たしてきた場所にある.現在, アユモドキの繁殖は,桂川との合流部近くにある灌 漑用ゴム布引製起伏堰(以下,可動堰とよぶ)の 起立によって上流部に生じる河道内の一時的水域 で行われ,その一時的水域や周辺の農業用水路が 本種の初期発育場所として利用されている.しか し,アユモドキの繁殖や初期発育は次の理由に よって,決して安定的なものではない.まず,まと まった繁殖場所がこの 1 地点のみであること,堰上 げ後は可動堰が移動障害物として機能すること, 降雨時に防災目的で急激な堰の転倒操作が行われ る場合があること,捕食性外来種が周辺のため池 から侵入する可能性があること,水田から急速に水 が落とされ,用水路が干上がること,などのためで あ る( 阿 部・ 岩 田,2007; 岩 田,2009). ま た 2013 年より,この生息地を含む農地約 14 ha が京 都府の専用球技場として大規模に開発される計画 が進められており,さらに周辺地域の開発・土地 利用変更や圃場整備などの進行に対して,保全対 策の必要性が高まっている(亀岡市,2012;日本 魚類学会,2013;上原,2013).本生息地におけ る個体数変動機構を明らかにし,本個体群の存続 可能性と脆弱性を評価する必要性は大きい. 著者の一人(AI)が属する保全・研究グループ (NPO 法人亀岡 人と自然のネットワーク)は,文 化庁,環境省,および京都府の許可を得たうえで, 2006 年以降,桂川におけるアユモドキの生息状況 を明らかにするために,繁殖期の終了後,可動堰 上流部で当歳魚および高齢魚の個体数のモニタリ ング 調 査 を 行ってきた. そこで, 本 研 究 で は, 2006–2013 年の 8 年間のデータを用い,複数の個 体数変動モデルを作成し,本個体群の存続可能性 分析を行った.その結果,本個体群の存続が不安 定な状況にあり,特に秋から次年の遡上時までの 生残率の改善が安定した存続のために求められるこ となどが明らかになったので,今後の本個体群の 保全方針に関して議論を行う. 材 料 と 方 法 調査場所と対象個体群 個体数推定および存続 可能性分析を行ったアユモドキ個体群は,近畿地 方,琵琶湖・淀川水系で唯一残る個体群であり, 京都府亀岡市を流れる桂川支流の T 川(関係者間 の申し合わせで河川名を伏す)とその周辺の農業 用水路,そして合流部周辺の桂川本流に生息する. 毎年 6 月初旬に,桂川との合流部より約 50 m に位 置する可動堰を起立後,普段は干出している堰上 流 部の植 物 帯を中 心に繁 殖 が 行 われる(岩 田, 2006a).堰の起立直後に,堰の下流に取り残され た個体の多くが,保全活動として人為的に上流に 移されている.体サイズに基づいて,雌雄とも 1 歳 の一部と 2 歳以上で成熟し,数年間は生きると推 察されている(片野,1997;岩田,2006a).仔稚 魚は,T 川および可動堰上部で合流する B 川,そ して B 川と通じた農業用水路に分散し,初期生活 史を送る.7 月末に一時的な水田の落水(土用干 し),また 9 月下旬に可動堰の転倒を行うため,農 業用水路の水が激減または消失し,この際にもでき る限り,人為的に稚幼魚を T 川へ移動させている. 可動堰の転倒後,秋季から冬季はおもに合流部に 近い本流で過ごすと考えられているが詳細は不明 である(京都府南丹土木事務所,未発表データ). 夏季にも未成熟個体を含む堰上流部に侵入できな かった個体が,わずかに本流で確認される(岩田,
2007;京都府南丹土木事務所, 未発表データ). また冬季に T 川の一部区間においてアユモドキが 確認されることもまれにある(岩田,2006b). 個体数推定 個体数推定は,2006 年から 2013 年の 8 年間,毎年,繁殖期が終わり,可動堰の転 倒が行われた 1–2 日後の 9 月中旬から下旬にかけ て,Petersen 法による標識再捕法によって行われた. 桂川と T 川の合流点から T 川上流の約 250 m の範 囲で,2 回に分け,毎年同様の方法と努力量により アユモドキの隠れ場所を悉皆的に調査し,アユモド キを手網で一時捕獲した(Table 1).1 回目の放流 個体については,いずれかの鰭の一部を切除し, 標識とした.捕獲個体を体サイズ分布や体色など から当歳魚とそれ以外(高齢魚)の 2 群に分け, それぞれ Chapman の式(伊藤・村井,1977 参照) によって,個体数を推定した. ただし,n1は 1 回目の標識放流個体数,n2は 2 回 目の捕獲個体数,m2は 2 回目の標識個体の再捕 個体数である. 標識再捕推定が可能であった年(当歳魚は全 8 年,高齢魚は 8 年のうち 7 年)について,推定個 体数と 2 回の平均捕獲数の比を当歳魚と高齢魚別 に推定した(∑推定個体数/ ∑平均捕獲数).再捕 個体が得られなかった回(2009 年の高齢魚のみ) については,Chapman の式からの推定値ではなく, この比(高齢魚 6.9)を平均捕獲数に乗じて個体数 を推定し,それの標準誤差は求めなかった.また 2008 年には当歳魚の捕獲が 0 個体であったが,後 の計算上,対数をとる必要があるので,これを 1 個体とした.この当歳魚の激減は,2008 年の繁殖・ 初 期 発 育 期 に 近 隣 の た め 池 からオオクチ バ ス Micropterus salmoides が多数侵入した結果であると 推定されている(岩田,2009). 個体数推定調査は可動堰の転倒直後(基本的に 1–2 日間)であるが,一部の個体がすでに流下して いる可能性を否定できないため,個体数が過小評 価されている可能性がある.したがって,個体数が 推定の 1.5 倍と 2 倍である場合についても,以下の モデル作成とシミュレーションを行った. 個体数変動モデル 個体数変動モデルとして, 大きく2 つのタイプのモデルを作成した.1 つは年 齢を区別しない「個体数ベースモデル count-based model」(モデル A),もう 1 つは当歳魚と高齢魚の 齢 構 成 を 考 慮 に 入 れ た「 齢 構 造 モ デ ル age-structured model」( モ デ ル B) で ある(Morris and Doak, 2002 参照).それぞれさらに 2 つのモデルを 以下のように構築した.なお,今回すべてのパラ メータの推定において,個体数推定上の誤差は取
Table 1. Mark-release-recapture data, estimated numbers of individuals and population parameters
of Parabotia curtus in Kameoka, Kyoto Prefecture
0+ ≥1+ n1 n2 m2 N SE n1 n2 m2 N SE r r´ S0 2006/9/16, 17–9/24 105 54 8 647 179 64 24 5 270 85 -0.1390 0.8739 0.6830 2007/9/15–9/23 10 12 1 71 34 102 98 13 727 162 -1.2839 -2.3262 0.1615 2008/9/25–23 0 0 0 1 0 79 46 16 220 37 2.3490 -5.3936 0.2006 2009/9/16, 17, 19–9/26 358 267 42 2,236 290 15 8 0 79a – -0.6979 3.3430 0.3079 2010/9/18–9/25 27 34 1 489 265 143 105 22 663 110 -0.3218 -0.3044 0.3276 2011/9/17–9/24 44 26 3 303 120 73 71 9 532 139 -0.3765 -0.5629 0.4482 2012/9/17–9/23 15 7 4 25 5 79 47 6 548 171 0.1594 -3.0874 0.5115 2013/9/23–9/29 20 22 2 160 69 34 43 2 512 237 -1.1632 Average 492 444 -0.0444 -1.0776 0.3772 SD 741 228 1.1496 2.6308 0.1833
n1: Number of marked and released individuals on first day; n2: number of individuals captured on second day; m2: number of
marked individuals; N and SE: number of individuals and standard error estimated by Chapman’s method. For estimated popu-lation parameters (r, r´ and S0), see Methods. S0 shown when u = 0.9 (proportion of individuals entering spawning site). a Obtained from average number of n
1 and n2 multiplied by ratio (6.9) of average catch to estimated number of ≥1+ individuals
り込まなかった.ただし, 個体数推定の誤差は, シミュレーションに基づく個体数変動の幅を大きく し,絶滅確率の過大評価につながるため,その影 響については別途検討した(考察参照). モデル A:個体数ベースモデルでは,まず 8 年 間の推定個体数(N)に基づき,連続する 2 年間 (t, t + 1)の瞬間増加率 rt = ln (Nt+1/Nt) の平均値 r _ と 標準偏差 σrを求めた.ただし,Ntは 9 月時点での 可動堰上部での生息数である.そして,指数成長 型モデル Nt+1 = er · Ntに対して,r を平均 r _ と標準偏 差 σrの正規乱数として与えた(モデル A-1).この rのばらつきには瞬間増加率に対する年間の環境 変動による確率性と人口学的確率性が含まれてい る.瞬間増加率に時間的な自己相関はないものと した.ただし,個体群の非現実的な成長を調節す るため,個体数に上限を与えた.上限は実際に観 察された上限値(約 2,300) の約 2 倍(5,000)と 約 10 倍(20,000) の 2 つ の 場 合を 検 討した. な お,個体数を推定値の 1.5 倍,2 倍と仮定した場合 にも,上限値を同様にそれぞれの 2 倍,10 倍に増 加させ,検討した.また極端な低密度下での存続 性の低下を組み入れるために,人口学的確率性や 遺伝的浮動による遺伝的多様性の喪失が顕著とな る任意の値として,10 個体未満となったときに絶滅 したとみなした.そのため,人口学的確率性や遺 伝的多様性の喪失については,モデルに明示的に 組み入れなかった. 次に, 高密度下での負の密度効果を考慮に入 れ,瞬間増加率 r が前年個体数と単純な負の傾き をもつ直線関係を示すモデルを作成した(ロジス ティックモデル;モデル A-2).まず実際のデータか ら得られた Ntと rtの一次回帰式を最小二乗法で求 め,実際のデータとの残差の平均と標準偏差を求 めた.A-1 と同型の指数成長型モデルにおいて,あ る年の r を,その年の個体数をもとに,回帰直線 と正規乱数とした残差によって決定した.ただし個 体数の上限値と絶滅基準を上記と同様に与えた. また,個体群増加,すなわち r が 0 となる Ntの値 KA(環境収容力)を求めた. モデル B:齢構造モデルでは,当歳魚(0+)と 高齢魚(≥ 1+)の 2 つの齢を考えた.まず当歳魚 はその年の高齢魚から出生するので, ただし,r′ は高齢魚の個体あたりの繁殖率,N0+,tと N≥ 1+,tは 9 月時点での可動堰上部での当歳魚と高齢 魚の生息数である.ここで高齢魚は繁殖後の生存 個体なので,繁殖期当初の個体数よりも死亡により 減少している可能性があるが,大きくは異ならな い,もしくは死亡率が年によって大きく変化しない ものとみなした.モデル B-1 では,実際のデータか ら,r′ の平均値と標準偏差を求め,正規乱数とし て与えた.モデル B-2 では,r′ をモデル A-2 と同 様に回帰直線の周囲でばらつく正規乱数とし,負 の密度効果を組み入れた. また, 高齢魚の個体数は,モデル B-1,B-2 の いずれにおいても次式で表される. ここで,S0は当歳魚の翌年までの生残率,S1は高 齢魚の翌年までの生残率である.u は繁殖場所に 遡上した個体の割合である.u は不明であるが,夏 季に本流で見られる数はわずかであるので(京都 府南丹土木事務所,未発表データ),0.9,0.75,0.5 の 3 通りを仮定した.年齢による u の変化は,モ デルで 1 歳以上の年齢を区別しておらず,また野 外データもないため,考慮しなかった.生残率に ついては,個体数の時系列データから,まず重回 帰分析により上式のような原点を通る直線として Ŝ0 と Ŝ1を推定し,その比 q = Ŝ1/Ŝ0を求めた.そして, 各年の個体数データを用いて,上式に基づき,S1 = q · S0として各年の S0を計算し,それと重回帰分析 から求められた Ŝ0の残差の平均と標準偏差を求め た.この平均と標準偏差をもつ正規乱数を,Ŝ0の 変動としてモデルに組み入れた.また,この乱数に より生成した S0に連動させて,S1(= q · S0)を生成 した.ただし,S0の下限および上限は 0.01 と 0.95, S1の上限は 0.95 とした.生残率に関する密度効果 は,実際のデータにおいて観察されなかったので, モデルに組み入れなかった.B-1,B-2 いずれにお いても,当歳魚の個体数の上限を実際に観察され た上限値(約 2,200)の約 2 倍(4,000)と約 10 倍 (20,000),また繁殖場所に遡上した高齢魚の個体 数の上限を実際に観察された上限値(約 800)の 約 2 倍(1,500)と約 10 倍(8,000)の各場合で検 討した.各齢の 8 年間の個体数の平均値に基づい て,個体数増減が 0 となるときの繁殖場所に遡上 した高齢魚の個体数 N≥ 1+,t(= KB)を,力まかせ探 索法(brute-force search),つまり周辺値に関して網 羅的な数値計算を行うことにより求めた. 存続可能性分析と感度分析 存続可能性の基準 として,25 年後および 50 年後の絶滅確率を用いた. シミュレーションの初期値は,いずれも 8 年間の平 均値を用いた(モデル A-1,A-2 では合計 935,モ デ ル B-1, B-2 で は 当 歳 魚 491, 遡 上した 高 齢 魚 444).シミュレ ーションは Microsoft Excel for Mac
2011 のスプレッドシート上で行い,それぞれのモデ ルや条件下で各 10,000 回の繰り返しを行った. モデル B1 と B2 において,シミュレーションで 50 年間絶滅しなかった場合について,有効集団サイ ズの指標として,50 年分の高齢魚の個体数の調和 平均を計算した(Frankham et al., 2002).これは, 性比や繁殖成功の個体間の偏りのために,実際の 有効集団サイズよりも過大と考えられ,有効集団サ イズの上限値とみなすことができる. パラメータの推定誤差による絶滅確率推定への 影響の評価,および有効な保全対策の検討のため に,感度分析を行った.前者に関しては,各パラ メータやそのばらつきの 95% 信頼区間およびその 半分を基準に,それぞれのパラメータに関して単 独で変化させたときの 50 年後の絶滅確率の変化を 算出した.また各パラメータ単独で絶滅確率を 5% 未満にする閾値を求めた.モデル A-1 では r とそ の標準偏差について検討した.r の標準偏差につ いては,ブートストラップ法(10,000 回の無作為復 元抽出)で 95% 信頼区間を推定した.信頼区間の 推 定 に は, 偏りを 補 正した パ ー センタイル 法 (bootstrap bias-corrected percentile confidence limits)
を用いた(Manly, 2006: 52).モデル A-2 では r の 回帰式における傾きと切片,および回帰直線から の残差について検討した.ただし,傾きを変化さ せる場合には,Ntと rtの平均値を通る直線とし, 切片も同時に動かした.モデル B-1 では r′,S0,お よびそれらの標準偏差について検討した.両者の 標準偏差に関する 95% 信頼区間はブートストラッ プ法で推定した.B-2 では r′ における傾きと切片, および回帰直線からの残差,S0とその標準偏差に ついて検討した.r′ における傾きは,N≥ 1+,tと rt′ の 平均値を通る直線について変化させ,S0の標準偏 差についてはブートストラップ法で 95% 信頼区間を 推定した. 有効な保全対策の検討のための感度分析におい ては,パラメータ空間における 50 年後の絶滅確率 の高低パターンを調べた.各モデルで検討したパ ラメータは上記と同様である.ただし,密度依存モ デル(A-2 と B-2)では,個体数増加が 0 となる Nt (KA)あるいは N≥ 1+,t(KB),つまり環境収容力につ いて,その値を変化させて,絶滅確率の変化を調 べた.保全策におけるパラメータの変化のさせや すさについては,関係するパラメータ値の実際の 観察値やばらつきを基準に判断した. 結 果 個体数変動 2006 年から 2013 年にかけて, 当 歳 魚 が 0–2,236 個 体( 平 均 ± 標 準 偏 差,492 ±
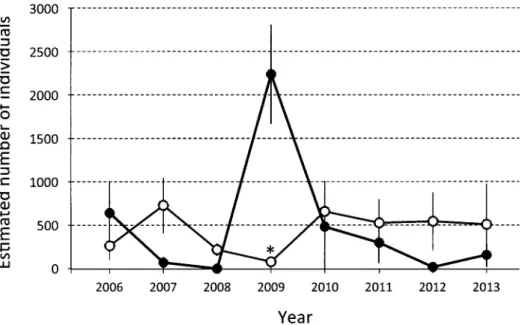
Fig. 1. Fluctuations of the Parabotia curtus population in Kameoka, Kyoto Prefecture. The estimation
was conducted in September each year for young-of-the-year (solid circles) and older individuals (open circles) using a mark–release–recapture method. Bars indicate 95% confidence intervals. *Confidence interval not estimated (see text and Table 1).
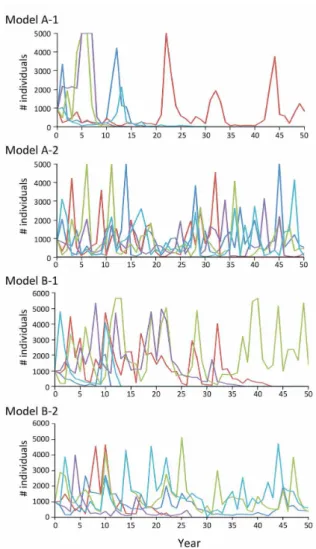
741; 変 動 係 数,151%), 高 齢 魚 が 79–727 個 体 (444 ± 228;51%), 合 計 で 221–2,315 個 体(935 ± 620;66%)が生息していたと推定された(Fig. 1; Table 1).8 年 間 を 通して, 当 歳 魚 の 割 合 は 52.5%, 高齢魚の割合は 47.5% であった. 高齢魚 の個体数の調和平均は 272 個体であった.標識再 捕による推定個体数と 1 回あたりの捕獲数の比は, 当歳魚で 7.9,高齢魚で 6.9 であった. 個体数変動モデル 8 年間のデータに基づく推 定 全 個 体 数 の 平 均 瞬 間 増 加 率 r_( ± SD) は, –0.0444(± 1.1496)であった(Table 2).そこで, モデル A-1 の指数成長型モデルにおいて,r の正 規乱数のパラメータとしてこれらの値を用いた. 全個体数 Ntと瞬間増加率 r は,r = –0.000973 Nt + 0.9020 の関係を示し(Fig. 2a),実際のデータの 残 差 は 平 均 0, 標 準 偏 差 0.9539 を示した(Table 2).そこでモデル A-2 の指数成長型モデルにおけ る r を,この回帰式の周りで正規分布 N(0, 0.9539) でランダムに変動する値とした.KAは 927 であった. 齢構造モデルにおける高齢個体群の個体数 N≥ 1+,t に基づくN0+,tの平均繁殖率 r _ ′ (± SD)は,–1.0776(± 2.6308)であった(Table 2).モデル B-1 における r′ をこれらのパラメータをもつ正規乱数とした.また r′ は N≥ 1+,tと,u = 0.9 の と き,r ′ = –0.003672 N≥ 1+,t + 0.5522 の関係を示し(Fig. 2b),実際のデータの残 差は,平均 0,標準偏差 2.4936 を示した(Table 2). そこでモデル B-2 の当歳魚に関する指数成長型モデ ルにおける r′ を,この回帰式の周りで正規分布 N(0, 2.4936)でランダムに変動する値とした. モデル B-1 および B-2 においては,当歳魚と高 齢魚の生残率 S0と S1は,遡上率 u に応じて,重回 帰分析により決定され,例えば u = 0.9 の場合,順 に 0.3545,0.6331 と推定された(q = 1.7856;Table 3).このとき各年の S0はこの回帰式から残差平均 0.0229, 標 準 偏 差 0.1834 でばらつ いていたので, シミュレーションにおいて,S0は回帰式の周りを正 規分布 N(0.0226, 0.1833)でランダムに変動する値 とした.S1は S0と q から決定した.個体数増減は r′ = 0.1397 のときに 0 であり,このとき KBは 112, 安 定 齢 構 成は当 歳 魚 0.509:高齢魚 0.491 であっ た.またこのときの個体群全体のサイズは u に応じ て 970–990 個体であり, モデル A-2 の KA(= 927) と近い値を示した. 存続可能性分析と感度分析 4 つの個体数変動 モデルのもとで存続可能性分析を行ったところ(Fig. 3),絶滅確率は増加率に負の密度効果を取り入れ な かった 場 合(A-1 と B-1)に,取り入 れ た 場 合 (A-2 と B-2)よりも高い値を示した(Table 3).前者 では 50 年絶滅確率が各設定で 70–80%(A-1;Table 3 のケース 1–2)および 65–82%(B-1;ケース 5–10) と近い値を示し,後者では,1–32%(A-2;ケース 3–4)および 26–42%(B-2;ケース 11–16)であった. 低密度下で大きな瞬間増加率を示すモデルである A-2 において低めの個体数上限値(5,000)を設定 した場合(ケース 3)を除いて,いずれも無視でき ない大きさの絶滅確率を示し(23–82%),つまり個 体数推定の過小評価や遡上率に関する条件設定に 対して頑強な結果であった(Table 3). モデル A-1 と B-1 においては,25 年絶滅確率も 45–55% と高い値を示した(Table 3).モデル B1 と B2 において,絶滅しなかったときの 50 年間の個体 数(高齢魚)の調和平均は 173–632(B-1;Table 3 の ケ ース 5–10)と 157–296(B-2; ケ ース 11–16) であり,それぞれにおいて,個体数の上限値が大 きく,遡上個体数が全個体数よりも少ないとみなし
Fig. 2. Relationship between population size and
instantaneous growth rate in the Parabotia curtus population in Kameoka, Kyoto Prefecture. (A) Model A-2, regression line: r = –0.000973 Nt + 0.9020 (R2 = 0.3116). (B) Model
B-2, regression line: r´ = –0.003672 N≥1+,t + 0.5522 (R2 =
Table 2.
PV
A
parameters estimated from the
Parabotia curtus
population in Kameoka, Kyoto Prefecture, and results of sensitivity analysis
±1/2 • 95% C.I.
±95% C.I.
Value needed for
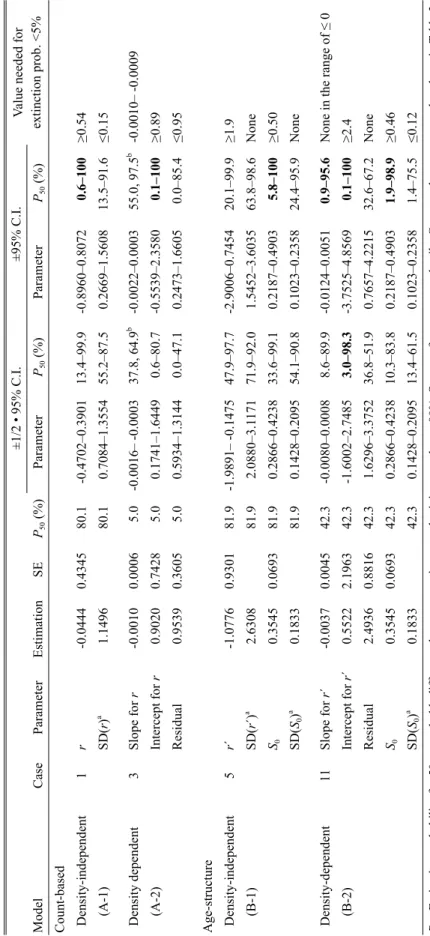
extinction prob. <5% Model Case Parameter Estimation SE P50 (%) Parameter P50 (%) Parameter P50 (%) Count-based Density-independent 1 r -0.0444 0.4345 80.1 -0.4702–0.3901 13.4–99.9 -0.8960–0.8072 0.6–100 ≥0.54 (A-1) SD( r) a 1.1496 80.1 0.7084–1.3554 55.2–87.5 0.2669–1.5608 13.5–91.6 ≤0.15 Density dependent 3 Slope for r -0.0010 0.0006 5.0 -0.0016– -0.0003 37.8, 64.9 b -0.0022–0.0003 55.0, 97.5 b -0.0010– -0.0009 (A-2) Intercept for r 0.9020 0.7428 5.0 0.1741–1.6449 0.6–80.7 -0.5539–2.3580 0.1–100 ≥0.89 Residual 0.9539 0.3605 5.0 0.5934–1.3144 0.0–47.1 0.2473–1.6605 0.0–85.4 ≤0.95 Age-structure Density-independent 5 r´ -1.0776 0.9301 81.9 -1.9891– -0.1475 47.9–97.7 -2.9006–0.7454 20.1–99.9 ≥1.9 (B-1) SD( r´) a 2.6308 81.9 2.0880–3.1 171 71.9–92.0 1.5452–3.6035 63.8–98.6 None S0 0.3545 0.0693 81.9 0.2866–0.4238 33.6–99.1 0.2187–0.4903 5.8–100 ≥0.50 SD( S0 ) a 0.1833 81.9 0.1428–0.2095 54.1–90.8 0.1023–0.2358 24.4–95.9 None Density-dependent 11 Slope for r´ -0.0037 0.0045 42.3 -0.0080–0.0008 8.6–89.9 -0.0124–0.0051 0.9–95.6
None in the range of ≤ 0
(B-2) Intercept for r´ 0.5522 2.1963 42.3 -1.6002–2.7485 3.0–98.3 -3.7525–4.8569 0.1–100 ≥2.4 Residual 2.4936 0.8816 42.3 1.6296–3.3752 36.8–51.9 0.7657–4.2215 32.6–67.2 None S0 0.3545 0.0693 42.3 0.2866–0.4238 10.3–83.8 0.2187–0.4903 1.9–98.9 ≥0.46 SD( S0 ) a 0.1833 42.3 0.1428–0.2095 13.4–61.5 0.1023–0.2358 1.4–75.5 ≤0.12 P50 : Extinction probability for 50 years; bold: dif ference between maximum and minimum values >90%. See text for parameter details. Case numbers correspond to those in Table 3.
u = 0.9. a Confidence intervals estimated as bias-corrected percentile c
onfidence limits based on 10,000 bootstrap resamplings.
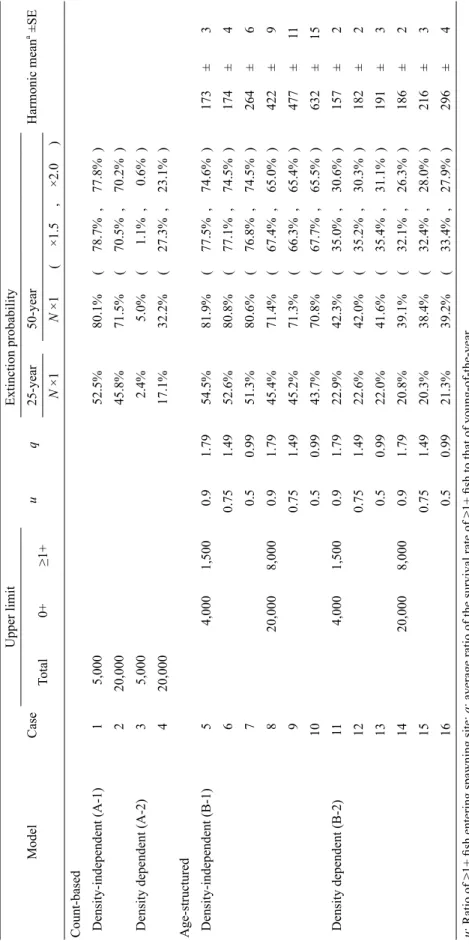
Table 3.
Extinction probability in four PV
A
models with several sets of parameters for the
Parabotia curtus
population in Kameoka, Kyoto Prefecture
Model Case Upper limit u q Extinction probability Harmonic mean a ±SE Total 0+ ≥1+ 25-year 50-year N ×1 N ×1 ( ×1.5 , ×2.0 )
Count-based Density-independent (A-1)
1 5,000 52.5% 80.1% ( 78.7% , 77.8% ) 2 20,000 45.8% 71.5% ( 70.5% , 70.2% )
Density dependent (A-2)
3 5,000 2.4% 5.0% ( 1.1% , 0.6% ) 4 20,000 17.1% 32.2% ( 27.3% , 23.1% ) Age-structured Density-independent (B-1) 5 4,000 1,500 0.9 1.79 54.5% 81.9% ( 77.5% , 74.6% ) 173 ± 3 6 0.75 1.49 52.6% 80.8% ( 77.1% , 74.5% ) 174 ± 4 7 0.5 0.99 51.3% 80.6% ( 76.8% , 74.5% ) 264 ± 6 8 20,000 8,000 0.9 1.79 45.4% 71.4% ( 67.4% , 65.0% ) 422 ± 9 9 0.75 1.49 45.2% 71.3% ( 66.3% , 65.4% ) 477 ± 11 10 0.5 0.99 43.7% 70.8% ( 67.7% , 65.5% ) 632 ± 15 Density dependent (B-2) 11 4,000 1,500 0.9 1.79 22.9% 42.3% ( 35.0% , 30.6% ) 157 ± 2 12 0.75 1.49 22.6% 42.0% ( 35.2% , 30.3% ) 182 ± 2 13 0.5 0.99 22.0% 41.6% ( 35.4% , 31.1% ) 191 ± 3 14 20,000 8,000 0.9 1.79 20.8% 39.1% ( 32.1% , 26.3% ) 186 ± 2 15 0.75 1.49 20.3% 38.4% ( 32.4% , 28.0% ) 216 ± 3 16 0.5 0.99 21.3% 39.2% ( 33.4% , 27.9% ) 296 ± 4
u: Ratio of ≥1+ fish entering spawning site;
q: average ratio of the survival rate of ≥1+ fish to that of youn
g-of-the-year
.
Population size assumed as 1, 1.5, or 2.0 times the estimation by a mark-release-recapture method (shown as
N
×1, ×1.5 and ×2, respectively).
a A
た設定で比較的大きい値を示した. 感度分析の結果,いくつかのパラメータにおけ る推定誤差が絶滅確率評価に大きな幅を与えるこ とが明らかとなった(Table 2).モデル A-1 では,r の 95% 信 頼 区 間 にお いて 50 年 絶 滅 確 率 が 0.6– 100% と大きく変わった.密度効果を入れた A-2 と B-2 では, 低密度下での r の値(回帰式の切片) が絶滅確率に大きな影響を与えた(いずれも 0.1– 100%). 齢構造モデル(B-1, B-2) における S0の 推 定 誤 差も, 大きく絶 滅 確 率の評 価に影 響した (順に 5.8–100%,1.9–98.9%).さらにモデル B-2 に おける r の回帰式の傾きの推定誤差も大きな影響 を示した(0.9–95.6%).ただし,これらほとんどの パラメータにおいて,信頼区間の幅を半分に狭め た場合(± 1/2 · 95% C.I.;Table 2)には絶滅確率 の推定幅が明瞭に狭まった.これらは,精度の高 い絶滅確率の評価のためには,さらに長い期間の 個体数変動データが必要であることを示す.一方, いくつかのパラメータでは,推定誤差にかかわらず 一貫して高い絶滅確率を示した.例えば,モデル B-1 の r′ については 95% 信頼区間において 20.1– 99.9%,B-2 の回帰式からの残差については同じく 32.6–67.2% と高い絶滅確率を示した.また変動性 に関するパラメータ(SD や残差)の推定誤差によ る絶滅確率評価への影響は,一部を除いて比較的 小さかった(Table 2). 各モデルにおいて,パラメータ値を連続的に変 化させ, 絶滅確率への影響を検討した(Fig. 4). モデル A-1 においては,平均 r を増加させ,r のば らつきを抑える,つまり安定して正の増加率を実現 することが 絶 滅 確 率を 下 げることにつ な がった (Fig. 4A).実際に観察された r のばらつきから,r が特に低い年が生じなければ,低い絶滅確率が実 現されると判断されたが,絶滅確率を 5% 未満にす るためには,7 年間のうち上位 3 位の r が必要で あった. モデル A-2 では, 環境収容力 KAの小さい方向 への変化に対して,明確な絶滅確率の上昇が予測 された(Fig. 4B).増加率の密度依存関係に関する ばらつき(残差)が大きいと KAにかかわらず絶滅 確率が明瞭に上昇し,最もばらつきの少ない 1 年 のデータを除くだけで,絶滅確率は 10% を超えた. r が全体傾向よりも極端に低い年を生じさせないこ とが絶滅確率を低下させるために必要であることを 意味する. モデル B-1 では,低い絶滅確率を実現するため に繁殖率 r′ の上昇が必須であるが(Fig. 4C),絶 滅確率を 5% 未満にするためには, 観察された 8 年のうち上位 2 位の r′ が達成される必要があった. 一方,生残率 S0については,7 年間のうち下位 2 位までの値を除くと絶滅確率は 5% 未満となり,相 対的に実現されやすいと判断された. モデル B-2 は,同じく密度依存モデルである A-2 と同様に,環境収容力 KBの小さい方向への変化 に対して明確な絶滅確率の上昇を示したが,大き い方向への変化に対してはあまり敏感ではなく,絶 滅確率はほとんど下がらなかった(Fig. 4B). 一 方,S0の 影 響 は KBにほとんど関 係 なく, およそ 0.45–0.50 以上(観察推定値 0.35)で絶滅確率は 5% 未満となり,7 年間の観察値の下位 2 位までを 除くことで達成された.
Fig. 3. Examples of PVA simulations on different models
for the Parabotia curtus population in Kameoka, Kyoto Prefecture. Total number of individuals (young-of-the-year plus older individuals) shown for B-1 and B-2 models.
考 察 個体数変動モデルと存続可能性分析の妥当性 アユモドキの個体群動態に関する解析は本研究が 初めてのものであり,地域の保全・研究グループ が活動の一環として行ってきた 8 年間の当歳魚と高 齢魚,つまり 2 齡構造の個体数データが,現在利 用できる最良のデータである.一般に個体数変動 モデルを構築する場合には,対象種の生物学に基 づく適切なモデルを適用し,精度の高い長期デー タからそのパラメータを得る必要がある.そのため には,より長期的な,またより詳しいコホート別の デ ータの 利 用 が 望ましく(Morris and Doak, 2002; Sæther and Engen, 2002),今後,さらに長期のデー タに基づく検証が必要である.しかし,以下に述べ るとおり,さまざまなモデルと条件を用いた解析の 結果から,今回の PVA 分析による現状評価と保全 策の検討は一定の頑健な結論を導くものである. 今回,4 タイプの個体数変動モデルを採用し, PVA 分析を実施した.それらは全個体数に基づく 単純な個体数ベースモデル(A)と 2 段階の齢構 造モデル(B)である.これらには個体数上限値を
Fig. 4. Results of PVA and sensitivity analysis for Parabotia curtus in Kameoka, Kyoto
Prefecture, represented as the distribution of extinction probabilities (within 50 years) in several parameter spaces. See text for parameter details. Case numbers correspond to those in Table 3. Cells surrounded by bold lines correspond to observed data. Approximate ranges of observed values, standard deviations (SD) and 95% confidence intervals for parameters given as bars when available.
与えたのに加え,A-2 と B-2 として,負の密度効果 (単純な個体数と瞬間増加率または繁殖率の間の 負の直線関係)を与えた.個体数の推定誤差を含 む時系列データにおいては,Ntと Nt+1/Ntに負の相 関関係を生じる偏りがあるので,密度効果の取り込 みに関しては検討が必要である.実際にモデル A において, 密度効果がないと仮定した場合でも, 個体数推定の誤差を考慮すると,観察された直線 回帰の傾き(–0.000972)の半分程度の傾きが生じ ると試算された(未発表データ).したがって,現 在のデータは瞬間増加率における負の密度効果を 積極的に支持するものではない.しかし,データ 年数が短いために検出力が低いことを考慮すると, 4 タイプすべての結果を相互比較しながら,PVA の 結果とその一致性を検討するのがよいと思われる. この場合,低密度下での高い瞬間増加率・繁殖率 Fig. 4-2
を与える A-2 と B-2 は, 絶滅リスクの評価におい て,より楽観的な結果とみなすことができ,密度効 果が小さいときにはそれぞれ A-1 と B-1 に近づく. 別の密度効果として,本研究ではすべてのモデ ルにおいて,観察された個体数の 2 倍または 10 倍 の高めの上限値を与えた.モデル A-2 を除いて, 低い上限値は絶滅確率を上昇させたが,いずれの 条件においても絶滅リスク評価において極端に大き な違いをもたらさなかった(Table 3).また,本研 究では絶滅基準を 10 個体未満と設定した.これを 厳密に 2 個体未満とした場合,例えば A-1(ケース 1)と B-1(ケース 5)では,絶滅確率が順に 65% (vs. 80%),66%(vs. 82%)といくらか下がる.しか し,個体群サイズがきわめて縮小したときに人口学 的・環境的確率性やアリー効果,遺伝的多様性の 喪失などの効果が増大することを考えると,実質的 な絶滅基準を 10 個体未満と置くことに無理はない だろう. 標識再捕に基づく個体数推定誤差は,個体数変 動のばらつきを過大評価することを通じて絶滅確率 の 過 大 評 価 をもたらす 可 能 性 が ある( 例 えば,
Longergan and Harwood, 2003).しかし,例えばモデ ル A-1 における瞬間増加率の標準偏差(1.1494) は個体数推定誤差を含んでいるが,その影響を除 去した推定値,すなわち誤差伝播則を用いた補正 (Morris and Doak, 2002)による値(1.10)や力まか せ探索法による推定値(1.11;未発表データ)と大 きくは異ならず,絶滅確率に大きな影響を与えるも のではなかった(Fig. 4A).したがって,本研究で は個体数推定誤差を直接パラメータの推定に用い ず,感度分析の結果を見ながら(Table 2; Fig. 4), 絶滅リスク評価の頑強性や相対変化を確認すること とした(Staples et al., 2009).また個体数推定の系 統的な過小評価が存在する場合(推定値の 1.5 あ るいは 2 倍が実際に存在する場合)にも,絶滅確 率は極端に下がることはなく,さらに支流に遡上し ない個体の割合も絶滅確率に大きな影響を与える ものではなかった(パラメータ u;Table 3). 感度分析の結果,短いデータ年数に由来するい くつかのパラメータの推定誤差が,絶滅確率推定 に大きな影響を与えることが示された(Table 2). このことは,今回の結果が現状で最善のものとはい Fig. 4-3
え,絶滅確率をそのまま絶対値としてとらえるべき ではないことへの注意を促すものである.今後もモ ニタリング調査を継続し,逐次更新された最新の データで絶滅確率の評価を行っていく必要がある. アユモドキ個体群の現状評価 アユモドキは琵 琶湖・淀川水系と山陽地方にのみ自然分布する が,琵琶湖・淀川水系において,近年生息が確認 されていた京都府南丹市(旧八木町)周辺の桂川 水系では 1980 年代に,淀川ワンド群では 1990 年 代に,また琵琶湖周辺でも 1990 年代に姿を消し, 本調査地である京都府亀岡市周辺の桂川水系が現 在唯一確実な生息地点である(岩田,2006a;藤 田ほか,2008;上原,2013).この個体群において も,繁殖は農業用可動堰の起立による河川内の氾 濫原様環境の形成に負っており,堰下流部に取り 残された個体の上流部への移動,そして初期生息 場所として利用される農業用水路の水位低下時の 救出・移動など,地域住民や市民団体,行政の人 為的な補助をともないながら存続している現状にあ る(阿部・岩田,2007). 2006–2013 年の 8 年間で, 個体群サイズは平均 約 1,000 個体(うち高齢魚, 平均約 400 個体, 調 和平均約 270 個体)であり,ほぼ 10 倍幅で変動し ていた.さらに初期成長を終えた 9 月時点で,当 歳魚は 0 から 2,000 個体以上まで大きな変動を示 し,ときおりの高い繁殖率による突出年級群が本 個体群を支えているようである.このような傾向は 岡山県の 2 地域個体群についても同様である(阿 部ほか,未発表データ).アユモドキの孕卵数は標 準体長 100 mm ほどの個体で 3,000–12,000 粒とされ (宮地ほか,1976), 繁殖親魚数の変動とともに, 基本的に初期減耗が大きいことが推察される.特 に近隣のため池からのオオクチバスの侵入による当 歳魚の食害(2008 年)や集中豪雨にともなう可動 堰の緊急転倒(2012 年)などは,当歳魚の加入に 大きな影響を与えてきたと考えられる.本個体群の 現状を考えると,今後もこのような非恒常的な事象 による大きな影響が予想され,さらに生息地とその 周辺での環境改変が行われる場合には未知の変動 要素がこれに加わることになる. 今回行われた複数の個体数変動モデルや条件に 基づくPVA から一貫して得られた予測として,本個 体群は,現状において,現実的な絶滅リスクに直 面していると考えられる.密度効果を含む個体数 ベースモデル(A-2)で上限を 5,000 個体とした場 合に,例外的に低い絶滅確率が得られたが,それ 以外のすべてのモデルと条件で,50 年間で 23–82% の高い絶滅確率が予測された.モデル A-2 では低 密度下において大きい増加率を示し,さらに上限 値を低く設定した場合には極端に大きい減少率を もたらす高密度状態が計算上排除されることによっ て,絶滅が起こりにくい状況が生じたものと考えら れる.しかし,この A-2 においても,瞬間増加率 の減少やばらつきのわずかな増大で高い絶滅確率 がもたらされた.PVA における絶滅確率は,一般 に必ずしも将来予測の絶対値として解釈することは で き な い( 例 え ば,Beissinger and MacCullough, 2002; Frankham et al., 2002).また前述のように今回 の PVA は,もとになるデータ年数が短いことや齢 構造モデルが単純で,特に高齢魚の生残パターン や寿命が不明であることなど,不十分な点が含ま れる.しかし,現状での最良の推察として,本個 体群が脆弱な状況にあることを前提とし,今後の生 息環境の維持や改善を通して,本個体群の存続に 最善の努力を払うべきである. PVA に基づく保全方策 各モデルのパラメータ 空間上の絶滅確率分布から,本個体群の有効な保 全方策の方向性について検討が可能である.まず 齢構造を考慮しない個体数ベースモデル(A)に おけるパラメータは瞬間増加率に関わるもののみで あり,定性的に予測されるとおり,絶滅確率を下げ るためには,瞬間増加率を上げ,またその変動を 抑える必要がある(Fig. 4A, B).ここで瞬間増加率 (r)はある年の 9 月から次年の 9 月の総個体数の 比に基づくものであり,冬季の生残率,成熟個体 の繁殖場所への遡上率,そして繁殖成功率とその 後 9 月までの仔稚魚の初期生残率が複合した値で ある.観察された瞬間増加率のばらつきの大きさか ら,潜在的に絶滅確率を下げることは可能だと考え られるが(Fig. 4A),そのためには調査期間中に実 際に見られたような極端な個体数の減少が抑えられ る必要がある.またモデル A-2 が示すように(Fig. 4B),環境収容力が減少した場合には顕著な絶滅 確率の上昇が予測されることに加え,環境収容力 がたとえ大きく増加した場合でも,個体数の変動性 が現状よりも増すと大幅な絶滅確率の増加がもたら される.以上の一貫した結果は,現在保全活動と して進められているオオクチバスの流入防止の徹底 や駆除,また極端な堰堤操作の見直しなどが,ア ユモドキの個体数の大幅な減少を回避することで 個体群を安定化し,今後,個体群存続性の上昇に 対して効果を表すことを期待させる.一方,今後生 息地とその周辺での環境改変が遡上率,また遡上 後の繁殖率や生残率の変動性を高めた場合には,
その深刻な悪影響が懸念される. 齢構造を考慮したモデル B では,繁殖率と生残 率を分離して検討できる.ここで繁殖率(r′)は繁 殖成功率と 9 月までの初期生残率の複合値であり, 生残率(S0,S1)は冬季の生残率と成熟魚の遡上 率に関係する.モデル B-1 において特徴的な点は, まず繁殖率については推定値の周辺に比較的広く 高 い 絶 滅 確 率を 示 す 領 域 が 広 がっており(Fig. 4C),モデル A と同様,低い絶滅確率の達成のた めには観察値のなかでも高い水準の値が必要とな ることである.つまり,繁殖率の上昇による絶滅確 率の低下は,潜在的には可能であるが,現状では 難易度が高いと推察される.これは,モデル B-2 において,環境収容力に関わる KBの広い範囲で 高い絶滅確率が維持されることによっても支持され る(Fig. 4D).500 個体以上の新規加入が時々見ら れる現状を考慮すると,繁殖率の改善のためには, 実現性が未知で予測性の低い繁殖・初期生息場所 の大規模な改善というより,現状の環境において適 切な人為的介助を維持することが有効で確実性の 高い方策であり,上述のとおり,極端に繁殖や初 期生残が悪い年が生じないための継続的な保全対 策が重要であろう. 一方,モデル B-1,B-2 において,生残率のパラ メータ(S0)は急峻な変化を示す狭い領域に位置 しており, 絶滅確率への感受性が高かった(Fig. 4C, D).このことは,保全策において,冬季の生残 率や成熟魚の遡上率の改善が絶滅リスクを下げる うえで効果が大きいことを示唆する.しかし,それ らを実現するためにはいくつかの不明な点がある. 本種の越冬場所に関する知見は少ないが,本生息 地では支流との合流部に近い桂川の自然または人 工の複雑な構造を示す岸辺で観察された例がある (京都府南丹土木事務所,未発表データ).そのよ うな場所を十分に確保することにより,繁殖遡上群 の個体群サイズを最大にし,変動を抑えることが重 要であろう.またできるだけ多くの成熟魚を繁殖場 所に遡上させるためには,遡上時,可動堰の下に 集まる際に現実的なリスクとして存在する鳥類やナ マズ Silurus asotus などによる過度の捕食を抑えるこ とが必要である(特定非営利活動法人亀岡 人と 自然のネットワーク,2011;岩田ほか,未発表デー タ).それとともに,可動堰の起立による遡上阻害 を改善する必要がある.しかし,可動堰の起立は, 現在アユモドキの繁殖・初期生育環境を形成する 重要な水文要因でもある.現在,遡上が妨げられ た個体を人為的にもち上げる作業が地域で行われ ており,このような継続的な保全活動を今後も継続 することが有効であろう.このような遡上阻害と繁 殖・初期生育環境の形成の間のジレンマは,岡山 県吉井川水系の生息地でも同様にみられ(阿部・ 岩田,2007;阿部,2013),有効な魚道をともなう 可動堰への改修などが今後検討されるべきである. 2006–2013 年の 8 年間で, 本生息地におけるア ユモドキ高齢魚の個体数の調和平均は約 270 個体 であり,これは有効集団サイズの上限推定値とみ なすことができるだろう.マイクロサテライト DNA に基づいて推定された本個体群の当歳魚に対する 有効(親魚)集団サイズは約 110 個体(5 年間の 調和平均)と推定され(Watanabe et al., 未発表デー タ),上記の生息個体数と整合的である.長期的な 存続と進化可能性の保持のために,しばしば野生 生 物の有 効 集 団サイズは 500 以 上 が 必 要とされ (例えば,Soulé, 1980; Lehmkuhl, 1984),実際の個 体数はこれより大幅に大きい必要がある(例えば, Frankham et al., 2002).本個体群の集団サイズはそ のような基準に大きく足りない. 本研究における PVA は,遺伝的多様性や進化可能性の維持に対し て十分な評価が行われていない.本個体群の有効 集団サイズを増加させ,この近畿地方に唯一残る 貴重なアユモドキ個体群の将来にわたる存続を保 証するためには,まず,上述のとおり,現在の生息 場所の保全を通じた個体群の安定的な維持を図る ことが必要である.一方で,局所個体群における 大きな年変動は氾濫原を利用する生物の特性であ るとも考えられる.したがって,その長期的な存続 を目指すためには,近隣の潜在的な生息環境を改 善し,生息・繁殖域を拡大することにより,環境変 動に頑強なメタ個体群を再生していくことが重要で ある. 謝 辞 本研究で利用した生息個体数データは,地域住 民,とりわけ地元自治会と特定非営利活動法人亀 岡 人と自然のネットワークのメンバー,そして亀 岡市, 京都府, 環境省等の職員の尽力, 協力に よって行われた調査により得られたものである.ま た 2 名の匿名査読者には解析と原稿の改善に役立 つ貴重なご意見をいただいた.記して厚くお礼申し 上げる.本研究の公表には環境省近畿地方環境事 務所の協力を得た.
引 用 文 献 阿部 司.2013.アユモドキ(Parabotia curta)を保 全することの難しさ―岡山県における保全活動とそ の課題解決に向けて―. 地域自然史と保全,35: 23–32. 阿 部 司・ 岩 田 明 久.2007.アユモドキ: 存 続の カギ を 握る繁 殖 場 所 の 保 全. 魚 類 学 雑 誌,54: 234–238.
Abe, T., I. Kobayashi, M. Kon and T. Sakamoto. 2007a. Spawning behavior of kissing loach (Leptobotia curta) in temporary waters. Zool. Sci., 24: 850–853.
Abe, T., I. Kobayashi, M. Kon and T. Sakamoto. 2007b. Spawning of kissing loach (Leptobotia curta) is limited after the formation of temporary waters. Zool. Sci., 24: 922–926.
Akçakaya, R., M. A. Burgman and L. R. Ginzburg. 1999. Applied population ecology: principles and computer exercises using RAMAS® EcoLab. 2nd edn. Sinauer,
Sunderland, MA. 285 pp.
Beissinger, S. R. and D. R. McCullough. 2002. Population viability analysis. The University of Chicago, Chicago. 577 pp.
Frankham, R., J. D. Ballou and D. A. Briscoe. 2002. Introduction to conservation genetics. Cambridge University Press, Cambridge. 617 pp.
藤田朝彦・西野麻知子・細谷和海.2008.魚類標本 から見た琵琶湖内湖の原風景.魚類学雑誌,55: 77–93. 伊 藤 嘉 昭・ 村 井 実.1977. 動 物 生 態 学 研 究 法, 上巻.古今書院,東京.268 pp. 岩田明久.2006a.アユモドキの生存条件について 水田農業の持つ意味.保全生態学研究,11: 133– 141. 岩田明久.2006b.平成 17 年度アユモドキ調査結果 報告書.未公刊資料(亀岡市,京都府,環境省, 文化庁). 岩田明久.2007.平成 18 年度アユモドキ調査結果 報告書.未公刊資料(亀岡市,京都府,環境省, 文化庁). 岩田明久.2009.亀岡におけるアユモドキの生態と その現状.亀岡文化資料館(編),pp. 13–17.保 津川のサカナのハンドブック―第 24 回特別展・国 の天然記念物アユモドキと保津川水系のサカナた ち―」.亀岡文化資料館,京都. 片野 修.1997.アユモドキ.長田芳和・細谷和海 (編),pp. 95–103.日本の希少淡水魚の現状と系 統保存.緑書房,東京.
Lehmkuhl, J. F. 1984. Determining size and dispersion
of minimum viable populations for land management planning and species conservation. Environ. Manag., 8: 167–176.
Longergan, M and J. Harwood. 2003. The potential effects of repeated outbreaks of phocine distemper among harbor seals: a response to Harding et al. (2002). Ecol. Lett., 6: 889–893. 亀岡市.2012.大規模スポーツ施設∼京都府の専用 球 技 場 ∼ 亀 岡 市 に決 定.http://www.city.kameoka. kyoto.jp/suisin/kurashi/kyoiku/leisure/sports-jigyo/ sutajiamu-kettei.html.(参照 2013-12-18). 前 畑 政 善.2003. アユ モドキ. 環 境 省( 編 ),pp. 48–49.改訂 ・ 日本の絶滅のおそれのある野生生 物−レッドデータブック− 4.汽水 ・ 淡水魚類.自 然環境研究センター,東京.
Manly, B. F. J. 2006. Randomization, bootstrap and Monte Carlo methods in biology, 3rd edition. Chapman & Hall. 480 pp.
宮地傳三郎・川那部浩哉・水野信彦.1976.全改 訂新版 原色日本淡水魚類図鑑.保育社,大阪. 462 pp.
Morris, W. F. and D. F. Doak. 2002. Quantitative conservation biology: theory and practice of population viability analysis. Sinauer, Sunderland, MA.
日本魚類学会.2013.京都府亀岡市のアユモドキ生 息地における大規模開発に関する意見.http://www. fish-isj.jp/iin/nature/teian/130528.html.(参照 2013-12-18).
Sæther, B.-E. and S. Engen. 2002. Including uncertainties in population viability analysis using population prediction intervals. Pages 191–212 in S. R. Beissinger and D. R. McCullough, eds. Population viability analysis. The University of Chicago, Chicago.
Soulé, M. E. 1980. Thresholds for survival: maintaining fitness and evolutionary potential. Pages 151–169 in M. E. Soulé and B. A. Wilcox, eds. Conservation biology: an evolutionary-ecological perspective. Sinauer, Sunderland, MA.
Staples, D. F., M. L. Taper, B. Dennis and R. J. Boik. 2009. Effects of sampling error and temporal correlations in population growth on process variance estimators. Environ. Ecol. Stat., 16: 547–560.
特定非営利活動法人亀岡 人と自然のネットワーク. 2011.平成 22 年度アユモドキ保護増殖調査事業 報告書,未公刊資料(亀岡市,京都府,環境省, 文化庁). 上原一彦.2013.琵琶湖淀川水系のアユモドキの現 状と保全.地域自然史と保全,35: 17–22.