1
学
位
論
文
Doctoral Thesis
論文題名 : 脊椎発生における脊索と神経管底板の役割に関する解析
(Examination of the role of the notochord and floor plate during the spinal development) 著 者 名 : 安藤 卓 Takashi Ando 指導教員名:熊本大学大学院医学教育部博士課程臨床医科学専攻運動骨格病態学分野 水田 博志 教授 審査委員名: 脳発生学担当教授 嶋村 健児 神経分化学担当教授 田中 英明 脳神経外科学担当教授 倉津 純一 形態構築学分野学担当教授 福田 孝一 2010年3月
2
目次
1) 要旨・・・p. 4
2) 発表論文リスト・・・p. 7
3) 謝辞・・・p. 8
4) 略語一覧・・・p. 9
5) 序論・・・p. 11
6) 材料と実験方法・・・p. 22
6-1)変異マウスの作製
6-2)骨格標本の作製
6-3)組織内における
-ガラクトシダーゼ(lacZ)活性の検出
6-4)DIG labeled RNA probe の作製
6-5)Whole mount in situ hybridization
6-6)組織標本の作製
6-7)統計学的処理
7) 結果・・・p. 29
7-1)Sd, Skt
Gt変異マウス後期胚における脊椎発生の解析
7-2)Sd-Skt
Gtダブル変異マウス胚の脊索における
-geo 遺伝子
発現検出による脊索発生の解析
3
7-3)Sd, Skt
Gt変異マウス胚における
Uncx, Paraxis 遺伝子を用いた
体節発生解析
7-4)Sd, Skt
Gt変異マウス胚における神経管底板発生の解析
7-5)Sd, Skt
Gt変異マウス胚における椎板発生の解析
7-6)Sd, Skt
Gt変異マウス胚における各組織のマーカー遺伝子発現
と形成された椎体数の比較
8) 考察・・・p. 36
9) 結語・・・p. 41
10)参考文献・・・p. 43
11)図表・・・p. 64
4
1) 要旨
【目的】脊索は脊椎発生の中心的役割を担当する。特に脊索は神経管より底板
を発生させるだけでなく、底板とともに脊椎形成の母床となる体節を分化させ
椎板(中軸骨格の直接的な原基)に誘導させることが知られている。しかし、「脊
椎発生にいつまで脊索が関与しているのか?」、「椎板発生に脊索と底板のど
ちらが主に作用しているのか?」という疑問に対して、これまで明確に説明し
ている文献は認めない。我々研究グループの先行実験で得られた、一度発生し
た脊索が早期に消失するものの、上位底板は発生し、下位脊椎の欠損を示す
Danforth’s short-tail (Sd) 変異マウス胚の表現型に注目した。本研究の目的は、Sd
変異ホモ接合体が示す「脊索は一度発生した後消失するが上位底板は誘導され
る」という表現型を利用する事で、脊椎発生における脊索と神経管底板の役割
を明らかにすることである。
【方法】Sd 変異のヘテロ接合体胚、ホモ接合体胚を見分けるため、Sd ゲノム変
異領域に隣接(0.95cM 下流)している Skt
Gt(我々の研究室でジーントラップ法によ
り得られた変異)をサテライトマーカーとして PCR 法による Sd 変異胚の
genotyping に使用した。また、Skt
Gtは脊索のレポーター遺伝子としても利用可能
であるため、X-gal 染色によって Sd 変異胚の脊索細胞の検出に使用した。解析
に使用する
wild type, +-+/+-Skt
Gt, Sd-+/+-+, Sd-Skt
Gt/+-+, Sd-Skt
Gt/+-Skt
Gt, Sd-+/Sd-+,
Sd-Skt
Gt/Sd-Skt
Gt胚を交配で作出した。骨格標本は胚生
18.5 日胚を用いて作製し
た。胚生 9.0, 10.0, 10.5 日胚の脊索、体節、神経管底板、椎板発生を、各種分子
マーカーを使用した in situ hybridization (ISH)法により検討した。
【結果】
Sd ホモ接合体は第 6 胸椎以下、Sd ヘテロ接合体は第 6 尾椎以下の中軸
骨格が欠失していた。
Sd ホモ接合体胚は E9.5 までに脊索が消失することがわか
5
カー
Pax1 を使用した ISH 法によって、ホモ接合体胚の底板における Shh 発現は、
第
17 体節レベル以下で消失し、椎板における Pax1 の発現は第 17 体節レベル以
下で消失していた。体節における
Uncx 発現には明らかな異常を認めなかった。
このことは、
Sd 変異胚の体節発生は正常であることを示している。また、いっ
たん誘導された底板は脊索消失後も
Shh を分泌し続けることがわかった。加え
て、底板発生領域(ホモ接合体: 第 6 胸椎レベルまで、Sd ヘテロ接合体: 第 6 尾
椎レベルまで)で椎板の誘導が起こっていることも明らかになった。その底板、
椎板発生領域は、脊索が消失する時期に発生した体節の領域と一致していた。
【考察】
Sd ホモ接合体胚でみられた下位脊椎欠損の原因は、体節から椎板への
分化異常に先行した脊索消失を起因とした底板の誘導不全にあることがわかっ
た。また、
Sd 変異胚で誘導された底板の領域・椎板の領域は、形成された中軸
骨格の椎体領域と一致していた。このことは、一度誘導された底板は単独で脊
椎発生が可能であることを強く示唆している。
【結論】本研究結果は、脊椎発生において脊索は、底板誘導時期まで関与して
おり、底板を誘導した後は主に脊椎発生の中心組織は底板になる事を示してい
る。
6
Abstract
Danforth’s short-tail (Sd) mouse is a semi-dominant mutation affecting the development of the
vertebral column. Although the notochord degenerates completely by embryonic day 9.5, the vertebral column exists up to the lumbar region, suggesting that the floor plate can substitute for notochord function. We previously established the mutant mouse line, SktGt, through gene trap
mutagenesis and identified the novel gene, Skt, which was mapped 0.95cM distal to the Sd locus. Taking advantage of the fact that monitoring notochordal development and genotyping of the Sd locus can be performed using the SktGt allele, we assessed the development of the vertebra,
notochord, somite, floor plate and sclerotome in +-+/+-SktGt, Sd-+/+-+, Sd-SktGt/+-+, Sd-SktGt/+-SktGt, Sd-+/Sd-+ and Sd-SktGt/Sd-SktGt embryos. In Sd homozygous mutants with a
C57BL/6 genetic background, the vertebral column was truncated in the 6th thoracic vertebra,
which was more severe than previously reported. The floor plate and sclerotome developed to the level of somite before notochord degeneration and the number of remaining vertebrae corresponded well with the level of development of the floor plate and sclerotome. Defects to the sclerotome and subsequent vertebral development were not due to failure of somitogenesis. Taken together, these results suggest that the notochord induced floor plate development before degeneration, and that the remaining floor plate is sufficient for maintenance of differentiation of the somite into the sclerotome and vertebra in the absence of the notochord.
7
2) 発表論文リスト
参考論文
①関連論文
1.Takashi Ando,
Kei Semba, Hiroko Suda, Akira Sei,
Hiroshi Mizuta,
Masatake Araki,
Kuniya Abe, Kenji Imai, Naomi Nakagata, Kimi Araki, and Ken-ichi Yamamura
The floor plate is sufficient for development of the sclerotome and spine without the
notochord.
Mechanisms of Development 2010, Article in Press
②その他の論文
1.Hiroko Suda, Lee, Kwang Jong, Kei Semba, Fumie Kyushima, Takashi Ando,
Masatake Araki, Kimi Araki, Yukihiro Inomata, Ken-ichi Yamamura
The Skt gene, required for anorectal development, is a candidate for a molecular
marker of the cloacal plate
8
3) 謝辞
熊本大学生命資源研究・支援センター・表現型クリニック分野において熊
本大学生命資源研究・支援センター・表現型クリニック分野・山村研一教
授と熊本大学大学院医学研究科運動骨格病態学分野・水田博志教授の御指
導の下、本研究を行いました。多くの御指導を頂き、深く感謝いたします。
また、熊本大学生命資源研究・支援センター・表現型クリニック分野・荒
木喜美准教授、熊本大学生命資源研究・支援センター・バイオ情報分野・
荒木正健准教授、熊本大学大学院医学研究科運動骨格病態学分野・仙波圭
先生には、日々の実験研究から論文作成まで幅広い御助言を頂きました。
また、表現型クリニック分野、運動骨格病態学分野の皆様には有形無形の
多くの御協力、ご支援を受けました。
心より感謝いたします。
9
4) 略語一覧
bHLH:basic helix-loop-helix
-gal:
-galactosidase
-geo:
-galactosidase と neomycin 耐性遺伝子の融合タンパクをコードした遺伝子BMP:bone morphogenetic protein
cM:centimorgan, センチモルガン
DNA:Deoxyribo nucleic acid, デオキシリボ核酸
E:Embryonic day, 胚生
EMT:Epithelial-to-Mesenchymal Transition, 上皮間葉移行
ES 細胞:Embryonic stem cell, 胚性幹細胞
HE :Hematoxylin-Eosin
IRES:internal ribosomal entry site
MCS:マルチクローニングサイト
MET:Mesenchymal-to-Epithelial Transition, 間葉上皮移行
MGI:mouse genome informatics
ns:not significant
Pax1:Paired-box gene 1
10
PCR:polymerase chain reaction, ポリメラーゼ連鎖反応
RT:room temperature, 室温
SD:Standard Deviation, 標準偏差
Sd:Danforth’s short tail マウス
Skt:Sickle tail 遺伝子
SktGt:B6;CB-SktGtAyu8021IMEG
Shh:Sonic hedgehog
11
5) 序論
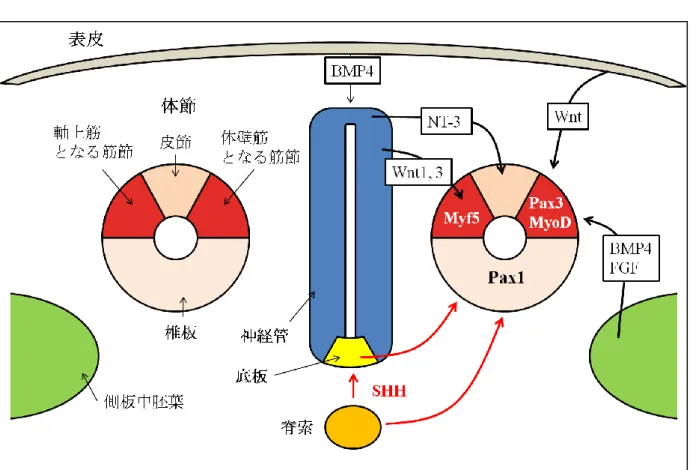
脊椎動物の体幹の中軸をなす骨格である脊柱は堅牢性、安定性、弾性そして可
動性を併せ持つ組織である。これらの特性は高等脊椎動物においては椎骨と椎
間板の繰り返し構造によりもたらされる。一方、下等脊椎動物は椎間板を持た
ず、椎骨は直接連結し脊柱を形成する(Romer and Parsons, 1986)。動物種による
これらの差異があるにも関わらず、椎体の発生過程は種間で良く保存されてい
る。脊柱を構成する要素のおおもとは傍軸中胚葉から派生する体節である。傍
軸中胚葉は原腸形成の間、脊索と神経外胚葉に挟まれる形で存在し、その後、
間葉上皮移行(mesenchymal- epithelial transition; MET)による上皮化、分割を経て
上皮性体節となる。体節はその背腹軸上の位置によって周囲組織から受けるシ
グナルが異なる。最背側は神経管より neurotrophin-3(NT-3)のシグナルを受け皮
節(dermatome)へ、中間は主に Wnt シグナルにより筋節(myotome)へ、そして最腹
側部は上皮間葉移行(Epithelial-Mesenchymal Transition; EMT)を経て脊椎、軟骨、
靭帯、さらには頭蓋底の一部などの前駆組織である椎板(sclerotome)へと分化す
る(Fig.1) (Gilbert, 2000)。椎板は疎性間葉組織であるが、まず外側部が凝集し椎
弓が形成され、後に内側部が凝集することで椎体、椎間板が形成される。上述
12
可欠な過程である(Gossler and Tam, 2002; Humzah.MD and Soames.R.W., 1988;
Langman.J, 1969; Theiler, 1988)。
体節を椎板へと分化誘導する際、中心的な役割を担う組織は脊索と神経管底板
である。脊索は原始結節(node)を形成する間葉細胞が、頭方に移動して、脊索突
起 notochordal process を形成するところよりその発生が始まる。脊索突起の中
を原始窩からの窪みが進入することにより、充実性から中腔性になり脊索管
(notochordal canal)となる。脊索管は内胚葉と融合し脊索板を形成し、組織の巻き
込みによって再び内胚葉より分離し脊索となる。次に脊索の誘導により外胚葉
が肥厚し、神経板を生ずる。神経板がしだいに原始線条のほうに伸長するに伴
い、側方縁が持ち上がって神経ヒダと呼ばれる構造になり、また押し下げられ
た中央域は神経溝となる。神経ヒダは徐々に正中線で接近し、やがて癒合して
管状の構造、すなわち神経管を形成する。神経管が形成されると引き続き脊索
からのシグナルを受ける最腹側部に神経管底板が誘導される。
これまで、脊索や神経管底板で発現する遺伝子について種々の報告がなされて
きた。Table.1, 2 に MGI (mouse genome informatics, http://www.informatics.jax.org/)
に登録されている過去の報告により脊索・神経管底板で発現が検出された遺伝
子の一覧を示す。表のようにさまざまな遺伝子発現が確認されているが、脊索
13
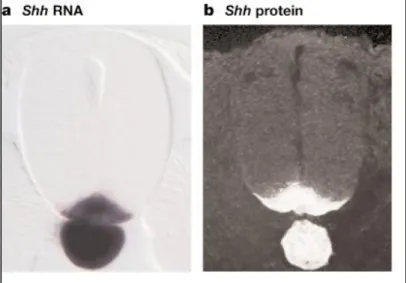
ものである。これまで明らかになっている神経管底板の役割は主にモルフォゲ
ンである Sonic hedgehog (Shh)分子を発現し神経管の背腹側軸における濃度勾配
を作ることで各レベルの細胞を腹側から順に運動神経、介在神経、感覚神経へ
と分化させるというものである(Jessell, 2000)。具体的なメカニズムとしては作ら
れた濃度勾配により Shh により発現が抑制される転写因子(class I)はその閾値が
低いほど背側の領域で発現し、発現が促進される転写因子(class II)はその閾値が
低いほど背側まで発現領域が拡大する。発現した class I、class II タンパクもお互
いを抑制しあい細かな発現のプロファイルが完成し、その領域の細胞をそれぞ
れの分化へ誘導する(Fig. 2)。
一方、体節から脊椎の直接の前駆組織である椎板の分化における脊索の役割に
ついては主にニワトリ胚における移植実験などにより明らかにされてきた。移
植された異所性の脊索のグラフトが接触する神経管に異所性の神経管底板を誘
導し、さらに周囲の中胚葉を軟骨へと分化させ、同時に皮膚や筋肉への分化を
抑制することが報告されている(Pourquie et al., 1993; Yamada et al., 1991)。しかし、
神経管底板の単独組織による体節から椎板への分化誘導に関する解析はいまだ
14
Table.1 脊索で発現が確認されている遺伝子
最初の報告者・文献 Acvr activin A receptor, type 1 Yoshikawa SI, Mech Dev 2000 Mar 1;91(1-2):439-44
Aebp1 AE binding protein 1 Ith B, Gene Expr Patterns 2005 Apr;5(4):533-7
Angptl1 angiopoietin-like 1 Lai DM, Dev Dyn 2007 Sep;236(9):2643-52
Anxa4 annexin A4 Hemre KM, J Comp Neurol 1996 May 13;368(4):527-37
Arhgef16 Rho guanine nucleotide exchange factor (GEF) 16 Sousa-Nunes R, Genome Res 2003 Dec;13(12):2609-20
Arsi arylsulfatase i Ratzka A, Dev Dyn 2010 Jun;239(6):1779-88
Bcor BCL6 interacting corepressor Wamstad JA, Gene Expr Patterns 2007 Apr;7(5):550-7
Bmp5 bone morphogenetic protein 5 Furuta Y, Development 1997 Jun;124(11):2203-12
Bmp7 bone morphogenetic protein 7 Furuta Y, Development 1997 Jun;124(11):2203-12
Cd44 CD44 antigen Wheatley SC, Development 1993 Oct;119(2):295-306
Cdh2 cadherin 2 Luo Y, Development 2001 Feb;128(4):459-69
Cdx2 caudal type homeobox 2 Beck F, Dev Dyn 1995 Nov;204(3):219-27
Cer1 cerberus 1 homolog (Xenopus laevis) Belo JA, Mech Dev 1997 Nov;68(1-2):45-57
Cfc1 cripto, FRL-1, cryptic family 1 Shen MM, Development 1997 Jan;124(2):429-42
Chrd chordin Goldman DC, Development 2000 May;127(10):2113-23
Cobl cordon-bleu Gasca S, Dev Genet 1995;17(2):141-54 (Gasca et al., 1995)
Col2a collagen, type II, alpha 1 Wood A, Development 1991 Apr;111(4):955-68
Crim1 cysteine rich transmembrane BMP regulator 1 Kolle G, Mech Dev 2000 Feb;90(2):181-93
Cthrc1 collagen triple helix repeat containing 1 Durmus T , Gene Expr Patterns 2006 Dec;6(8):935-40
D230030E09Rik Riken cDNA D230030E09 gene Hassan AS, Gene Expr Patterns 2010 Feb 12;10(2-3):127-34
Daam2 dishevelled associated activator of morphogenesis 2 Nakaya MA, Gene Expr Patterns 2004 Nov;5(1):97-105
Ddx3x DEAD/H (Asp-Glu-Ala-Asp/His) box polypeptide 3, X-linked Sowden JC, Mamm Genome 1997 Jan;8(1):42-4
Dlg1 discs, large homolog 1 (Drosophila) Caruana G, Mol Cell Biol 2001 Mar;21(5):1475-83
Fam198b family with sequence similarity 198, member B Meszaros R, Int J Dev Biol 2008;52(8):1119-22
Fn1 fibronectin 1 George EL, Development 1993 Dec;119(4):1079-91
Foxa1 forkhead box A1 Sasaki H, Development 1993 May;118(1):47-59
Foxa2 forkhead box A2 Sasaki H, Development 1993 May;118(1):47-59
Foxc1 forkhead box C1 Sasaki H, Development 1993 May;118(1):47-59
Furin furin (paired basic amino acid cleaving enzyme) Roebroek AJ, Development 1998 Dec;125(24):4863-76
Fut4 fucosyltransferase 4 Fox N, Dev Biol 1981 Apr 30;83(2):391-8
Fyn Fyn proto-oncogene Yagi T , Oncogene 1994 Sep;9(9):2433-40
Gad1 glutamic acid decarboxylase 1 Maddox DM, BMC Dev Biol 2001;1(1):1
Gal galanin T amplin OJ, BMC Genomics 2008;9(1):511
Gdf1 growth differentiation factor 1 Wall NA, Dev Biol 2000 Nov 15;227(2):495-509
Gli2 GLI-Kruppel family member GLI2 Ding Q, Development 1998 Jul;125(14):2533-43
Golph3 golgi phosphoprotein 3 Sowden JC, Mamm Genome 1997 Jan;8(1):42-4
Gpr98 G protein-coupled receptor 98 McMillan DR, J Biol Chem 2002 Jan 4;277(1):785-92
Gtf2ird1 general transcription factor II I repeat domain-containing 1 Bayarsaihan D, Gene Expr Patterns 2003 Oct;3(5):579-89
Hoxb4 homeobox B4 Brend T , Development 2003 Jun;130(12):2717-28
Igdcc4 immunoglobulin superfamily, DCC subclass, member 4 Salbaum JM, Genomics 2000 Feb 15;64(1):15-23
Ippk inositol 1,3,4,5,6-pentakisphosphate 2-kinase Verbsky J, P roc Natl Acad Sci U S A 2005 Jun 14;102(24):8448-53
Itga3 integrin alpha 3 T amplin OJ, BMC Genomics 2008;9(1):511
Lgals3 lectin, galactose binding, soluble 3 Fowlis D, Dev Dyn 1995 Jun;203(2):241-51
Lhx1 LIM homeobox protein 1 Barnes JD, Dev Biol 1994 Jan;161(1):168-78
Lyve1 lymphatic vessel endothelial hyaluronan receptor 1 Gittenberger-De Groot AC, Dev Dyn 2004 Jun;230(2):378-84
Map3k5 mitogen-activated protein kinase kinase kinase 5 Ferrer-Vaquer A, Gene Expr Patterns 2007 Aug;7(7):808-16
Mtrr 5-methyltetrahydrofolate-homocysteine methyltransferase reductase Elmore CL, Mol Genet Metab 2007 May;91(1):85-97
Ncam1 neural cell adhesion molecule 1 Moase CE, Development 1991 Nov;113(3):1049-58
Nck1 non-catalytic region of tyrosine kinase adaptor protein 1 Bladt F, Mol Cell Biol 2003 Jul;23(13):4586-97
Nck2 non-catalytic region of tyrosine kinase adaptor protein 2 Bladt F, Mol Cell Biol 2003 Jul;23(13):4586-97
Nes nestin Yaworsky PJ, Dev Biol 1999 JAN 15;205(2):309-321
Nodal Nodal Welsh IC, Dev Biol 2000 Sep 1;225(1):151-68
Nog noggin Goldman DC, Development 2000 May;127(10):2113-23
Notch2 Notch gene homolog 2 (Drosophila) Przemeck GK, Development 2003 Jan;130(1):3-13
Noto notochord homolog (Xenopus laevis) Plouhinec JL, Gene Expr Patterns 2004 Nov;5(1):11-22
Npr3 natriuretic peptide receptor 3 DiCicco-Bloom E, Dev Biol 2004 Jul 1;271(1):161-75
Nrg1 neuregulin 1 Meyer D, Development 1997 Sep;124(18):3575-86
Otx2 orthodenticle homolog 2 (Drosophila) Acampora D, Development 1995 Oct;121(10):3279-90
Pdgfc platelet-derived growth factor, C polypeptide Aase K, Mech Dev 2002 Jan;110(1-2):187-91
Ppp1r1a protein phosphatase 1, regulatory (inhibitor) subunit 1A T amplin OJ, BMC Genomics 2008;9(1):511
Prmt1 protein arginine N-methyltransferase 1 Pawlak MR, Mol Cell Biol 2000 Jul;20(13):4859-69
Prox1 prospero-related homeobox 1 Burke Z, Mech Dev 2002 Oct;118(1-2):147
Ptch1 patched homolog 1 Jeong Y, Development 2003 Aug;130(16):3891-902
Ptprd protein tyrosine phosphatase, receptor type, D T amplin OJ, BMC Genomics 2008;9(1):511
15
Table.1 脊索で発現が確認されている遺伝子の続き
最初の報告者・文献 Nodal Nodal Welsh IC, Dev Biol 2000 Sep 1;225(1):151-68
Nog noggin Goldman DC, Development 2000 May;127(10):2113-23
Notch2 Notch gene homolog 2 (Drosophila) Przemeck GK, Development 2003 Jan;130(1):3-13
Noto notochord homolog (Xenopus laevis) Plouhinec JL, Gene Expr Patterns 2004 Nov;5(1):11-22
Npr3 natriuretic peptide receptor 3 DiCicco-Bloom E, Dev Biol 2004 Jul 1;271(1):161-75
Nrg1 neuregulin 1 Meyer D, Development 1997 Sep;124(18):3575-86
Otx2 orthodenticle homolog 2 (Drosophila) Acampora D, Development 1995 Oct;121(10):3279-90
Pdgfc platelet-derived growth factor, C polypeptide Aase K, Mech Dev 2002 Jan;110(1-2):187-91
Ppp1r1a protein phosphatase 1, regulatory (inhibitor) subunit 1A T amplin OJ, BMC Genomics 2008;9(1):511
Prmt1 protein arginine N-methyltransferase 1 Pawlak MR, Mol Cell Biol 2000 Jul;20(13):4859-69
Prox1 prospero-related homeobox 1 Burke Z, Mech Dev 2002 Oct;118(1-2):147
Ptch1 patched homolog 1 Jeong Y, Development 2003 Aug;130(16):3891-902
Ptprd protein tyrosine phosphatase, receptor type, D T amplin OJ, BMC Genomics 2008;9(1):511
Rbp1 retinol binding protein 1, cellular Maden M, Development 1990 May;109(1):75-80
Ric8 resistance to inhibitors of cholinesterase 8 homolog (C. elegans) T onissoo T , Gene Expr Patterns 2003 Oct;3(5):591-4
Sall3 sal-like 3 (Drosophila) Ott T , Mech Dev 2001 Mar;101(1-2):203-7
Sdc1 syndecan 1 Sutherland AE, Development 1991 Sep;113(1):339-51
Sema5a sema domain, seven thrombospondin repeats (type 1 and type 1-like), transmembrane domain (TM) and short cytoplasmic domain, (semaphorin) 5A
Adams RH, Mech Dev 1996 Jun;57(1):33-45
Shh sonic hedgehog Echelard Y, Cell 1993 Dec 31;75(7):1417-30
Shisa2 shisa homolog 2 (Xenopus laevis) Filipe M, Dev Dyn 2006 Jun 13;235(9):2567-2573
Six1 sine oculis-related homeobox 1 homolog (Drosophila) Oliver G, Development 1995 Mar;121(3):693-705
Skp1a S-phase kinase-associated protein 1A Sowden JC, Mamm Genome 1997 Jan;8(1):42-4
Skt (Etl4) Sickle tail Semba K, Genetics 2006 Jan; 172: 445-456
Smad2 MAD homolog 2 (Drosophila) de Sousa Lopes SM, Gene Expr Patterns 2003 Jun;3(3):355-60
Smoc1 SPARC related modular calcium binding 1 T amplin OJ, BMC Genomics 2008;9(1):511
Sod1 superoxide dismutase 1, soluble Yon JM, J Mol Histol 2008;39(1):95-103
Sp5 trans-acting transcription factor 5 T reichel D, Mech Dev 2001 Mar;101(1-2):175-9
T T, brachyury Wilkinson DG, Nature 1990 Feb 15;343(6259):657-9
Tacc1 transforming, acidic coiled-coil containing protein 1 Lauffart B, Dev Dyn 2006 Jun;235(6):1638-47
Tcf12 transcription factor 12 Uittenbogaard M, Brain Res Gene Expr P atterns 2002 Jan;1(2):115-21
Tctn1 tectonic family member 1 Reiter JF, Genes Dev 2006 Jan 1;20(1):22-7
Tdgf1 teratocarcinoma-derived growth factor 1 Dono R, Development 1993 Aug;118(4):1157-68
Tes testis derived transcript Crompton LA, Dev Dyn 2007 Nov;236(11):3137-43
Trp53bp2 transformation related protein 53 binding protein 2 Sowden JC, Mamm Genome 1997 Jan;8(1):42-4
Use1 unconventional SNARE in the ER 1 homolog (S. cerevisiae) Duan JZ, Yi Chuan Xue Bao 2006 Aug;33(8):692-701
Vtn vitronectin Seiffert D, Dev Dyn 1995 May;203(1):71-9
Zic2 zinc finger protein of the cerebellum 2 Elms P, Gene Expr Patterns 2004 Sep;4(5):505-11
Zmiz1 zinc finger, MIZ-type containing 1 Rodriguez-Magadan H, Gene Expr Patterns 2008 Feb;8(3):206-13
Zmiz2 zinc finger, MIZ-type containing 2 Rodriguez-Magadan H, Gene Expr Patterns 2008 Feb;8(3):206-13
16
Table.2 神経管底板で発現が確認されている遺伝子
最初の報告者、文献 Alcam activated leukocyte cell adhesion molecule Fraboulet S, Mech Dev 2000 Jul;95(1-2):221-4
Anxa4 annexin A4 Hemre KM, J Comp Neurol 1996 May 13;368(4):527-37
Arx aristaless related homeobox Miura H, Mech Dev 1997 Jul;65(1-2):99-109
Bmp1 bone morphogenetic protein 1 Suzuki N, Development 1996 Nov;122(11):3587-95
Cacna1h calcium channel, voltage-dependent, T type, alpha 1H subunit Masuda T , Brain Res 2009 Jan 16;1249():61-7
Cd9 CD9 antigen Schmidt C, J Neurosci Res 1996 Jan 1;43(1):12-31
Cobl cordon-bleu Gasca S, Dev Genet 1995;17(2):141-54
Dbpht2 DNA binding protein with his-thr domain Miyamoto T , Dev Genes Evol 2003 May;213(4):199-202
Dst dystonin Dowling J, Dev Biol 1997 Jul 15;187(2):131-42
Enpp2 ectonucleotide pyrophosphatase/phosphodiesterase 2 Bachner D, Mech Dev 1999 Jun;84(1-2):121-5
Fam198b with sequence similarity 198, member B Meszaros R, Int J Dev Biol 2008;52(8):1119-22
Foxa1 forkhead box A2 Sasaki H, Development 1993 May;118(1):47-59
Foxa2 forkhead box A2 Sasaki H, Development 1993 May;118(1):47-59
Fstl5 follistatin-like 5 Masuda T , Brain Res 2009 Jan 16;1249():61-7
Gtf2ird1 general transcription factor II I repeat domain-containing 1Bayarsaihan D, Gene Expr Patterns 2003 Oct;3(5):579-89
Hspg2 perlecan (heparan sulfate proteoglycan 2) Joseph SJ, Development 1996 Nov;122(11):3443-52
Kitl kit ligand Wehrle-Haller B, Dev Biol 1999 Jun 1;210(1):71-86
L1cam L1 cell adhesion molecule Matise MP, Development 1999 Aug;126(16):3649-59
Mtss1l metastasis suppressor 1-like Saarikangas J, J Cell Sci 2008 May 1;121(Pt 9):1444-54
Nrcam neuron-glia-CAM-related cell adhesion molecule Matise MP, Development 1999 Aug;126(16):3649-59
Ntn1 netrin 1 Greene ND, Mech Dev 1998 Apr;73(1):59-72
Pdgfa platelet derived growth factor, alpha Orr-Urtreger A, Development 1992 Aug;115(4):1045-58
Ptch1 patched homolog 1 Goodrich LV, Genes Dev 1996 Feb 1;10(3):301-12
Rgma RGM domain family, member A Schmidtmer J, Gene Expr Patterns 2004 Jan;4(1):105-10
Rhob ras homolog gene family, member B Henderson DJ, Mech Dev 2000 Jul;95(1-2):211-4
Sall1 sal-like 1 (Drosophila) Ott T , Mech Dev 2001 Mar;101(1-2):203-7
Shh sonic hedgehog Echelard Y, Cell 1993 Dec 31;75(7):1417-30
Shroom3 shroom family member 3 Hildebrand JD, Cell 1999 Nov 24;99(5):485-97
Sorcs2 sortilin-related VPS10 domain containing receptor 2 Rezgaoui M, Mech Dev 2001 Feb;100(2):335-8
Sulf1 sulfatase 1 Lum DH, Mol Cell Biol 2007 Jan;27(2):678-88
Sulf2 sulfatase 2 Lum DH, Mol Cell Biol 2007 Jan;27(2):678-88
Trim9 tripartite motif-containing 9 Berti C, Mech Dev 2002 May;113(2):159-62
Vtn vitronectin Seiffert D, Dev Dyn 1995 May;203(1):71-9
17
この脊索・神経管底板による体節の椎板への分化を担っている分子もまた神経
発生において重要な役割を持っている Shh であることもその後の解析により明
らかになっている(Ang and Rossant, 1994; Chiang et al., 1996; Roelink et al., 1995;
Wilson et al., 1995)。Shh のノックアウトマウスはほとんどが胚生(Embryonic day;
E)12 日までに致死になるが、
まれに E18.0 まで発生するものも確認されている。
後期まで発生した胚は眼球、耳、外生殖器、手指・足趾骨、脊椎を完全に欠損
する非常に重篤な表現型を示す。これらの解析により脊索や誘導された神経管
底板より分泌された Shh シグナルが体節の椎板への分化、またそれに続く脊柱
の形成に必須であることが明らかになった(Chiang et al., 1996; Fan and
Tessier-Lavigne, 1994; Roelink et al., 1995; Teillet et al., 1998)。また、McMahon らは
ノックアウトマウスの研究により脊索由来の Noggin が椎板の分化に必要である
ことを示した。一方、bone morphogenetic protein (BMP)2 や BMP4 が Shh シグナ
ルにより誘導されるべき体節間葉における Pax1 発現を抑制し、椎板、ひいては
軟骨の分化を阻害していることも明らかになっている(McMahon et al., 1998;
Monsoro-Burq et al., 1996)。Noggin は BMP2 や BMP4 に強固に結合しそれらが受
容体に結合するのを妨げる(Zimmerman et al., 1996)。上述してきたように、脊索
や神経管底板からの Shh シグナルに反応して体節腹内側部が椎板へと分化する
18
Danforth’s short tail (Sd) マウスは短く曲がった尾椎、泌尿生殖器異常、肛門直腸
奇形を持つ自然発生の半優性変異体である(Dunn et al., 1940; Favre et al., 1999;
Gruneberg, 1958)。これまでの研究により Sd 変異がマウス 2 番染色体近位に
mapping されているにもかかわらず、責任遺伝子はいまだ同定されていない
(Alfred et al., 1997; Lane and Birkenmeier, 1993; Maatman et al., 1997)。Sd ホモ接合
体胚において、脊索はいったん形成されるものの E9.5-11.0 の間に消失してしま
うことが報告されている(Dietrich et al., 1993; Gruneberg, 1958)。Sd ホモ接合体後
期胚は腎臓の欠損、鎖肛、腰仙部での脊柱の途絶が表現型として報告されてお
り、また、脊椎欠損の程度は遺伝的背景に影響されることも明らかになってい
る(Dietrich et al., 1993; Dunn et al., 1940; Theiler, 1988)。Dietrich ら (1993) はまた、
Sd ホモ接合体後期胚が尾側の椎体や神経管底板を欠失している一方、頭側のそ
れらはほとんど問題なく形成されていることも示している。これらの知見によ
り、彼らは脊柱の発生において、神経管底板が誘導された後は脊索が存在しな
くても発生が進むのではないかと推測している。しかし、神経管底板が脊索の
機能を代替している十分な実験的知見を示す事に成功しているとは言えなかっ
た。その最大の理由は、Sd 責任遺伝子が明らかになっていないため genotyping
が不可能で、genotyping を外見から得られる表現型(最も早く外観より認めるホ
モ接合体の表現型の出現は E11.5)に依存するしか無く、外観上の表現型が現れる
19
までの胚発生に対する解析が困難だったためである。脊椎発生における神経管
底板の役割を研究するためには E10.5 以前の早期胚を用いた解析が必要である
が、E10.5 以前の Sd 変異胚は外見上の特徴をほとんど示さないことより研究を
進める上での大きな障壁となっていた。
我々の研究室では Cre-loxP システムを応用したトラップベクターを用いた
randomized mutagenesis を行っている。使用しているトラップベクターはスプラ
イスアクセプター、IRES 配列、lacZ と neomycin 耐性遺伝子の fusion タンパクで
ある
-geo、polyA シグナルを持っており、遺伝子のエクソン間に挿入されると
その部位より下流の遺伝子機能を破壊することができる(Araki et al., 1999)。我々
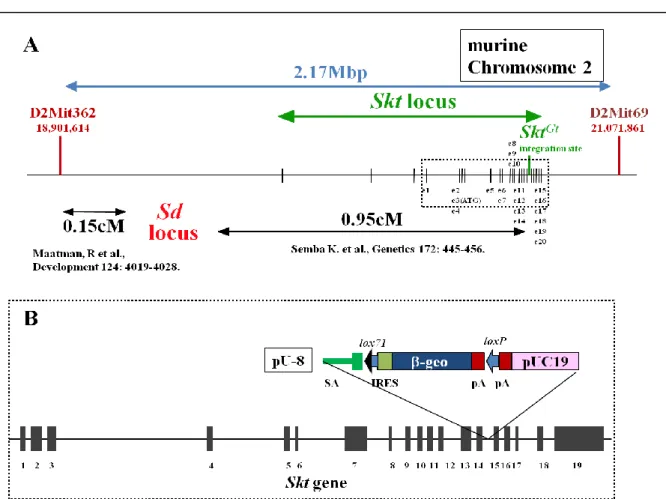
のグループでは過去、このトラップベクターpU-8 を ES 細胞に導入する gene trap
法により脊索でレポーターを発現する Skt
Gt変異マウスラインを作製し報告した
(Fig. 3 A and B, Fig. 5, Semba et al., 2006)。この際同定した Skt (Sickle tail)は新規遺
伝子であり、掛け合わせの結果により Sd 変異 locus の 0.95cM 遠位に位置してい
ることが明らかになった(Semba et al., 2006)。我々は Sd 変異マウスと Skt
Gt変異マ
ウスを掛け合わせることにより、2 つの変異が cis -configuration で存在するダブ
ル変異マウス(Sd Skt
Gt/+ +)を得ることができた。Skt locus に挿入された lacZ 遺伝
子は Sd 変異の genotyping マーカーとして有用であるだけでなく、脊索でのレポ
ーター発現解析により Sd 変異体胚の脊索変性もモニターすることができる。さ
20
らに Skt
Gtホモ接合体マウスは表現型として尾椎先端の椎間板形成異常による
屈曲変形を認める他に大きな異常を持たず、成体まで成長し妊孕性も持つ
(Semba et al., 2006)。以上より、Skt
Gtアレルは Sd 変異体胚を用いた神経管底板
による脊索の代替機能を研究するうえで非常に有用であると言える。
今回の研究ではまず C57BL/6 genetic background の Sd 変異体の脊椎数に対する影
響を検討した。次に種々の分子マーカーを用いて脊椎発生における脊索や神経
管底板の役割を解析した。分子マーカーとして脊索;Skt
Gt、体節;Uncx and
Paraxis、神経管底板;Shh、椎板;Pax1 をそれぞれ用いた(Burgess et al., 1995;
Furumoto et al., 1999; Mansouri et al., 1997; Roelink et al., 1995; Semba et al., 2006)。
Uncx は体節が前後極性の確立に関わる転写因子であり、体節の尾側半分で発現
する(Mansouri et al., 1997)。このことより体節マーカー、特に前後極性の分化マ
ーカーとして汎用されている。Paraxis は bHLH 型の転写因子であり、新たに形
成された上皮性体節で強く発現し、その後体節の分化が進むに従ってその発現
は減弱していくことが知られている(Burgess et al., 1995)。このことより特に新規
体節形成マーカーとして用いられている。
Pax1 は脊椎の前駆組織である椎板や
第三~第六咽頭弓の軟骨の形態形成に関わる転写因子である(Deutsch et al.,
1988; Furumoto et al., 1999)。体幹では椎板特異的に発現し椎板の分化マーカーと
して良く用いられている。
21
本研究では、Sd 変異マウス胚に対して行った前述の各種分子マーカーの発現解
析、骨格欠損レベルの解析を行うことにより、Sd 変異マウス胚が傍軸中胚葉の
体節形成能に異常をきたさない事、脊索が消失しても神経管底板が誘導されて
いれば、神経管底板単独で体節から椎板への分化、ひいては脊椎の形成誘導を
維持できることを示すことができた。
22
6) 材料と実験方法
6-1)変異マウスの作製
Danforth’s short tail mouse (Sd)マウスはJackson研究所より購入し、胚移植により
繁殖させ、C57BL/6バックグラウンドへ10回以上戻し交配したものを使用した。
Skt
Gt/+マウスは仙波らが表現型クリニック分野においてジーントラップ法を用
いて樹立したものを使用した(Araki et al., 1999; Semba et al., 2006)。Sd-Skt
Gtダブル
変異マウスはSd/+マウスと、Skt
Gt/+マウスを交配することにより作製した。Sdマ
ウスはその外観によりgenotypeを判断し、Skt
Gtはgenomic PCR法でgenotypingを行
った。wild-type alleleはSkt遺伝子の14番イントロンに設計したGTSプライマー(5’-
CCACCCCTACATGTGTCTTT -3’)とGTAプライマー(5’-
CGAGTAAGTAACATCCCTCC -3’)を使用し、339bpの産物を得ることで検出でき、
trapped alleleはZ1プライマー(5’- GCGTTACCCAACTTAATCG -3’)とZ2プライマ
ー(5’- TGTGAGCGAGTAACACAAG -3’)を使用し、320bpの産物を確認すること
で検出することができた。
6-2)骨格標本の作製
各genotype、18.5日胚の皮膚を剥離し、95%エタノールで3日間固定した。1%水
酸化カリウム溶液に浸漬し透明化した後にアルシアンブルー、アリザリンレッ
ド染色を行った。2%水酸化カリウム溶液で過剰な染色液を除去した後に、グリ
23
セロールに置換した(Hogan et al., 1994)。脊椎数をカウントする際、明らかな異
常の無い物を正常とし、カウントした(その際わずかな大きさの変化は考慮しな
かった)。
6-3)組織内における
-ガラクトシダーゼ(lacZ)活性の検出
Allenらの方法によりWhole mount X-gal 染色を行った(Allen et al., 1988)。サンプ
ルは固定液(1% formaldehyde, 0.2% glutaraldehyde, and 0.02% Tween-20 in
phosphate-buffered saline (PBST))中で室温にて30分固定し、PBSで30分間、2回洗
浄する。その後、X-gal染色液(5 mM potassium ferricyanide, 5 mM potassium
ferrocyanide, 2 mM MgCl
2, 0.5% (v/v) X-gal in PBS)に浸漬しovernight、30℃で染色
した。その後、PBSで5分間、2回洗浄し、4% paraformaldehyde in PBSで再固定し
た。観察の際はエタノールステップ(25%, 50%, 75%, 100% and 100% それぞれ
30分反応)で脱水し、benzylalcohol/benzylbenzoate (1:2)で透明化した。
6-4)DIG labeled RNA probe の作製
Shh, Pax1, Uncx の cDNA はすでに当教室(表現型クリニック分野)にクローニング
されたベクターが存在していた。
Paraxis については東海大学の阿部幸一郎先生
のご厚意により
cDNA がクローニングされたベクターを頂いた(Kokubu et al.,
2004)。それらベクターを大腸菌にて増幅させた後にプラスミドを QIAGEN
24
cDNA の 5’側の MCS の制限酵素サイトで digest した(37℃、ovenight)ベクターを、
RNA Polymerase (T3, T7 or Sp6 プロモーター)(Roche Diagnostics)、DIG RNA
Labeling Mix(Roche Diagnostics)とともに 37℃、2 時間 incubate して作製した。in
vitro transcription 後の RNA プローブの精製にはマイクロバイオスピンカラム 30
Tris RNase free(BioRad)を用いた。作製したプローブは 80℃、3 分間 denature し
た後に
fresh な 1×TBE バッファーで作製したアガロースゲルで 100V、15 分電
気泳動して目的のサイズが得られているかを確認した。
6-5)Whole mount in situ hybridization
Rosen らの方法を一部改変した Kokubu らの方法により Whole mount in situ
hybridization を行った(Kokubu et al., 2004; Rosen and Beddington, 1993)。
i) サンプルの剖出、固定、保存
PBS に浸漬した状態で on ice で解剖により採取した胚は固定液(4%
paraformaldehyde in PBS)中で 4℃、overnight 固定し、PBS containing 0.1% Tween-20
(PBST)で洗浄、メタノールステップ(25%, 50%, 75%, 100% and 100% それぞれ
30 分反応)にて脱水した後に-80℃で保存した。
ii) Hybridization
・MetOH 中で保存している胚を on ice で 75%、50%、25% MetOH/PBS で 10 分
25
・PBST で 10 分、2 回 wash (on ice)、液交換してさらに 5 分 wash (on ice)
・RIPA バッファー(0.05% SDS, 0.15M NaCl, 1% NP40, 0.5% デオキシコール酸,
1mM EDTA, 5mM Tris-HCl pH8.0) で 10 分震盪せずに incubate (on ice)した。
・大量の PBST で 5 分、震盪せずに 2 回 wash (on ice)した。
・4%PFA/0.2%グルタールアルデヒド/PBST できっちり 20 分間再固定した。
・PBST で 5 分、2 回 wash した。
・hybe-buffer (50% formamide, 5×SSC, 500µg heparin, 0.02% Tween20)/PBT (1:1 mix)
で 10 分間 incubate (RT)後、hybe-buffer で 10 分間 wash (RT)した。
・ hybe-buffer + tRNA (100µg/ml, invitrogen) で 68 ℃ 、 3 時 間 incubate し
prehybridization を行った。
・RNA プローブを 80℃, 3 分間 denature した後に、hybe-buffer + tRNA (100µg/ml)
に 1:100 になるように添加し(250ng/ml)、68℃、overnight で hybridization させた。
iii) 余剰プローブの除去、DIG 抗体反応
・翌日、プローブ溶液をピペットで注意深く取り除き、hybe-buffer で 30 分、65℃
で 2 回 wash した。
・hybe-buffer と RNase solution (0.5M NaCl, 10mM Tris-HCl pH7.5, 0.1% Tween20)
の 1:1 混合溶液で 5 分間 wash (RT)した。
26
37℃、60 分 incubate した。
・RNase solution と SSC/FA/Tween20 (50% formamide, 2×SSC, 0.1% Tween20) の
1:1 混合溶液で 5 分間 wash (RT)した。
・SSC/FA/Tween20 で 5 分/2 回、10 分/3 回、30 分/5 回 wash(65℃)した後に胚を
室温に戻す。
・SSC/FA/Tween20 と Tween-20 を含む Tris buffered saline (TBST; 25 mM Tris, 137
mM NaCl, 2.68 mM KCl, 0.1% Tween-20) の 1:1 混合溶液で 5 分間 wash (RT)した。
・TBST で 10 分、2 回 wash(RT)した後に溶液を MABT(100mM マレイン酸, 150mM
NaCl, pH7.5, 0.1% Tween-20)に置換しさらに 10 分、2 回 wash(RT)した。
・blocking buffer (MABT, 1.5% blocking reagent [Roche Diagnostics])で 1 時間
blocking RT)し、並行して DIG 抗体(Anti-Digoxigenin-AP, Fab fragments [Roche
Diagnostics]) を 別 に 用 意 し た blocking buffer に加え (1 : 2000) 、 4 ℃ , 1 時間
preabsorption した。
・blocking buffer を DIG 抗体添加バッファーに置換し、震盪しながら overnight
反応させた(4℃)。
iv) 余剰抗体の洗浄、発色
・まず TBST で 5 分、3 回 wash(RT)した後に 1 時間ごとに 8 回液交換し、wash (RT)
27
・TBST を交換し、震盪しながら 3 時間 wash (RT)した後に溶液を NTMT[100mM
NaCl, 100mM Tris-HCl (pH 9.5), 50mM MgCl2, 0.1% Tween20]に置換し 5 分、2 回
wash (RT)した。
・NBT/BCIP Stock Solution (Roche Diagnostics)/ NTMT 溶液を用いて発色させた(暗
所、4℃ or RT)。発色の程度は 5 分ごとに実体顕微鏡下でチェックした。
・発色させた胚は NTMT で 5 分/2 回、PBST で 10 分/2 回洗浄し、4%
paraformaldehyde /PBS で 30 分再固定した(RT)後に PBST 溶液内で保存した(4℃)。
・観察の際はエタノールステップ(25%, 50%, 75%, 100% and 100% それぞれ 30
分反応)で脱水し、benzylalcohol/benzylbenzoate (1:2)で透明化した。
6-6)組織標本の作製
組織標本は採取、または染色した胚を3.7% formaldehyde/PBSで再固定し、全自
動パラフィン置換装置 HISTOS5(Leica Microsystems)を用いてパラフィン置換を
行い、包埋した後に5
m(HE用)、10
m (X-gal染色、Whole mount in situ
hybridization後のサンプル)の厚さで薄切した。5
m の厚さのものは
Hematoxylin-Eosin (HE)で染色し、X-gal染色の対比染色にはNuclear Fast red を用
いた。
6-7)統計学的処理
28
Pax1 の発現レベル、そして椎体数を Student's t test で処理し、P < 0.05 を有意差
29
7) 結果
7-1)Sd, Skt
Gt変異マウス後期胚における脊椎発生の解析
我々はまず、
Sd 変異体における椎体欠失の程度、また Skt
Gtアレルが Sd 変異体
の椎体欠失に影響を与えるかどうかについて検討した。方法として、wild-type、
+-+/+-Skt
Gt、
Sd-+/+-+、Sd-Skt
Gt/+-+、Sd-Skt
Gt/+-Skt
Gt, Sd-+/Sd-、 Sd-Skt
Gt/Sd-Skt
Gtそ
れぞれの胚を
E18.5 で採取して骨格標本を作製し、椎体数を比較することで解析
を行った。
wild-type、+-+/+-Skt
Gt胚の椎体数は約
60 であり、内訳は頚椎;7 椎体、
胸椎;
13 椎体、腰椎;6 椎体、仙椎;4 椎体、尾椎;約 30 椎体であった(Fig.4A and
B)。Sd-+/+-+ (n=9)、 Sd-Skt
Gt/+-+ (n=16)、Sd-Skt
Gt/+-Skt
Gt(n=9) 変異胚は脊椎が
それぞれ第
6 尾椎、第 2 尾椎、第 4 仙椎で途絶していた(Fig. 4C-E)。また Sd-+/+-+
と
Sd-Skt
Gtダブル変異体(Sd-Skt
Gt/+-+、Sd-Skt
Gt/+-Skt
Gt)の椎体数の間には Student’s
t-test による解析の結果、統計学的有意差が認められた(P < 0.005, Fig. 4H)。一方
Sd-+
/Sd-+ (n=6)、Sd-Skt
Gt/Sd-Skt
Gt(n=4)変異胚では脊椎がそれぞれ第 7 胸椎、第 6
胸椎で途絶しており(Fig. 4F and G)、2 つの群に統計学的有意差は認めなかった
(Fig. 4H)。以上の結果により、Sd アレルが存在すると加算的に激しい椎体欠失
を引き起こすこと、また、
Skt
Gtアレル単独で存在しても椎体形成に影響はない
が、
Sd 変異体に Skt
Gtアレルが加わることで仙椎、尾椎の椎体形成に影響を及ぼ
すことが示された。一方、
Skt
Gtアレルは上位椎体の形成には全く影響しないこ
30
とが示唆される結果となった。
7-2)Sd-Skt
Gtダブル変異マウス胚の脊索における lacZ 遺伝子発現検出による
脊索発生の解析
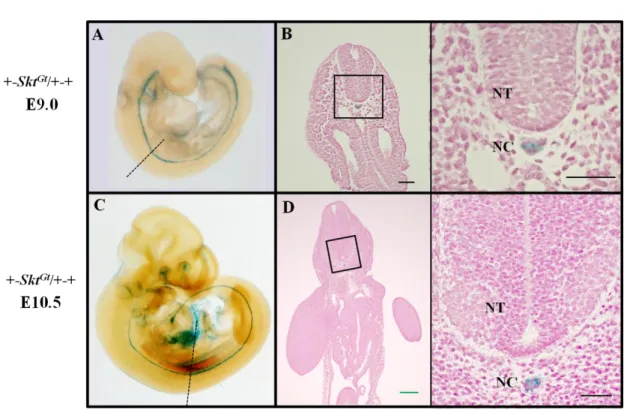
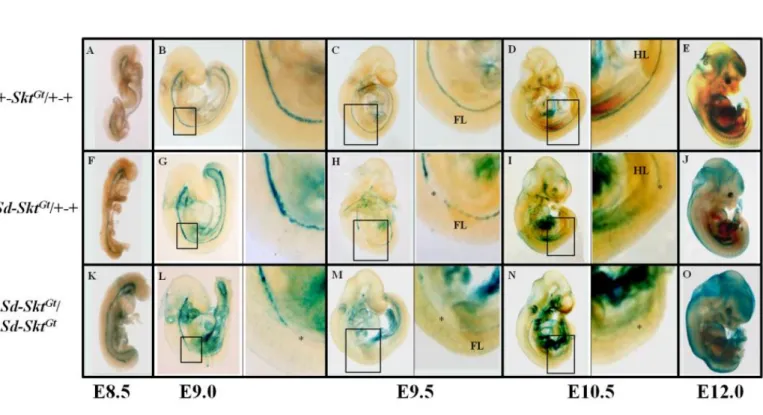
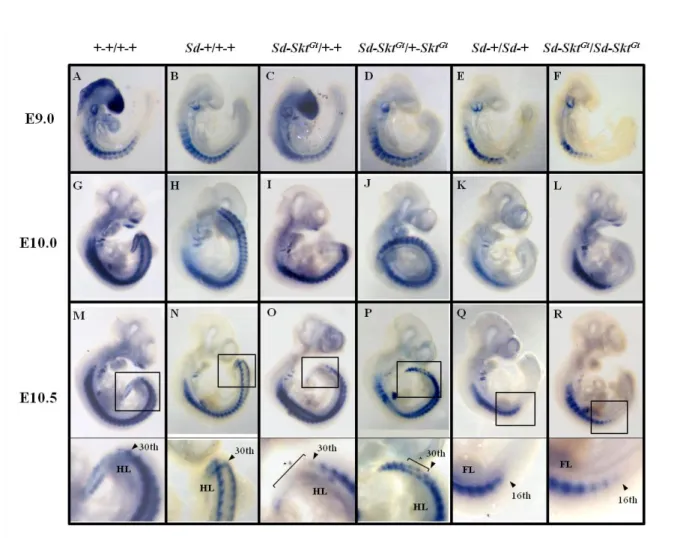
Skt locus に挿入された lacZ 遺伝子の脊索における発現は X-gal 染色によって検
出することができる(Fig. 5)ことから、E8.5、E9.0、E9.5、E10.5、E12.0 それぞれ
のステージにおける+-Skt
Gt/+-+、Sd-Skt
Gt/+-+、Sd-Skt
Gt/Sd-Skt
Gt胚に対し、whole
mount X-gal 染色法を用いて脊索形態のモニタリングを行った。+-Skt
Gt/+-+胚の
脊索では発生を通じて正常な
-galactosidase(
-gal)活性が検出された(Fig. 6A-E)。
Sd-Skt
Gt/+-+胚では E8.5 における
-gal 活性は+-Skt
Gt/+-+ 胚と同様であった(Fig.
6F)。しかし、E9.0 になると 5
th体節レベル以下で
-gal 活性の減弱化が観察され
(Fig. 6G)、E9.5 では同レベル以下の体節で検出されるシグナルが分断化していた
(Fig. 6H)。E10.5 になると 27
th体節レベルより近位での
-gal 活性が全く検出さ
れなくなり(Fig. 6I)、E12.0 では胚全長において活性が消失していた(Fig. 6J)。
Sd-Skt
Gt/Sd-Skt
Gt胚においても E8.5 では
-gal 活性が胚全長において認められた
(Fig. 6K)。E9.0 になると活性が 8
th体節レベル以下で減弱・分断化を認め(Fig. 6L)、
E9.5 以降では完全に消失していた(Fig. 6M, N, O)。さらに、E12.0 胚は下半身の
著明な低形成を示していた(Fig. 6O)。
31
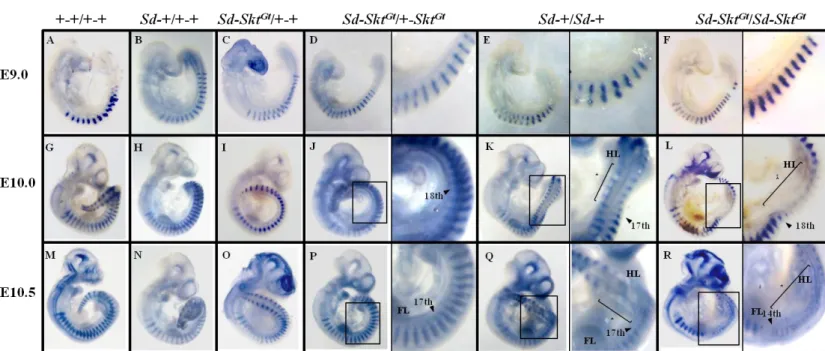
析
Sd変異マウスにおける脊椎欠損が、傍軸中胚葉の体節形成不全によるものかど
うかを調べるために、体節マーカー遺伝子である
UncxとParaxisを解析に用いた。
Uncxは新たに形成された体節の尾部半分で発現し、その発現異常は体節の前後
軸パターン形成の破綻を意味する。発生が進むとその発現は椎板外側に限局し、
そこに由来する脊椎の椎間関節、横突起、上肢などの形成に関わることが知ら
れている(Leitges et al., 2000)。bHLH型の転写因子であるParaxisは新たに形成さ
れた上皮性体節で強く発現し、その後体節の分化が進むに従ってその発現は減
弱していくことが知られている(Burgess et al., 1995; Johnson et al., 2001)。
ま ず 、
E9.0 、 E10.0 、 E10.5 ス テ ー ジ の wild-type 、 Sd-+/+-+ 、 Sd-Skt
Gt/+-+ 、
Sd-Skt
Gt/+-Skt
Gt、
Sd-+/Sd-+、Sd-Skt
Gt/Sd-Skt
Gt胚に対し、whole mount in situ
hybridization 法により Uncx の mRNA を検出した。今回解析したステージにおい
て
wild type 、 Sd ヘ テ ロ ア レ ル を 持 つ 変 異 体 (Sd-+/+-+, Sd-Skt
Gt/+-+ and
Sd-Skt
Gt/+-Skt
Gt)の胚における Uncx 発現は正常であった(Fig. 7A-D, G-J and M-P)。
一方、E9.0 Sd-+/Sd-+ 胚での Uncx 発現は wild type と同様であったが(Fig. 7E)、
E10.0 になると 17
th-22
nd体節での発現が減弱し(Fig. 7K)、E10.5 になると発現の減
弱領域がより後方(28
th体節)へ伸長した(Fig. 7Q)。Sd-Skt
Gt/Sd-Skt
Gt胚においても
E9.0 では正常な発現パターンを示し(Fig. 7F)、E10.0 で 17
th-24
th(Fig. 7L)、E10.5
32
になると
16
th体節レベル以下の発現が全て減弱しているといった(Fig. 7R)、より
重篤な表現型を示した。
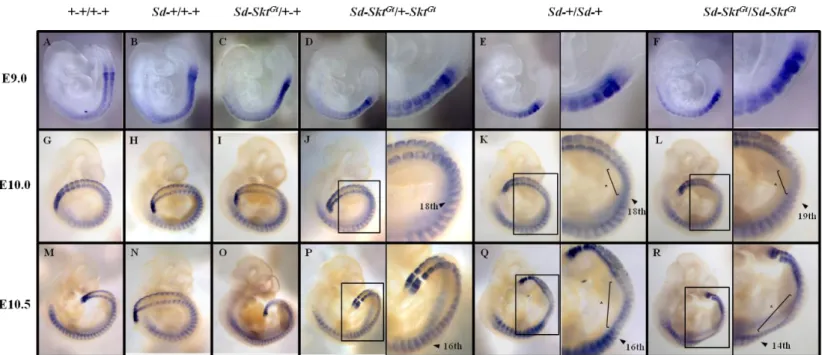
次に
E9.0、E10.0、E10.5 ステージの wild-type、Sd-+/+-+、Sd-Skt
Gt/+-+、Sd-Skt
Gt/+-Skt
Gt、
Sd-+/Sd-+、Sd-Skt
Gt/Sd-Skt
Gt胚を用いて、whole mount in situ hybridization 法によ
り
Paraxis の mRNA を検出した。wild type、Sd-+/+-+、Sd-Skt
Gt/+-+、Sd-Skt
Gt/+-Skt
Gt胚における
Paraxis 発現は Uncx の結果と同様に正常パターンを示した(Fig. 8A-D,
G-J and M-P)。Sd-+/Sd-+、Sd-Skt
Gt/Sd-Skt
Gt胚においても Uncx の結果と同様に E9.0
での発現は正常 (Fig. 8E and F)、E10.0 になると体幹下部での発現の減弱が認め
られ(Sd-+/Sd-+; 18
th-22
nd体節, Sd-Skt
Gt/Sd-Skt
Gt; 19
th-23
th体節, Fig. 8K and L)、E10.5
では発現の減弱領域の後方への伸長(Sd-+/Sd-+; 17
th-27
th体節, Sd-Skt
Gt/Sd-Skt
Gt;
15
th-28
th体節, Fig. 8Q and R)が認められた。
これらの結果は
Sd-+/+-+、Sd-Skt
Gt/+-+、Sd-Skt
Gt/+-Skt
Gt胚の傍軸中胚葉における
体 節 形 成 が 正 常 に 起 こ っ て い る こ と を 示 し て い る 。 ま た 、
Sd-+/Sd-+ 、
Sd-Skt
Gt/Sd-Skt
Gt胚の結果から Sd ホモ接合体においても体節形成は正常に起こ
っているが、E9.0 以降に何らかの原因により体幹下部における体節構造の維持
ができていないことが示唆された。
7-4)Sd, Skt
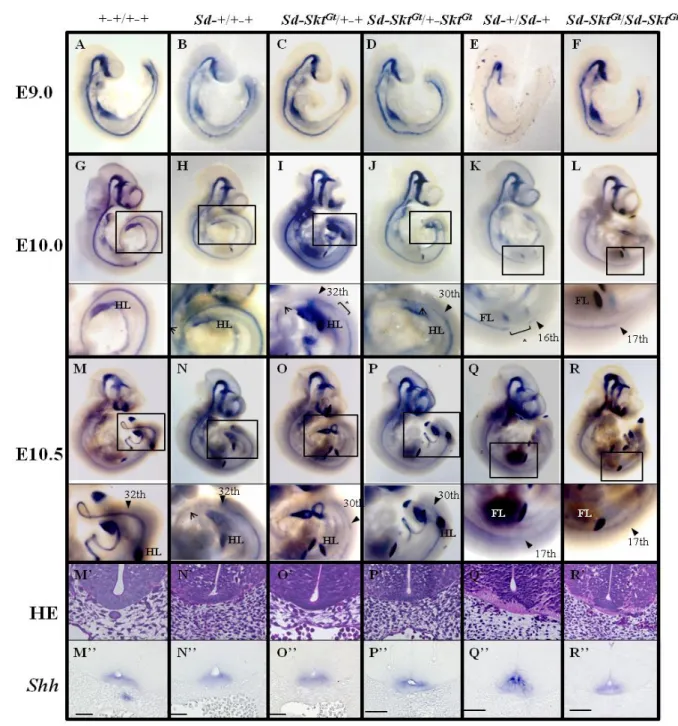
Gt変異マウス胚における神経管底板発生の解析
次に、
Sd 変異体胚における神経管底板の発生を調べるために E9.0、E10.0、E10.5
33
ステージの
wild-type、Sd-+/+-+、Sd-Skt
Gt/+-+、Sd-Skt
Gt/+-Skt
Gt、
Sd-+/Sd-+、
Sd-Skt
Gt/Sd-Skt
Gt胚を用いて、whole mount in situ hybridization 法により Shh の
mRNA を検出した。E8.5 での Shh 発現パターンは全ての genotype で同等であっ
た(data not shown)。wild type 胚での Shh 発現は今回解析したステージを通じて脊
索、神経管底板、肢芽、脳および腸管で検出された(Fig. 9A, G, M, M’’)。E9.0 で
の
Sd-+/+-+、Sd-Skt
Gt/+-+、Sd-Skt
Gt/+-Skt
Gt胚における発現パターンは wild type
と同等であった(Fig. 9B-D)。E10.0、E10.5 における Sd-+/+-+胚では脊索での Shh
発現が近位より分断化を示していたが、神経管底板での発現は体幹全長にわた
り保たれていた(Fig. 9H, N and N’’)。E10.0、E10.5 Sd-Skt
Gt/+-+、Sd-Skt
Gt/+-Skt
Gt胚
の神経管底板における
Shh 発現は 31
st体節レベル以降途絶していた(Fig. 9I, J, O,
P)。Sd-Skt
Gt/+-+、Sd-Skt
Gt/+-Skt
Gt胚における脊索での Shh 発現パターンは分断化、
消失が
Sd-+/+-+胚よりやや重篤になっているのが観察され、X-gal 染色の結果
(Fig.6)とよく一致していた。
Sd-+/Sd-+、Sd-Skt
Gt/Sd-Skt
Gt胚では E9.0 の時点ですでに神経管底板における Shh
発現の途絶が見られ(Sd-+/Sd-+; 18
th体節レベル, Sd-Skt
Gt/Sd-Skt
Gt; 17
th体節レベル,
Fig. 9E, F)、E10.0-10.5 と発生ステージが進行しても同レベル以降での発現は消
失したままであった(Fig. 9K, L, Q, R)。
34
作製、
Shh プローブを用いた section in situ hybridization (Fig. 9M’’-R’’)を行った。
wild-type 胚を除き、脊索の消失と形態的な神経管底板の形成、底板における Shh
発現が確認された。以上の結果から、
Sd 変異マウス胚では、脊索により神経管
底板の発生が誘導されると、脊索が消失してもその前に誘導された底板が
Shh
を分泌し続けることを示している。
7-5)Sd, Skt
Gt変異マウス胚における椎板発生の解析
最後に、
Sd 変異体胚における体節から椎板への分化を調べるために椎板マーカ
ーである
Pax1 遺伝子の RNA probe を用いた whole mount in situ hybridization 法を
行った。
対象としてこれまでの実験と同様
E9.0、E10.0、E10.5 ステージの wild-type、
Sd-+/+-+、Sd-Skt
Gt/+-+、Sd-Skt
Gt/+-Skt
Gt、
Sd-+/Sd-+、Sd-Skt
Gt/Sd-Skt
Gt胚を用いた。
wild type 胚における Pax1 の発現は E9.0 で 17
th、
E10.0 で 32
nd、
E10.5 で 38
th体節
レベルの椎板まで連続して検出された(Fig. 10A, G, M)。Sd-+/+-+、Sd-Skt
Gt/+-+、
および
Sd-Skt
Gt/+-Skt
Gt胚における
Pax1 発現は E9.0 と E10.0 においては正常胚と
同じパターンを示した(Fig. 10B-D, H-J)。E10.5 Sd-+/+-+胚における Pax1 発現は
wild type 胚とほぼ同等であった (Fig. 10N)。しかし、E10.5 の Sd-Skt
Gt/+-+、
Sd-Skt
Gt/+-Skt
Gt胚においては 30
th- 33
rd体節レベルでの Pax1 発現が減弱していた
(Fig. 10O, P)。Sd-+/Sd-+、Sd-Skt
Gt/Sd-Skt
Gt胚においても E9.0 での Pax1 発現は
35
以下の
Pax1 発現は全く見られず、E10.5 においても変化はなかった(Fig. 10K, L,
Q, R)。上記の結果により、Sd ホモ接合体胚では 17
th体節レベル以下の体節から
椎板への分化ができなくなっていることが示され、またそのレベルは神経管底
板の誘導レベル、形成された椎体数と一致していた。
7-6)Sd, Skt
Gt変異マウス胚における各組織のマーカー遺伝子発現と形成された
椎体数の比較
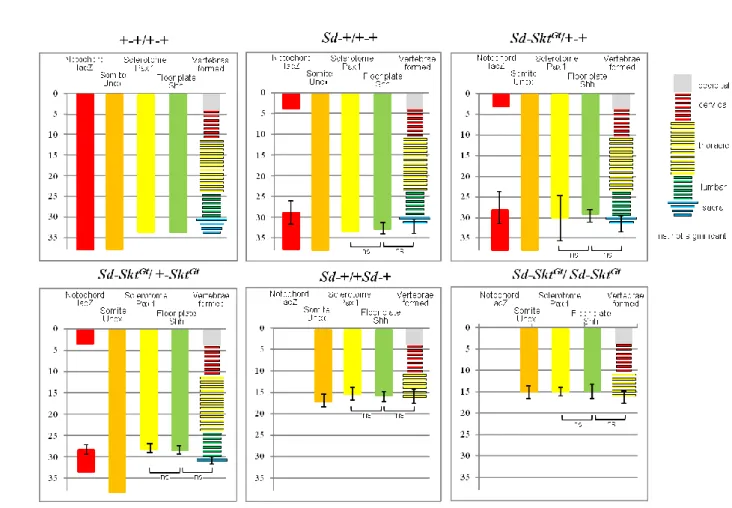
各マーカー遺伝子発現、椎体数の比較を容易にするために
E10.5 でのそれぞれの
genotype における発現レベルと E18.5 における形成された椎体数をグラフで表し
た(Fig. 11)。発生に神経管底板の影響がない尾椎はこのグラフにおいて省略して
いる。尾部においてはその腹側表面に存在する
ventral ectodermal ridge (VER)が
Shh ではなく、BMP のアンタゴニストである Noggin を発現して椎体を含む尾部
の発生、伸長を制御していることが明らかになっている(Goldman et al., 2000)。
このグラフにより、底板の形成されたレベルと椎板の誘導レベル、椎体数の間
に統計学的有意差なく一致していることが示され、脊索が消失しても神経管底
36
8) 考察
本研究の結果により種々の
genotype を脊椎欠損レベルより 3 群に分けることが
できる。脊椎発生の正常群(+-+/+-+, +-+/+-Skt
Gt)、中等度の脊椎欠損を示す群
(Sd-+/+-+, Sd-Skt
Gt/+-+ and Sd-Skt
Gt/+-Skt
Gt)、そして重篤な脊椎欠損を示す群
(Sd-+/Sd-+, Sd-Skt
Gt/Sd-Skt
Gt)である。ここでは中等度群、重篤群の代表として
Sd-Skt
Gt/+-+、Sd-Skt
Gt/Sd-Skt
Gtの結果について論じる。
Sd-Skt
Gt/+-+マウスの尾椎を
除いた脊椎数は
27 であり、Shh、Pax1 の発現は 30
th体節レベルまで検出されて
いる。一方、
Sd-Skt
Gt/Sd-Skt
Gtマウスの脊椎数は
13 であり、Shh、Pax1 の発現は
17
th体節レベルまで検出されている。1
st-4
th体節は脊椎ではなく後頭骨に分化す
るため(Table. 3) (Gossler and Tam, 2002)、形成された椎体数に 4 を加えるとマー
カー遺伝子発現体節レベルとよく一致していることがわかる(Fig. 11)。また、体
節 マー カー遺伝 子(Uncx, Paraxis)の発現は Sd-Skt
Gt/+-+胚では正常であり、
Sd-Skt
Gt/Sd-Skt
Gt胚においては
17
th体節以下での減弱化が見られるものの発現パ
ターンは正常群と同様であった。このことは
Sd 変異体における椎板への分化異
常、また脊椎形成異常が傍軸中胚葉の体節形成異常によるものではないことを
示している。今回の研究の結果では脊索が消失する前に
Sd-Skt
Gt/+-+胚では 30
th体節レベル、
Sd-Skt
Gt/Sd-Skt
Gt胚においては
17
th体節レベルまで神経管底板の形成
を誘導し、形成された神経管底板が脊索機能を代替してその後の体節の椎板へ
37
の分化、ひいては脊椎への分化誘導過程を維持できることが示された。これま
で脊椎発生における脊索の役割は隣接する神経管の腹側に底板を誘導し、底板
と協調して周囲体節組織を椎板へ分化誘導することだと考えられていた。しか
し、本研究の結果により脊椎発生における脊索の役割は底板の誘導が主である
ことを強く示唆している。
Sd変異マウス解析におけるSkt
Gtアレルの有用性
我々は以前の研究でジーントラップ法を用いたmutagenesisによりSkt
Gtマウスラ
インを作製し、報告した(Semba et al., 2006)。このSkt遺伝子はSd locusの約1cM下
流に位置し、早期胚より脊索で発現する。掛け合わせにより
Sd、Skt
Gtアレルが
同一染色体に存在する
Sd-Skt
Gtダブル変異マウスを得ることができ、
Skt
Gtアレル
に挿入された
lacZ遺伝子を用いることでSd変異体の脊索の形態のモニタリング
とgenotypingの両者が可能となった。本研究の結果より、Skt
Gtアレルは
Sd変異を
解析する上で脊索形態のモニタリングだけでなくgenotypingとしても非常に有
用なツールであった。
遺伝的背景が
Sd 変異マウスの脊椎形成に与える影響
これまでの報告における
Sd マウスの genetic background は Jackson Laboratory が
提供している
RSV/LeJ を用いているものがほとんどである。今回の研究で我々
38
途絶しており、過去の報告の中で最も重篤な表現型を示した。これは genetic
background が表現型に影響を与える典型的な例と言えるがそのメカニズムにつ
いては本研究では明らかにすることができなかったが、
Sd 変異マウスの脊椎欠
損を最大に示す
C57BL/6 genetic background マウスを使用した事が本研究結果を
示す上でのアドバンテージとなった。今後
129/Sv 系統等他の近交系マウスの
genetic background を用いて Sd マウスを作製し、Sd 変異の脊椎欠損の表現型に影
響を与える
modifier の検討を行うのも興味深い研究となるであろう。
Sickle tail 遺伝子の脊椎形成における機能
Skt
Gtホモ接合体マウスは椎間板形成異常による尾椎先端の屈曲変形を示し、Skt
遺伝子は
proline-rich region とケラチン、ミオシンなどの線維性タンパクの重合
に必要とされる
coiled-coil domain を持つことが先の研究で明らかになっている。
また、
Skt 遺伝子は発生初期より脊索において発現し、成体においても脊索の遺
残である椎間板髄核で発現し続ける(Semba et al., 2006)。また連鎖解析の結果、
マウス
Skt 遺伝子のヒトホモログである KIAA1217(SKT)に存在する SNP が腰椎
椎間板ヘルニアの危険因子となることも明らかにされており(Karasugi et al.,
2009)、椎間板構造の強度保持に関連する遺伝子であることが示唆されている。
本研究では
Skt
Gtアレルが
Sd 変異体の下位脊椎(仙椎、尾椎)形成に影響を及ぼす
一方、上位脊椎には影響を与えないことが明らかになった。つまり、今回の結
39
果は
Skt 遺伝子が椎間板のみならず下位脊椎の形成に関する何らかの機能を持
っていることが示唆されており、今後さらなる機能解析が必要と考えている。
本研究により、Sd 変異胚は、一度発生した脊索が早期に消失するものの、脊索
が消失するまでに脊索が存在した領域でのみ底板組織が発生することがわかっ
た。また、分化誘導された底板のレベル・椎板のレベルは、後期胚で確認され
た中軸骨格の欠損レベルと一致していることがわかった。このことより、脊椎
発生において脊索は、底板誘導時期まで関与しており、一度底板を誘導した後
は主に底板が脊椎発生の中心組織になっている事を示していると考えられた。
いまだ不明な点が多い
Sd 変異マウスの表現型の主因は、脊索構造の破綻による
ものと考えられる。今後の解析により、
Sd 責任遺伝子の同定、またタンパク機
能解析を明らかにすることで、脊索発生、脊椎発生の更なる知見を深めること
ができるであろう。
40