Terumi Ikawa
INTRODUCTION
Although insects surpass all other living organisms on the earth in species numbers, they are relatively unsuccessful in the marine realms. Out of one million extant insect species, a few thousands are marine (Cheng and Frank 1993) and only fi ve species of ocean skaters
(Fig. 1), and , have successfully
colonized in the high seas (Andersen and Polhemus 1976 ; Cheng 1985 ; Andersen and Cheng 2004 ; Ikawa et al. 2012). Thus, ocean skaters are quite unique organisms from the viewpoint of ecology and evolutionary biology. They occupy vast areas in the tropical and subtropical region of three major oceans (Andersen and Polhemus 1976 ; Cheng 1985, 1989 ; Ikawa et al. 2012). In fact, they are distinctive not only in having extended their habitat to the open ocean but also in having the largest distribution range among the insects. One of the most intriguing questions about the ecology of ocean skaters is about their unusual distribution ranges, e.g., how the fi ve
species of oceanic are distributed in the world oceans and what determines their
distribution ranges.
Herring(1961) was the fi rst to construct the worldwide distribution maps of ocean skaters, compiling collection records from diff erent cruises over the years. Since then, several authors have updated the maps, adding more data from more cruises (Cheng 1973, 1985 ; Andersen and Polhemus 1976) and examining the factors that delimit the distribution ranges (Cheng 1989 ; Ikawa et al. 2012). However, these maps are still incomplete due to the lack of sampling data in some areas of the seas, because, although hitherto numerous research cruises have been organized by universities, institutions and national/international projects, the world oceans are still too immense to be fully covered and investigated. More samplings, together with
Figure 1. Male adult of ocean skater .
environmental data, are defi nitely necessary to complete the distribution maps and grasp the factors infl uencing the distribution ranges of ocean skaters.
Because the worldwide distribution maps are based on the compilation of sampling data from different seasons and years, a problem inevitably arises, i.e., seasonal/annual changes have been totally neglected although distribution ranges of living organisms are not static but change, according to temporal changes of the environments. Therefore, the next essential step is to study temporal changes of distribution ranges and responsible factors. However, regular research cruises to survey fi xed sites of the sea are seldom organized making this study quite
diffi cult, and so far, only a handful of studies include repeated surveys of populations (in
the Banda Sea by Cheng et al. 1990 ; in the Atlantic Ocean by Cheng 1973 ; in the eastern Pacifi c Ocean by Engel et al. 2014 ; in the western Pacifi c Ocean by Ikawa et al. 2002 ; off the Japanese coast by Ikawa et al. 2004).
With the above in mind, samplings of ocean skaters were made in the westernmost area of the North Equatorial Current in the western Pacifi c Ocean in 2007 during the R/V Hakuho Maru cruise. The area surveyed belongs to a peculiar region where the distribution ranges
of three species, and , signifi cantly overlap (see the maps
in Cheng 1985 ; Ikawa et al. 2012). This region is ca. 7̊N 33̊N of the western Pacifi c Ocean, hereafter referred to as the overlap region. Except for this overlap region, the distribution
ranges of the fi ve oceanic appear to be well separated. Ikawa et al. (2002) conducted
samplings during R/V Hakuho Maru cruises in 1994, 1995 and 1998 in the westernmost area of North Equatorial Current and showed that only one species was found in high density during each cruise but the dominant species was diff erent among cruises. Thus, the temporal change of dominant species appeared to have had caused the temporal changes in distribution ranges, possibly generating apparent overlaps in the distribution maps. The accumulation of more sampling data from diff erent years and seasons is indispensable to clarify the temporal changes of distribution ranges and responsible factors in this region.
Thus, the purposes of this study are as follows : firstly, to provide sampling data to contribute to the completion of worldwide distribution maps ; secondly, to investigate species composition and densities to determine the dominant species in the study area ; thirdly, to compare obtained data with past data during the Hakuho Maru criuses of 1994, 1995 and 1998 so as to get a deeper insight into the temporal changes of the dominant species and distribution ranges.
MATERIALS AND METHODS
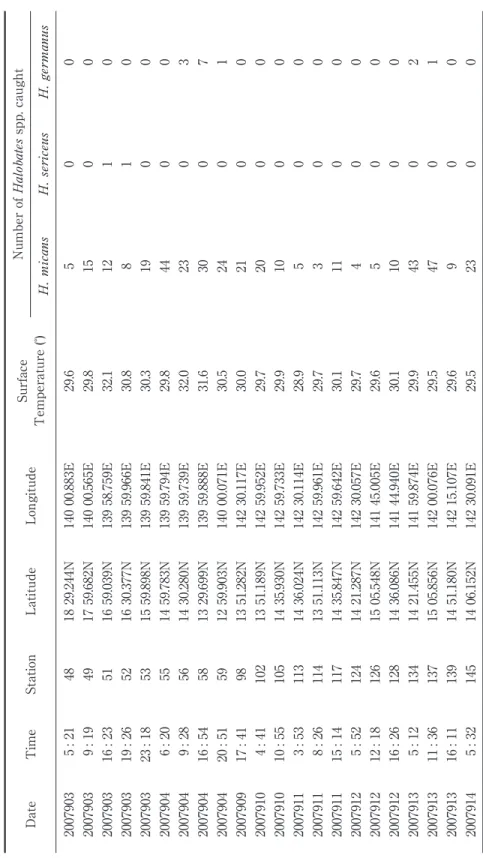
During the cruise of R/V Hakuho Maru KH07-2 Leg.3 conducted in September 2007, ocean skaters were collected at 22 stations (Fig. 2A, 2B) in the area between latitudes ca. 13̊N 18.5̊N and longitudes ca. 140̊E 143̊E. This area belongs to the area commonly surveyed during the past three R/V Hakuho Maru cruises in 1994, 1995 and 1998 (Ikawa et al. 2002). Ocean skaters were sampled by the use of a neuston net (40 x 100 cm in mouth opening and 450 µm mesh size) towed at the sea surface at a speed of 2 3 knots for 10 35 min. The ocean skaters captured were sorted on board. Adult females were either identifi ed and dissected under microscope on
board or were stored in 1:1 physiological solution and 99% ethanol for further investigation in the laboratory. Male adults and nymphs were preserved in 99% ethanol and identifi ed in the laboratory. The distance towed was estimated by the use of the fl owmeter attached to the net. The areas towed were calculated by multiplying the width of the net opening by the distance
towed. Densities of spp. were estimated by dividing the number of insects caught by
the towed area.
RESULTS AND DISCUSSION
From the distribution maps (Cheng 1985, 1989 ; Ikawa et al. 2012), it was expected that three
species, , would be collected in the study area. As is shown
in Table 1, all three species were collected. (Fig. 1) was caught from all 22 tows.
The number of individuals for each tow ranged from 3 to 44, and in total 391 individuals were
collected. was collected from fi ve tows and in total 14 individual were sampled.
was collected from only two tows and two individuals were sampled. The estimated
densities in the study area were 11,000 individuals/km2 for , 400 individuals/km2 for
and 60 individuals/km2 for . Thus, was dominant in the study
Figure 2A. The area surveyed (hatched) by the Hakuho Maru cruise KH07-2.
Figure 2B. The enlarged map of the area surveyed in Fig. 2A. Dots are locations of samplings with station numbers.
Table 1. List of dates, stations, locations, sea-surface tempe
ratures, and numbers of spp. caught.
Date Time Station Latitude Longitude Surface Temperature (̊) Number of spp. caught 2007903 5 : 21 48 18 29.244N 140 00.883E 29.6 5 0 0 2007903 9 : 19 49 17 59.682N 140 00.565E 29.8 15 0 0 2007903 16 : 23 51 16 59.039N 139 58.759E 32.1 12 1 0 2007903 19 : 26 52 16 30.377N 139 59.966E 30.8 8 1 0 2007903 23 : 18 53 15 59.898N 139 59.841E 30.3 19 0 0 2007904 6 : 20 55 14 59.783N 139 59.794E 29.8 44 0 0 2007904 9 : 28 56 14 30.280N 139 59.739E 32.0 23 0 3 2007904 16 : 54 58 13 29.699N 139 59.888E 31.6 30 0 7 2007904 20 : 51 59 12 59.903N 140 00.071E 30.5 24 0 1 2007909 17 : 41 98 13 51.282N 142 30.117E 30.0 21 0 0 2007910 4 : 41 102 13 51.189N 142 59.952E 29.7 20 0 0 2007910 10 : 55 105 14 35.930N 142 59.733E 29.9 10 0 0 2007911 3 : 53 113 14 36.024N 142 30.114E 28.9 5 0 0 2007911 8 : 26 114 13 51.113N 142 59.961E 29.7 3 0 0 2007911 15 : 14 117 14 35.847N 142 59.642E 30.1 11 0 0 2007912 5 : 52 124 14 21.287N 142 30.057E 29.7 4 0 0 2007912 12 : 18 126 15 05.548N 141 45.005E 29.6 5 0 0 2007912 16 : 26 128 14 36.086N 141 44.940E 30.1 10 0 0 2007913 5 : 12 134 14 21.455N 141 59.874E 29.9 43 0 2 2007913 11 : 36 137 15 05.856N 142 00.076E 29.5 47 0 1 2007913 16 : 11 139 14 51.180N 142 15.107E 29.6 9 0 0 2007914 5 : 32 145 14 06.152N 142 30.091E 29.5 23 0 0
area during this cruise and and can be considered strays.
Considering the fact that we caught only a small number of during this cruise
and none during the three other Hakuho Maru cruises conducted in 1994, 1995, and 1998 (Ikawa
et al, 2002), the density of seemed constantly low or rarely became high in the
study area. Compared to other oceanic species, tends to be distributing near the
coastlines of islands or continents (Andersen and Polhemus 1976 ; Cheng 1985). Therefore, the
study area might have been too far from the coasts for to be collected in high
densities.
The dominant species was during this cruise and the Hakuho Maru cruises of
1994 and 1995 while it was replaced by during the Hakuho Maru cruise of 1998
(Ikawa et al. 2002), Thus, only one species was dominant during these four cruises in the study area and it can be concluded that the distribution ranges of the three species did not overlap but stayed separate although there were some strays of other species. The overlap in the worldwide distribution maps in the study area would have been generated from overlaying sampling data from diff erent seasons and years. So far, off the Japanese coast is the only area known where two diff erent species are co-dominant and the distribution ranges actually overlap (Ikawa et al. 2004). The Kuroshio Current is considered to be responsible for this overlap. Eddies generated by the frontal disturbance of this strong western boundary current would provide heterogeneous environments for diff erent species to co-exist.
1) Sea surface temperature
The sea surface temperature (SST) is considered to be one of the most important environmental factors to determine the distribution ranges of ocean skaters (Cheng 1985 ;
Andersen and Cheng 2004 ; Ikawa et al. 2012). It appears that prefers warmer waters
compared to , and in the North Pacifi c Ocean occupies the cooler area
north of the tropical habitat zone of .
As shown in Table 1, in all the sampling stations the SST was quite high, ranging from 29.6̊C to 32.1̊C. During the other three cruises the SST remained as high as during this cruise (Ikawa et al. 2002). Actually, the area sampled belongs to the western Pacifi c warm pool where the SST remains high throughout the year (Delcroix 1998). Therefore, the SST can be eliminated from the factors responsible for the change of the dominant species during the four cruises. 2) Shift of salinity front
In the North Pacifi c Ocean, occupies the northern area of habitat
zone where the water is less saline due to much rainfall. Therefore, the salinity front, existing between the northern high salinity water and the southern low salinity water, may serve to separate the two species. Kimura et al. (2001) found that during normal years the salinity front was located at ca. 16̊N while during years of the El Nino events the salinity front shifted southward to ca. 10̊N. The boundary of the distribution ranges of the two species would be
expected to shift toward the south or north with the shift of the salinity front. As a matter of
fact, was predominant in the study area during the Hakuho Maru cruise of 1998
when the El Nino event was observed and the salinity front was located at ca. 10̊N. On the
other hand, was predominant during the cruises of 1994 and 1995 when the salinity
front was at ca. 16̊N (Ikawa et al. 2002). During this cruise in 2007, the salinity front was found at ca. 18.5̊N along 140̊E (Zenimoto et al. 2007). Therefore, almost all the sampling stations for
ocean skaters were located south of the salinity front and there was dominant. Thus,
the shift of the salinity front would serve to replace the dominant species and change the distribution ranges.
3) Seasonal changes of prevailing winds
Because ocean skaters live exclusively on the sea surface where there is no refuge for them, the winds would strongly infl uence their distributions. The seasonal changes of prevailing winds
could shift the distribution ranges of spp., carrying them away for a long distance on
the ocean surface (Ikawa et al. 2002, 2004, 2007). In this study area, the direction of prevailing winds is southwestward from November to April and then it reverses northward from June to
September (Kutsuwada 1987). In the North Pacifi c Ocean, lives north of the habitat
zone of and the boundary is between 12̊N 20̊N (Cheng 1985, 1989 ; Ikawa et al.
2012). Blown by the seasonal prevailing winds, their boundary could shift toward the south in the winter season and toward the north in the summer season (Ikawa et al. 2012). Because the samplings of the present cruise were conducted in September and those of 1994 in June-July, and 1995 in July-September, the boundary between the two species might have shifted north
and as the result, might have been predominant in the study area. On the other hand,
those of 1998 were conducted in May June, a little earlier than other three cruises. Hence, the boundary of the two species might have remained south of the sampled area and
might have been dominant.
So far, the results of the present cruise could be explained without contradicting the discussions of a previous paper (Ikawa et al. 2002) and could confi rm its conclusion. However, more repeated samplings are indispensable. Especially since all the samplings of ocean skaters of this cruise and the other three cruises of 1994, 1995 and 1998 were conducted from May to September, we have no information about distributions in the other seasons. To understand spatio-temporal features of distributions and densities of ocean skaters, it is essential to
accumulate more seasonal and annual samplings in spite of the diffi culties to organize regular
cruises in the same region of the open ocean.
ACKNOWLEDGEMENTS
The author off ers sincere thanks to Professor K. Tsukamoto, the chief scientist of KH07-2, for his warm encouragement and generous support which made this research possible under the very tight sampling schedule for the quest of eel eggs. I am grateful to Captain Inaba, the crew, Mr. M. Sasaki, Ms. W. Taki and all other scientists on board for their help during the cruise, to Drs. H. Okabe, S. Hoshizaki and M. Watanabe for helpful discussions and to Ms. R. Takeya and
Ms. N. Yamashita, for their help with sorting and identifying the specimens in the laboratory. My heartfelt thanks go to the fellow scientists and the crew of the four-to-eight watch for their skillful assistance and joyful cooperation throughout this cruise. Finally, I thank the anonymous reviewer for the valuable comments to improve this paper.
REFERENCES
Andersen NM, Cheng L. (2004) The Marine insect (Heteroptera : Gerridae) : biology, adaptations, distribution, and phylogeny. 42 : 119 180.
Andersen NM, Polhemus JT. (1976) Water-striders (Hemiptera : Gerridae, Veliidae, etc.). In : Cheng L. (ed) , pp.187 224. North-Holland, Amsterdam, Netherlands. 581 pp.
Cheng L. (1973) . Oceanography and Marine Biology, An Annual Review 11 : 223 235.
Cheng L. (1985) Biology of (Heteroptera : Gerridae). 30 : 111 134.
Cheng L. (1989) Factors limiting the distribution of species. In : Ryland JS, Tyler PA (eds) Reproduction, , pp.357 362. Olsen and Olsen, Fredensborg, Denmark. Cheng L, Baars MA, Oosterhuis SS. (1990) in the Banda Sea (Indonesia) : monsoonal diff erences in
abundance and species composition. 47 : 421 430. Cheng L, Frank JH. (1993) Marine insects and their reproduction.
31 : 479 506.
Delcroix T. (1998) Observed surface oceanic and atmospheric variability in the tropical Pacifi c at seasonal and ENSO time scale : A tentative overview. 103 : 18611 18633.
Engels M, Correia L, Piwinski S, Cheng L, Zettler E. (2014) Seasonal and decadal changes in distribution patterns of (Hemiptera : Gerridae) populations in the eastern tropical Pacifi c. 161 : 1241 1250. Herring JL. (1961) The genus (Hemiptera : Gerridae). 3 : 223 305.
Ikawa T, Okabe H, Cheng L. (2012) Skaters of the seas―comparative ecology of nearshore and pelagic (Hemiptera : Gerridae), with special reference to Japanese species. 8 : 915 936. Ikawa T, Okabe H, Hoshizaki S, Fuchi T, Cheng L. (2002) Species composition and distribution of ocean skaters
(Hemiptera : Gerridae) in the western Pacifi c Ocean. 5 : 1 6. Ikawa T, Okabe H, Hoshizaki S, Kamikado T, Cheng L. (2004) Distribution of the oceanic insects
(Hemiptera : Gerridae) off the south coast of Japan. 7 : 351 357.
Ikawa T, Onodera S, Okabe H, Hoshizaki S, Cheng L. (2007) Occurrence and density of (Hemiptera : Gerridae) in the eastern South Indian Ocean. 10 : 213 215.
Kimura S, Inoue T, Sugimoto S. (2001) Fluctuation in distribution of low-salinity water in the North Equatorial Current and its eff ect on the larval transport of the Japanese eel. 10 : 51 60.
Kutsuwada, K. (1987) Monthly maps of surface wind stress fi elds over the North Pacifi c during 1961 1984. 24, 1 100.
Zenimoto K, Miyazaki S, Yamaoka N, Kitagawa T. (2007) Studies on physical environments of eel larval transport. 25 27.