Efficient Purification of an Antihemorrhagic Factor (HSF) in Serum of Japanese Habu (Trimeresurus flavoviridis)
Masanobu DESHIMARU, Chie TANAKA, Aika TOKUNAGA, Masaki GOTO, and Shigeyuki TERADA
Department of Chemistry, Faculty of Science, Fukuoka University, 8‑19‑1 Nanakuma, Jonan‑ku, Fukuoka 814‑0180, Japan
((Received May 31, 2003)
Abbreviations: FTC, fluorescein isothiocyanate labeled; HSF,‑ antihemorrhagic factor in serum of Trimeresurus flavoviridis; MALDI‑TOF‑MS, matrix assisted laser desorption/ionization time of flight mass spectrometry; TFA,‑ ‑ ‑
trifluoroacetic acid.
Abstract
In order to attain the efficient purification anti hemorrhagic factor(HSF)from Habu‑ snake serum, we examined fractional precipitation by addition of ammonium sulfate or ethanol to the serum. The precipitate of high quality HSF was obtained by 45‑60%
saturation of ammonium sulfate or addition of 1.0‑1.25 volume of ethanol to the serum.
Further purification of the fractionated HSF by HPLC and gel filtration gave a single band around 68 kDa on SDS‑PAGE and its molecular mass was 47,810 by MALDI‑TOF‑
MS. The purified HSF bound to brevilysin H6 from Chinese mamushi snake venom
(Kd=3.7×10‑9)with 1:1 stoichiometry. It remarkably suppressed hemorrhage in dose‑ dependent manner when injected to mice with crude Habu snake venom. These results indicate that the fractional precipitation by ammonium sulfate or especially ethanol can be served as a useful starting method on HSF purification.
Introduction
While snake bite has been a serious medical problem for humans, certain warm‑blooded animals and snakes themselves have a remarkable resistance to envenomation of snakes.1‑3)
Naturally occurring factors that neutralize snake venom hemorrhagins or decrease the localized and lethal effects have been found in the sera of opossums, hedgehogs, mongooses, raccoon, squirrels, woodrats, and various snakes as well as in the venoms.4‑7)
Two antihemorrhagic proteins, HSF and BJ46a, have been purified from the sera of venomous snakes Trimeresurus flavoviridis and Bothrops jararaca, respectively.6,8)They are acidic glycoproteins having no proteolytic activity.
On the basis of primary structures, they were classified as a member of fetuin family that possesses a double headed cystatin domain.‑ Despite of the presence of cystatin domain, HSF are inhibitory to neither papain nor cathepsin B.6)
The firstly reported method to purify HSF
has been rather complicated and time consuming.‑ 9)
Since HSF binds to a hemorrhagin HR1,9)an improved method was then introduced that consists of an affinity chromatography on HR1‑conjugated Sepharose 4B, gel filtration on Sephadex G‑200, and reverse phase HPLC on‑ a phenyl column. Although the purification by affinity chromatography allowed the specific and large scale preparation of HSF, this method still has some defects: the prior purification of large amount of HR1 was required and a high concentration of guanidine HCl used as an eluent‑ will destroy the activity of the ligand, resulting in the great decrease of binding affinity to HSF.
In order to further improve the purification method for HSF, a rapid and easy procedure has been examined for the first step. We here describe a new method for the preparation of HSF, which consists of ethanol precipitation, preparative reverse phase HPLC,‑ and gel filtration on a Sephacryl S‑200HR column.
Materials and Methods
Materials: Blood of Japanese Habu(T. flavoviridis)
in Amami Oshima Islands was collected by decapitation, and the serum was collected by centrifugation. The venom of T. flavoviridis was also collected, lyophilized, and stored at
‑20℃. Low molecular weight metalloproteases corresponding to H proteinase,2‑ HR2a, and HR 2b were purified from T. flavoviridis venom.
FTC casein was prepared according to
10,11) ‑
Twining.12)All other chemicals were purchased from Wako Pure Chem. Inc.(Osaka). Fractionation of Snake Serum by Ammonium Sulfate: Solid ammonium sulfate was added into the snake serum(50 ml)to 30% saturation and stirred for 2 h at 4℃. The precipitate
(P‑30) was collected by centrifugation at 7,000rpm at 4℃ for 15 min, and the supernatant fluid was used to further fractionation by ammonium sulfate to 45% and 60% saturations
to give P‑45 and P‑60. All the precipitates were dissolved in water, dialyzed against water, and lyophilized. The supernatant after 60%
saturation was used without any purification.
Fractionation of Snake Serum by Cold Ethanol:
To a 50 ml of the snake serum, a 25 ml of ice cooled ethanol was added in drop wise.‑ The mixture was kept at 0℃ for 1 h under stirring. The precipitate(E )was collected0.5
by centrifugation at 7,000 rpm at 4℃ for15 min.
To the supernatant fluid, a 12.5 ml of ethanol was added. The mixture was stirred at 0℃
for 1 h, and centrifuged to give precipitate
(E0.75)and supernatant. Successive additions of ethanol(12.5 ml each)to the supernatant yield five additional fractions(E1.0, E1.25, E1.5, E1.75, and E )2.0 . All the precipitates were dried in vacuo at room temperature for 2 h using a vacuum drying oven DP23(Yamato), and then dissolved in water and lyophilized.
Column Chromatography: An analytical reverse‑ phase HPLC was done on a TSK‑GEL ODS‑
120T(0.46×25 cm)column(Tosoh)with a linear gradient of acetonitrile in 0.1% TFA at a flow rate of 1.0 ml/min, and elution was monitored at 230 nm. Preparative HPLC was done on a µBondasphere 5µ‑C8‑300Å(1.9×
15 cm)column (Waters)at a flow rate of 5.0 ml/min, and elution was monitored at220 nm using a flow cell of 1‑mm light path. Gel filtra‑
tion was done on a Sephacryl S‑200HR column
(2.6×94 cm)equilibrated with 50 mM Tris‑
HCl(pH 8.0).
Measurement of inhibitory activity to proteinases:
Inhibitory activity was measured as follows.
Various amounts of sample were mixed with proteinase solution and left at 0℃ for 5 min.
Proteolytic reaction was initiated by the addition of FTC casein(final‑ 0.2%). After incubation at 37℃ for 15 min, the reaction was terminated by adding of 6% trichloroacetic acid containing 1 mM EDTA. After standing 30 min in the dark, the mixture was centrifuged at 3,000×
g for 15 min, and the supernatant was diluted
Table 1. Inhibitory activity of ammonium sulfate fractions of T. flavoviridis serum.a)
ten times with‑ 0.5 M Tris HCl (pH‑ 8.5). The fluorescence was analyzed on a FP‑550A spectrofluorometer(Jasco)at 520 nm with the excitation at 490 nm. Buffers used for digestion of FTC casein were‑ 5 mM CaCl2‑40 mM Tris HCl(pH 8.5)and 80 mM Tricine NaOH(pH
‑ ‑
9.0)for brevilysin H6 and other proteinases, respectively.
Electrophoresis: S D S polyacryla mide gel‑ electrophoresis(SDS‑PAGE)was carried out on 10‑12.5% gel by Laemmli's method.13)Bovine serum albumin(67 kDa), ovalbumin(45 kDa), carbonic anhydrase(30 kDa), chymotrypsinogen
(25 kDa), Kunitz soybean trypsin inhibitor
(20.5kDa),lysozyme(14.3kDa),and cytochrome c(12.4 kDa) were used as the molecular weight marker proteins. After running under the constant current(10‑20 mA)for several hours, the gels were stained by 0.1% Coomassie brilliant blue R‑250 and destained by 10%
acetic acid.
Mass spectrometric analysis: Mass spectrum was measured on a Voyager DE‑STR MALDI‑
TOF mass spectrometer‑ (PerSeptive Biosystems). Sample was dissolved in 0.1% trifluoroacetic acid‑50% acetonitrile containing α cyano‑ ‑ ‑4 hydroxycinnamic acid(10 mg/ml)as the matrix, and 2‑µl aliquots were analyzed. Spectrum was calibrated by the molecular mass of apomyoglobin.
Protein Sequence Analysis: The amino acid sequences of proteins were determined by an automatic protein sequencing system PPSQ‑21
(Shimadzu).
Determination of hemorrhagic activity: Hemorrhagic activity was determined by the method of Kondo et al. with some modifications.14)Briefly, 5 µg of crude T. flavoviridis venom in a 50 µl of saline were injected intracutaneously in abdomen of mice. The mice were killed after 4 h and the skin removed immediately.The cross diameters‑ of each hemorrhagic spot were measured from the visceral side of the removed skin. The hemorrhagic activity was evaluated from the average diameter.
Results
Fractionation of the serum by ammonium sulfate or ethanol:
The snake serum was fractionated by ammonium sulfate yielding 4 fractions(P‑30, P‑45, P‑60, and the supernatant of P‑60), and their inhibitory activities to HR2a were measured. Though a majority of inhibitory activity was observed in P‑60, some activities were also found in P‑45(Table 1). The purity of HSF in these fractions was determined by SDS‑PAGE. As shown in Fig. 1A, the major component was a 67‑kDa protein. This protein was estimated to be albumin with respect to
Fig. 1. SDS‑PAGE analysis of the serum fractions.(A)Fractions(P‑30‑P‑60)after ammonium sulfate precipitation. A 12% gel was used.(B)Fractions(E0.5 ‑E )2.0 after ethanol precipitation. A 12.5% gel was used. M, marker proteins. Arrows indicate the band of HSF.
Fig. 2. Inhibition of T. flavoviridis metalloproteases.
Each of 40 µg/ml of H ‑proteinase(●),2
HR2a(○),and HR2b(△)was mixed with various amounts of P‑60, and the residual protease activity was measured at 37℃ and pH 9.0.
the molecular size and the abundance. It was reported that the molecular weight of HSF was 65,000‑70,000. A faint band found just above that of albumin was considered as the band of HSF in SDS‑PAGE of P‑45 and P‑60.
The effect of P‑60 on the proteinase activities of three proteinase preparations from T. flavoviridis venom was examined. H proteinase showed2‑ a high proteinase activity, while HR2a and HR 2b had much lower activities. HR2b showed the highest susceptibility to P‑60, while the non hemorrhagic H proteinase was inhibited‑ 2‑ about 10 times weakly(Fig. 2).
The serum was also fractionated by cold ethanol. After addition of 0.5‑2.0 volumes of ice cooled ethanol,‑ the precipitates were collected by centrifugation. Seven fractions(E0.5‑E )2.0
were obtained, and the inhibitory activities to HR2a are listed in Table 2. With increasing concentration of ethanol, the inhibitory activity also increased. Though the specific activities of E1.25and E1.5 were the highest,E1.25contained inhibitory activity at much higher level rather than E1.5. SDS‑PAGE analysis also indicated
Table 2. Inhibitory activity of ethanol fractions of T. flavoviridis serum.a)
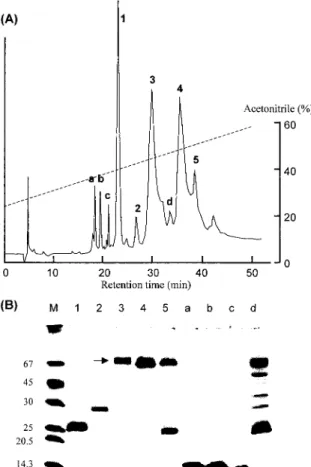
Fig. 3. Purification of P‑60.(A)Reverse‑phase HPLC of P‑60 on a TSK‑GEL ODS‑120T column.
Proteins in peaks 1‑5 and a d were recovered‑ by lyophilization.(B)SDS‑PAGE analysis of the proteins separated by HPLC. M, marker proteins. Arrows indicate the band of HSF.
the abundance of HSF in the fraction E1.25
(Fig. 1B). Most of high molecular mass proteins including several globulins were precipitated until the addition of equal volume of ethanol.
Four fractions (E1.25‑E2.0) contained lo w molecular mass proteins. We repeated the experiment twice and obtained similar results.
Separation of HSF and serum albumin by HPLC:
The main component in P‑60 and E1.25
was serum albumin (Fig. 1). Since serum albumin could not be removed completely by fractional precipitations, the effectiveness of reverse‑phase HPLC was then examined. P‑60
(1.5 mg)was fractionated on a TSK‑GEL ODS‑
120T column, which gave 5 major(1‑5)and 4 minor (a d)peaks as shown in Fig. 3A.‑ Peaks 1‑5 were lyophilized, and their N‑ terminal sequences and inhibitory activities were determined(Table 3). The sequence of peak 1 was very close to that of phospholipase A inhibitor I.2 15)The peak‑3 protein was assumed HSF because it exhibited a strong inhibitory activity toT. flavoviridis metalloprotease, and the sequence was almost identical to that of HSF except for Ser1, which was reported to be Asp.6)The peak‑4 protein is expected to be serum albumin from its molecular mass 68,700(M + H )determined by MALDI‑TOF‑+ MS. Its N terminal sequence showed some‑
Table 3. N‑terminal sequences and the inhibitory activities of some proteins purified by HPLC of P‑60.
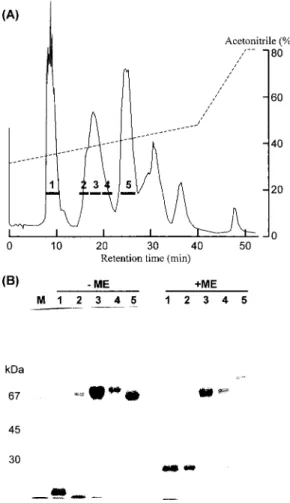
Fig. 4. Purification of HSF.(A)Preparative reverse‑
phase HPLC on a µBondasphere 5µ‑C ‑3008
Å(1.9×15 cm)column.(B)SDS‑PAGE of fractions obtained by HPLC. A 10% gel was used. M, marker proteins. Right five lanes were run in the presence of 2 mercaptoethanol‑
(ME).
homology to cobra serum albumin(HVDDQHS TIRLASQI).16)
SDS‑PAGE analysis of these peaks indicated the complete separation of HSF and serum albumin(lanes 3 and 4 in Fig. 3B). However, HSF obtained has a minor band at about 22kDa.
We found that this minor band protein has a trypsin inhibitory activity and binds to a Blue‑ Sepharose column, which did not bind HSF
(data not shown).
Purification of HSF:
Based on the above results, we purified HSF from the serum of T. flavoviridis. Because the ammonium sulfate precipitation requires desalting, the fractional precipitation by ethanol was firstly employed. The crude serum(100 ml)was fractionated by ethanol precipitation and 888 mg of E1.25protein was obtained. This
(about 200 mg each)was dissolved in 0.1%
TFA and applied to a column of µBondasphere 5µ‑C8‑300Å(1.9×15 cm), and eluted by a linear gradient of acetonitrile as shown in Fig.
4A. The peak 3 containing HSF had a small amount of impurity(below30 kDa)(Fig. 4B). The impurity could be removed by gel filtration on a Sephacryl S‑200HR column (data not shown).
The parameters of each purification of HSF are summarized in Table 4. HSF activity was assayed by brevilysin H6 because it was much more sensitive to HSF than HR2a and could
Table 4. Summary of purification of HSF from T. flavoviridis serum.a)
Fig. 5. Mass spectrum of the purified HSF. Number near the peak shows the measured molecular masses(M + H )by MALDI‑TOF‑MS.+
be obtained easily from the venom of Gloydius halys brevicaudus.17,18)HSF was purified 24‑fold from the crude serum by three steps. The purified HSF showed a single band of 68 kDa on SDS‑PAGE. The molecular mass was 47,810determined by MALDI‑TOF‑MS(Fig. 5). This value was larger than that expected from the amino acid sequence.6)As HSF has three carbohydrate binding sequences,‑ the difference may be due to carbohydrate chains.
No trypsin inhibitory activity was detected in the HSF preparation (data not shown). The purified HSF strongly inhibited brevilysin H6(Fig. 6); H6(20 µg/ml = 4.05×10‑7 M)
for 19 µg/ml(3.97×10‑7 M)of HSF, indicat‑
ing that HSF bound to H6 with 1:1 stoichiomet‑
ry.The dissociation constant,Kd,of the inhibitor‑ enzyme complex was calculated to be 3.7×
10‑9 M by the method of Green and Work.19)
Fig. 6. Inhibition of brevilysin H6 by the purified HSF. The final proteinase concentration was 20 µg/ml(4.05×10‑7 M). Activity was measure at 37℃ in 5 mM CaCl ‑40 mM2
Tris ‑HCl(pH 8.5).‑
Fig. 7. Effect of HSF on hemorrhage induced by crude T. flavoviridis venom. 5 µg of crude venom mixed with various amounts of HSF were injected in three mice for each group.
Incidence of hemorrhage is cited in parenthesis.
The suppressive effect of HSF on hemorrhage induced by 5 µg of crude T. flavoviridis venom was tested in mice. HSF suppressed the hemorrhage dose dependently(Fig. 7)‑ . At 5 µg
of HSF, only one mouse showed the hemorrhage, and no hemorrhage was observed at the dose of more than 10 µg of HSF.
Discussion
High concentrations of salts or water miscible‑ organic solvents has been often used for the fractional precipitation of proteins. Because ammonium sulfate is employed the most frequently for this purpose, we examined the effect of this salt on the purification of HSF. The protein in Habu snake serum precipitated at 45‑60%
saturation of ammonium sulfate. However, the purity of HSF was not high and the incomplete separation from serum albumin was observed
(Fig. 1A). Ethanol precipitation may be much convenient method because it does not require any desalting step after the fractionation. The purity of HSF by this method is slightly higher
than that by the ammonium sulfate method.
A trypsin inhibitor (about 22 kDa) was observed in a HSF preparation obtained by the ammonium sulfate fractionation. Since bound to Blue Sepharose column,‑ this inhibitor can easily be removed by the column. HSF did not bind to Blue Sepharose.‑ On the other hand,the fraction(E1.25)from ethanol precipitation as well as the purified HSF showed no trypsin inhibitory activity, indicating that ethanol precipitation is better than salt precipitation.‑
In the present study, we established a new and rapid method for the purification of HSF from the crude serum of T. flavoviridis.It includes three steps, ethanol precipitation, preparative reverse phase HPLC on a C column,‑ 8 and gel
filtration. Because HSF is quite stable to high temperature and extreme pH, acidic conditions used in HPLC step did not cause any denaturation or degradation of the protein. Practically, HSF purer than 95% could be obtained from the first two steps.
Since hemorrhagins are metalloproteinases, the inhibitors of such enzymes can be used as new drugs to prevent the hemorrhage.3)HSF is thought as the excellent candidate. It actually inhibited the hemorrhage induced byT. flavoviridis venom(Fig. 7).
References
1)G.B.Domont, J.Perales and H.Moussatche:
Toxicon, 29, 1183‑1194(1991).
2)M. M. Thwin and P. Gopalakrishnakone:
Toxicon, 36, 1471‑1482(1998).
3)J. C. Perez and E.´ E. Sanchez:´ Toxicon, 37, 703‑728(1999).
4)J.J.Catanese and L.F. Kress: Biochemistry, 31, 410‑418(1992).
5)J. C. Perez,´ W. Haws, V. Garcia and B. Jennings: Toxicon, 16, 375‑383(1978). 6) Y. Yamakawa and T. Omori Satoh:‑
J. Biochem., 112, 583‑589(1992).
7)K. F. Huang, C. C. Hung, S. H. Wu and S. H. Chiou: Biochem. Biophys. Res.
Commun., 248, 562‑568(1998).
8)R. H. Valente, B. Dragulev, J. Perales,
J. W. Fox and G. B. Domont: Eur.
J. Biochem ., 268, 3042‑3052(2001). 9)T. Omori Satoh,‑ S. Sadahiro, A. Ohsaka
and R. Murata: Biochim. Biophys. Acta, 285, 414‑426(1972).
10) T. Takahashi and A. Ohsaka: Biochim.
Biophys. Acta, 198, 293‑307(1970). 11) T. Takahashi and A. Ohsaka: Biochim.
Biophys. Acta, 207, 65‑75(1970).
12)S. S. Twining: Anal. Biochem., 143, 30‑34
(1984).
13)U.K. Laemmli: Nature, 227, 680‑685(1970). 14)H. Kondo, S. Kondo, H. Ikezawa, R.
Murata and A. Ohsaka, Japan. J. Med. Sci.
Biol., 13, 43‑51(1960).
15)I. Nobuhisa, S. Inamasu, M. Nakai, A.
T atsui, T. M i m ori, T. O ga w a, Y. Shimohigashi, Y. Fukumaki, S. Hattori, H. Kihara and M. Ohno: Eur. J. Biochem., 249, 838‑845(1997).
16)X. Wang, B. Havsteen and H. Hansen:
Biol. Chem. Hoppe Seyler, 376:545‑553(1995)‑ . 17)S. Terada, E. Kimoto,H.Kawasaki,W.‑X.
Hao and H.‑W. Li: Fukuoka Univ. Sci. Rep.
21, 147‑153(1991)
18)S. Fujimura, K. Oshikawa, S. Terada and E. Kimoto: J. Biochem., 128 , 167‑
173(2000).
19)N. M. Green and E. Work: Biochem. J., 54, 347‑352(1953).