近畿大学学術情報リポジトリ
9
0
0
全文
(2) H. Masaki. risk of tumorigenicity, and recurrent autoimmune responses against the transplanted grafts.2 Thus, xenotransplantation is still the method closest to clinical application at present, and it is worthwhile to strive for a comprehensive understanding of the cellular and molecular bases of xeno-reactive immune responses. Apart from hyperacute xenograft rejection, which is mostly due to natural xeno-reactive antibodies, it has been well-established that acute or chronic xenograft rejection is mainly mediated by cellular immunity.3-1° In phylogenetically disparate combinations, such as mouse-human, donor xeno-antigens need to be processed in recipient antigen-presenting cells (APCs) to be recognized by xeno-reactive T cells. This mode of peptide recognition associated with self major histocompatibility complex (MHC) molecules is identical to the recognition of other foreign antigens (i.e., via the indirect recognition pathway). In contrast, it has become evident that donor xeno-antigens can be directly recognized by recipient xeno-reactive T cells as intact MHC molecules on the cell surface (i.e., via the direct recognition pathway). This takes place in phylogenetically close combinations, such as mouse-rat, and is similar to alloreactivity within species.8'11-13 Xeno-reactive CD4 T cells activated via the indirect recognition pathway have been reported to be prominent in xeno-recognition, and play an essential role in xenograft rejection.7'9"0 However, some reports indicated that CD8 T cells, once activated, probably via the direct recognition pathway, are capable of rejecting xenografts by themselves.5'6 Thus, some controversies still exist regarding xeno-recognition and xenograft rejection by T cells. To elucidate the role of indirect-versus-direct recognition in anti-xeno responses and xenograft rejection by xeno-reactive T cells, we established mouse T-cell clones reactive to rat xeno-antigens. We plan to perform adoptive transfer into severe combined immunodeficiency (SCID) mice and establish T-cell receptor (TcR)-transgenic mice to obtain naïve xeno-reactive T cells. The availability of these tools would facilitate the elucidation of the initial stages of anti-xeno responses. In the present report, we first describe the establishment of xeno-reactive T-cell clones recognizing xeno-antigen either via the indirect or direct pathway, to pave the way for these future studies.. 102. Materials. and. Methods. Animals Six- to 9-week-old male C57BL/6 (B6) (H-2b), BALB/c (H-2"), C3H/HeJ (H-2k), and H-2b-congenic BALB.B mice were obtained from the National Cancer Institute, Frederick, MD, USA. Six- to 9-week-old male Wistar Furth (WF) (RT1u), LEWIS (RT1'), and PVG (RT1C) rats were obtained from Harlan Sprague Dawley, Indianapolis, IN, USA. BBDR/Wor and RT1C"congenic PVG.1R8 rats were obtained from the colony at the University of Massachusetts Medical Center, Worcester, MA, USA. They were certified as pathogen-free, and housed in a specific pathogen-free facility until use. They were maintained in accordance with recommendations in the Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, National Academy of Sciences, 1996) and the guidelines of the institutional animal care and use committee of the University of Massachusetts Medical School. Antibodies and flow cytometry Fluorescein isothiocyanate (FITC)-conjugated rat anti-mouse CD8 monoclonal antibody (mAb) (clone 53-6.7) and phycoerythrin (PE)conjugated rat anti-mouse CD4 mAb (clone RM4.5) were obtained from BD PharMingen (San Diego, CA, USA). Flow cytometry was performed as previously described.14 Briefly, 1x106 viable cloned T cells were incubated with a mixture of the above FITC- and PE-conjugated mAbs (1 1ag each) for 30 min at 4°C. Thereafter, cells were washed extensively with Dulbecco's phosphate-buffered saline (PBS) containing 0.1% fetal calf serum (FCS) and 0.02% NaN3 (FACS medium), and then fixed with 1% paraformaldehyde in FACS medium. FITC- and PE-conjugated isotype control Igs (rat IgGza, x) were also obtained from BD PharMingen, and used for all analyses. Cells were analyzed using a Becton Dickinson FACS® instrument (Becton Dickinson, San Jose, CA, USA). The lymphocyte fraction was gated for viable cells on the basis of forward and side scatter. WinList 4.0 software (Verity Software, Topsham, MA, USA) was used for data analysis. For the blockade of T-cell proliferative responses, rat anti-mouse CD4 mAb (clone RM4.5), rat anti-mouse CD8 mAb (clone 53-6.7), mouse anti-rat RT1A mAb (clone OX-l8),.
(3) Xeno-reactive. mouse anti-rat RT 1B mAb (clone OX-6), and mouse anti-rat RT1D mAb (clone OX-17) were obtained from BD PharMingen. The preservative NaN3 was removed by passing through a desalting pre-packed gel filtration column (PD10, GE Healthcare Bio-Science, Piscataway, NJ, USA) in accordance with the manufacturer's instructions. The antibodies were then added to the cultures. Preparation of rat T-cell growth factor (TCGF) Spleens were collected from 10-week-old male BBDR/Wor rats, minced to yield single-cell suspensions, and erythrocytes were lysed by treatment with 155 mM NH4C1 hemolytic solution. The remaining cells were washed with RPM] 1640 medium, and suspended at lx107/m1 in EHAA medium (Invitrogen, Carlsbad, CA, USA) supplemented with 2 mM L-glutamine, 5x10-5 M 2-mercaptoethanol (2-ME), 100 U/ml penicillin, 100 ,ug/ml streptomycin, 10 mM HEPES, and 10% heat-inactivated FCS (Complete medium 1) containing 5 pg/ml of concanavalin A (Sigma, St. Louis, MO, USA, Cat.# : C-5275). They were cultured at 37°C in 5% CO2 for 48 hrs. Culture supernatants were harvested by centrifugation at 1,500 G for 10 min, filtered through 0.45-pm-pore size membranes, and stored at —70°C until use. Generation of B6 mouse anti-WF rat T-cell lines and clones B6 mouse anti-WF rat T-cell lines were generated from anti -WF rat mixed lymphocyte reaction (MLR) cultures after in vivo immunization with WF rat cells. Xeno-reactive T-cell clones were established from the T-cell lines using the limiting dilution method. Briefly, B6 mice were immunized with 3x107 WF rat splenocytes intraperitoneally. Four weeks later, spleens were collected, pooled, and minced to yield single-cell suspensions, and 3x107 of the B6 splenocytes were co-cultured with the same number of 33-Gy 7-irradiated WF rat thymocytes in 20 ml of Complete medium 1 at 37°C in 5% CO2. Irradiation (^-184c Gy/min) was carried out using a 137Cssource (Gammacell 40 ; Atomic Energy of Canada, Ottawa, ON, Canada). On day 14, viable cells were harvested using a density gradient method on Lympholite-M© (Cedarlane, Hornby, ON, Canada), and 6x105 cells were cocultured with 1.5x107 irradiated syngeneic. T-cell. clones. splenocytes as APCs and the same number of irradiated WF rat thymocytes as sources of xenoantigen in 6 ml of Complete medium 1 for 7 days. Viable cells were then harvested and restimulated under the same conditions every week. After 4 restimulation cycles, viable cells were seeded into 96-well flat-bottomed plates containing 5x105 syngeneic APCs and the same number of irradiated WF rat thymocytes suspended in 150pl of Complete medium 1 supplemented with 20% TCGF at a dilution of 0.3 cells / well. Five days later, 100 p 1 of fresh Complete medium 1 containing 20% TCGF was added to each well. On day 14, growing cells were screened for their specificity using the proliferation assay described below. Antigenspecific cells were expanded by stimulating with APCs (2.5x106/ml) and irradiated WF rat thymocytes (2.5x106/ml) in Complete medium 1 containing 10% TCGF every 7 days. Once expanded, clones were maintained by stimulating with APCs (2.5x106/ml) and the same number of irradiated WF rat thymocytes in Complete medium 1 containing 2% TCGF approximately every 1-2 weeks. Using this protocol, we obtained only CD8-positive clones (CD8 clones). To establish CD4-positive clones (CD4 clones), the same method was applied except that B6 mice were immunized with WF rat thymocytes instead of splenocytes, and cultures were carried out in the presence of anti-mouse CD8 mAb (10 pg/m1) for two restimulation cycles before initiating limiting dilution culture and cloning, in order to inhibit the proliferation of CD8-positive cells. Proliferation assay Ten thousand cells were cultured with or without irradiated rat thymocytes (5x105) in the presence or absence of APCs (5x105) in 200,u1 of Complete medium 1 in 96-well round-bottomed plates at 37°C in 5% CO2. For CD8-positive cell proliferation assays, the culture medium was supplemented with 2% TCGF. After 52 hrs, proliferation was quantified by pulsing with 1 pCi of [311-thymidine (New England Nuclear, Boston, MA, USA) for 24 hrs. Cells were harvested onto glass fiber filter paper, and nuclear radioactivity was quantified by liquid scintillation counting (1450 Microbeta Trilux, Wallac, Gaithersburg, MD, USA). Assays were performed in triplicate, and the data are expressed as the mean value ± standard deviation (SD) of 103.
(4) H. Masaki. counts per minute (cpm) except for specificity screening of limiting dilution culture, in which stimulation indices (Si) (cpm with irradiated WF rat thymocytes/cpm without those) were evaluated.. release.. sed as the mean. ±. SD of % specific. lysis.. Results. We selected 4 clones (2C5, 3B6, 6C5, and 6G4) with the highest SI (i.e., greatest specific proliferation in response to WF rat thymocytes, relative to the lower background in the absence of specific stimulation) and propagated them for further study. Cells from these clones were stained with FITC-conjugated anti-mouse CD8 mAb and PE-conjugated anti-mouse CD4 mAb to determine their phenotypes. As shown in Fig. IA, all were CD8-single positive (i.e., CD8 clones). It is unclear whether or not we obtained only CD8 clones and no CD4 clones by chance. Therefore, we set out to generate CD4 clones by. B). 2C5. 7E11. 3B6. -. 6C5 '. 8E38G4. 6G4. D. CD8 ------------------------------•. 104. are expres-. Statistical analysis The statistical significance was determined by Student's t-test using StatFlex® version 6 software (Artech, Osaka, Japan), and P<0.05 was considered to be statistically significant.. Fig.. o ~. value. Data. Quantification of cytokine production Cytokine production by CD4 clones responding to WF rat xeno-antigen was analyzed using enzyme-linked immunosorbent assay (ELISA) cytokine quantification kits. CD4 clone cells (1x105) were stimulated with irradiated syngeneic splenocytes (2.5x106) and irradiated WF rat thymocytes (2.5x106) in 2 ml of Complete medium 1 in 24-well plates at 37°C in 5% CO2. Supernatants were collected at 24, 48, and 72 hrs. Concentrations of interleukins 2, 4, and 10 (IL2, IL-4, and IL-10) and interferon y (IFN-y) were measured using R&D Systems mouse cytokine ELISA kits, Quantikine M (M2000, M4000, M1000, and MIF00, R&D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions.. Cytotoxicity assay Cytotoxic activity of CD8 clones against NKresistant C58NT(D) lymphoma cells of WF rat origin15,16 was measured using 51Cr-release assays. C58NT(D) cells were obtained from the American Type Culture Collection (Manassas, VA, USA), and were maintained in RPMI 1640 medium (Invitrogen) supplemented with 5x10-5 M 2-ME, 100 U/ml penicillin, 100 jig/m1 streptomycin, 10 mM HEPES, and 10% FCS (Complete medium 2) at 37°C in 5% CO2. C58NT(D) cells (1x106) in the growing phase were labeled with 100 j Ci of Na251CrO4 (New England Nuclear) for 1 hr, and then washed extensively with RPMI 1640 medium. 51Cr-labeled C58NT(D) cells were suspended in Complete medium 2, and seeded in 96-well V-bottom plates at lx104 cells per well. CD8 clone cells were added to the plates to give effector/target ratios (E/T ratios) ranging from 0.2 to 5 : 1 in a total volume of 0.2 ml per well, and the plate was incubated at 37°C in 5% CO2. Four hrs later, 100-,ul aliquots of the supernatant were transferred to a separate 96-well plate containing 100 ,i l Optiphase Supermix f-scintillation fluid (Wallac), and the 51Cr content was measured with a 1450 Microbeta Trilux instrument (Wallac). Maximum 51Crrelease was assessed by adding 2% Triton X-100, and spontaneous 51Crrelease was determined from wells that contained target cells and medium only. Assays were performed in triplicate, and the percent specific lysis was calculated by the formula :% specific lysis=100 x [(release with effector cells —spontaneous release)/(maximum release—spontaneous release)]. Spontaneous release did not exceed A). 20.6% of the maximum. ;. i U. CD8. 8B2. 1. Flow cytometric analysis of surface markers on xeno-reactive T-cell clones. Cells were stained with FTTC-conjugated anti-mouse CD8 mAb and PE-conjugated anti-mouse CD4 mAb, and subjected to flow cytometric analysis, as described in Materials and Methods. Clones 2C5, 3B6, 6C5, and 6G4 are CD8-positive (A), whereas clones 7E11, 8B2, 8E3, and 8G4 are CD4-positive (B)..
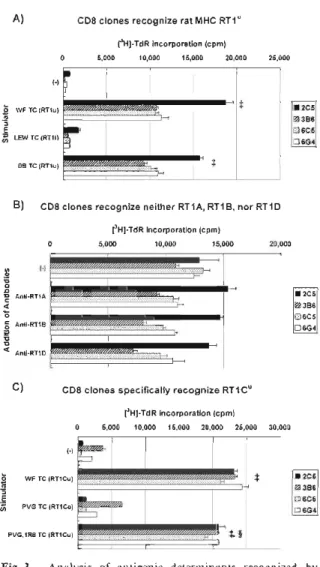
(5) Xeno-reactive. adding anti-CD8 mAb to the cultures to inhibit proliferation of the CD8-positive cells. Four clones, 7E11, 8B2, 8E3, and 8G4, with the best SI, were selected and phenotyped. In this case, all were CD4 single positive, with no CD8 clones present (Fig. 1B). The addition of anti-CD8 but not CD4 mAb to the cultures inhibited the specific proliferation of clones 2C5, 3B6, 6C5, and 6G4 in response to WF rat thymocytes, whereas anti-CD4 but not CD8 mAb, inhibited the specific proliferation of clones 7E11, 8B2, 8E3, and 8G4 (data not shown). Thus, clones 2C5, 3B6, 6C5, and 6G4 possess functional CD8 but not CD4 molecules, whereas clones 7E11, 8B2, 8E3, and 8G4 possess functional CD4 but not CD8 molecules. We also confirmed that all clones were positive for Thy1.2, CD3, and the TcR f chain, but did not express 76-TcR (data not shown), indicating that they are typical a/6T cells. It has been reported that there are two pathways for the recognition of xeno-antigens by xeno-reactive T cells, namely direct and indirect. Therefore, we investigated by which pathway our xeno-reactive T-cell clones recognized rat xenoantigens. As shown in Fig. 2A, CD8 clones 2C5, 3B6, 6C5, and 6G4 all proliferated in response to WF rat thymocytes alone to the same extent as to WF rat thymocytes mixed with syngeneic B6 mouse splenocytes. In contrast, all four CD4 clones proliferated in response to WF rat thymocytes only when syngeneic splenocytes were also present (Fig. 2B). These results indicate that all CD8 clones recognize WF rat xenoantigen via the direct pathway, while all CD4 clones exclusively use the indirect recognition pathway. We also observed that the CD8 clones responded to allogeneic mouse splenocytes, especially to C3H/HeJ cells which possess the H-2k haplotype (data not shown). Of the CD4 clones, 8B2 in particular proliferated significantly in response to C3H/HeJ splenocytes as in direct recognition (see Fig. 5 legend). This suggests that these clones possessed a heterogeneous repertoire, and that the ontogeny of (direct) xenorecognition may be cross-reaction to allo-recognition. We next analyzed the nature of the xeno-MHC molecules recognized by our CD8 clones. They all proliferated significantly when stimulated with RT1u haplotype WF or BBDR/Wor rat thymocytes, but not with RT11 haplotype LEWIS cells (Fig. 3A). Therefore, they recog-. T-cell. clones. A). CD8 clones recognize xeno-antigen via direct recognition pathway. 0. 5,000. [31]-TdR incorporation (cpm) 10,000 15,000 20,000. 25,000. i-1. a. • 2C5 ®3B6. 86 SC. E. ^6C5 ^ 6G4. WF TC. 86 S C + WF TC. B). CD4 clones recognize xeno-antigen via indirect recognition pathway. 05,000. (3H]-TdRincorporation (cpm) 10,000 15,000 20,000. 25,000. (-) a. B6 SC. ^ 7E11 08B2. e. 8E3 WF TC. • 8G4. 86 SC+ W F TC. Fig. 2. Proliferative response of xeno-reactive T-cell clones against WF rat thymocytes with or without syngeneic B6 mouse splenocytes. Proliferation was evaluated as [3H]-thymidine incorporation, as described in Materials and Methods. (-), none ; B6 SC, B6 mouse splenocytes ; WF TC, WF rat thymocytes ; [3H]-TdR ; [3H] -thymidine. $ P < 0.05 compared with (-), § P < 0.05 compared with B6 SC.. nize haplotype u of the RT1 complex, presumably one of the MHC class I loci because they were CD8-positive. We subsequently examined whether anti-MHC class I antibody anti-RT1A or anti-class II antibodies anti-RT 1B and antiRT1D inhibited the proliferation of these clones. As shown in Fig. 3B, none of them could significantly inhibit proliferation. It was therefore concluded that neither RT1A, RT1B, nor RT1D was recognized by these CD8 clones. We further tested whether they proliferated in response to PVG or PVG.1R8 rat thymocytes. As shown in Fig. 3C, they did not respond to PVG, which possesses RT 1Eu, RT 1Gc, and RT 1Cc, but did respond to PVG.1R8, which possesses RT1Eu and RT1CU. They also responded to WF, which possesses RT1EU, RT1Gc, and RT1CU (Table l).'' Taken together, these results strongly suggest that all CD8 clones specifically recognize the rat MHC class I RT I C1'-allele via the direct recognition pathway. CD8 is a marker of cytotoxic T lymphocytes (CTLs). Thus, we tested whether the CD8 clones 2C5, 3B6, 6C5, and 6G4 were cytotoxic to WF 105.
(6) H. Masaki. A). CD8 clones. recognize. [3H]-TdRincorporation (cpm) 10,000 15,000 20,000. 05,000. Table. rat MHC RT1". 1. RTI. alleles. CD8. clones. 25,000. of rat. strains. RT 1 complext ABDE • 2C5. WF TC (RT1u). 3B6. E. CD8 clones. recognize. neither. RT1A,. (3H1-TdR incorporation. RT1B,. nor. RT1D. 20,000. U. U. U. u. c. u. Yes. BB. U. U. u. u. NU. ND. Yes. LEW. 1. 1. 1. c. c. c. u. 1 c. No. PVG. a c. a. u. u. u. tRT 1 complex H.. O • 2C5. Anti-RT1A. 3B6. 4 O. 6C5 Anti-RT1 B. 06G. and. and. CD8. clones. specifically. recognize. [3H]-TdR incorporation 05,000. 10,000. 15,000. MHC. clone. §ND , not. RT1Cu. Commonly. class. I loci,. Yes. u. is cited Used. and. from. Rat. in Immunology.'7. response 3A. Strains. RT1.A, RT1.B. Kunz. and. in. E, G, D are. and. is based. on the. findings. shown. 3C.. determined.. 25,000. CD8 clones. 30,000. lyse C58NT(D) cells. 70. ................................................... 60. t I. • 2C5. WF TC (RT1Cuj. 03B6. E Cl,. ND. rat strain. (cpm) 20,000. (-)iororiri I O. No. II loci.. in Figures CD8. T.,. Protocols. C are. class. of each. Gill. Current. Anti-RT1 D. C). responses. WF. PVG.1R8. 15,000. (-). 4. C. Congenic. (cpm). 05,00010,000. O -o. G. ^6G4. BB TC (RT1 u). N d. clone. Inbred. t26C5 LEW TC (RT11). B). CD8. of. Strain. (-). a. and response. 0 6C5 PVG TC (RT1CC). 50. Vz1P'/. u. 06G. 40 0. t. 2C5. f. 3B6. —~6C5 PVG.1R8. TC (RT1Cuj. $, §. U. 30 ~6G4 20. Fig. 3. Analysis of antigenic determinants recognized by CD8 clones. Proliferation was evaluated as [3H]-thymidine incorporation, as described in Materials and Methods. A) Proliferative responses against different strains of rat thymocytes without APCs (syngeneic splenocytes) (i. e., direct recognition). (-), none ; WF TC, WF rat thymocytes ; LEW, LEWIS rat ; BB, BBDR/Wor rat. The RT1 haplotype of each strain is given in parentheses. $ P<0.05 compared with (-). B) Proliferative responses against WF rat thymocytes without APCs (i.e., direct recognition) in the presence of anti-RTl antibody (10,ug/m1 in culture). None of the antibodies tested significantly inhibited proliferation. C) Proliferative responses against WF, PVG, or PVG.1R8 rat thymocytes without APCs (i. e., direct recognition). Alleles of the RT1C locus are given in parentheses. I P <0.05 compared with (-), § P <0.05 compared with PVG TC.. rat-derived cells. As shown in Fig. 4, all CD8 clones dose-dependently lysed C58NT(D) cells, which are NK-resistant lymphoma cells of WF rat origin. These results suggest that the CD8 clones indeed possess functional CTL characteristics. 106. 10. 0 0.21. 5 E / T ratio. Fig. 4. Cytotoxicity of CD8 clones against C58NT(D) cells. Cells of each CD8 clone were co-incubated with "Crlabeled C58NT(D) cells for 4 hours, and the % cytotoxicity was evaluated, as described in Materials and Methods. E/T ratio, effector cell (i.e., CD8 clone cells)/target cell (i.e., C58NT(D) cells) ratio. CD8 clone cells lysed C58NT(D) cells in a dose-dependent manner. Note that C58NT(D) are NK-resistant lymphoma cells of WF rat origin. $ P<0.05 compared with E/T ratio 0.2.. Next, we analyzed the MHC restriction of the CD4 clones 7E11, 8B2, 8E3, and 8G4 which recognized WF rat xeno-antigens via the indirect pathway. They proliferated in response to WF rat thymocytes only when they were co-cultured with B6 or BALB.B mouse splenocytes as APCs, both of which possess the H-2b haplotype. In contrast, splenocytes from BALB/c (H-2d haplotype) or C3H/HeJ (H-2k haplotype) mice did not support their specific proliferation (Fig. 5A)..
(7) Xeno-reactive. These. results. indicate. that. recognize. rat xeno-antigens. manner.. We. A). further. all. 4. CD4. clones. in an H-2b-restricted. investigated. whether. rat. CD4clonesrecognize xeno-antigens in H-2b- restrictedmanner 05,000. [3H]-1]incorporation (Acpm) 10,000 15,000. 20,000. 0.B6SC (H-2b)yiyi~iyi<iyiiyiiiriii~ri~iiri~rii.t, § oli ~~2 SC (H-2d)/ •BALBIc ^7E11 ^ 8B2 218E3 8G4. a •C3H SC(H-2k)~• dl~ O BALB.B SC(H-2b). CD4clones recognize rat xeno-antigens shared between different strains. B). 0. [3H]-TdRincorporation (cpm) 5,000 10,000 15,000 20,000. 25,000. T-cell. clones. xeno-antigenic determinants recognized by these clones were derived from any specific RT1 haplotype. As shown in Fig. 5B, all CD4 clones proliferated essentially to a similar degree against different strains of rat thymocytes not sharing the same RT1 haplotype. Taken together, these results indicate that all CD4 clones recognize rat xeno-antigens in thymocytes shared between different strains but in the context of the mouse MHC H-2b haplotype. CD4 is a marker of helper T cells (Th), which classically can be categorized into either Th 1 or Th2 cells according to their cytokine production pattern. We therefore assayed the cytokines secreted by these CD4 clones in response to rat xeno-antigens. As shown in Table 2, they mainly secreted IFN-y but very little of either IL-4 or IL-10, indicating that they all possessed a hallmark characteristic of Thl cells.. (-) o. o. Discussion. wF TC (RT1u) WF ^ 7E11 O 8B2. LE \A TC (RT11) LEW. o 25. ® 8E3 • 8G4. PVG TC (RT1c). F PVG. Y BB BBTC (RT1u) Fig. 5. CD4 between. clones. recognize. different. strains. rat of rats. xeno-antigens. shared. in an H-2b-restricted. manner.. The proliferative response was evaluated as PH] thymidine incorporation, as described in Materials and Methods. A) Proliferative responses against WF rat thymocytes in the presence of mouse splenocytes of different strains as APCs (i.e., indirect recognition). Data are expressed as Acpm, which is the difference in the cpm value between responses in cultures with WF rat thymocytes and those without (i.e., background response). B6 SC, B6 mouse splenocytes ; BALB/c, BALB/c mouse ; C3H, C3H/ HeJ mouse ; BALB.B, BALB.B mouse. Background responses were as follows : clone 7E11 ; B6 SC : 88+12, BALB/c SC : 403+87, C3H SC : 344+58, BALB.B SC : 379+70, clone 8B2 ; B6 SC : 125+25, BALB/c SC : 4,813+243, C3H SC : 8,950+693, BALB.B SC : 1,455+143, clone 8E3 ; B6 SC : 131+ 18, BALB/c SC : 677+357, C3H SC : 477+79, BALB.B SC : 832+158, clone 8G4 ; B6 SC : 405+ 164, BALB/c SC : 2,367+590, C3H SC : 1,643+ 168, BALB.B SC : 2,992+325. $ P<0.05 compared with BALB/c SC, § P<0.05 compared with C3H SC. B) Proliferative responses against rat thymocytes of different strains in the presence of syngeneic B6 mouse splenocytes as APCs (i.e., indirect recognition). (-), none ; WF TC, WF rat thymocytes ; LEW, LEWTS rat ; PVG, PVG rat BB, BBDR/Wor rat. $ P < 0.05 compared with (-), except that P < 0.0595 with clone 7E11.. We have isolated and characterized four CD8 and four CD4 T-cell clones of B6 mouse origin reactive to WF rat xeno-antigens. Hirota et al. also established CD4 clones as well as CD8 clones of B6 mouse origin specifically reactive to Fischer 344 (F344) (haplotype lvl), but not to WKAH rat cells.1s_21 Those CD8 clones recognized MHC class 1 RT 1A molecules via the direct recognition pathway, and specifically Table 2. Clone. Cytokine production of CD4 clones in response to rat xeno-antigens Concentration. Culture time (hr). (pg/ml). TL-2. TFN-y. IL-4. 24. 0. 7,350. 81. 81. 48. 23. 6,157. 23. 140. 72. 40. 5,074. 93. 146. 24. 0. 58,140. 123. 177. 48. 0. 59,420. 0. 200. 72. 23. 57,160. 17. 399. 24. 0. 14,590. 0. 117. 48. 44. 12,300. 0. 115. 72. 0. 9,703. 0. 71. 24. 0. 20,040. 14. 46. 48. 0. 27,840. 26. 28. 72. 19. 24,140. 98. 12. Cytokine. concentration. in. each. point. 7E11. 8B2. 8E3. 8G4. time. quantification and. was. measured. ELISA. kit. culture using. as described. IL-l0. supernatant mouse. at. cytokine. in Materials. Methods.. 107.
(8) H. Masaki. lysed F344 rat lymphoblasts, indicating that they were CTLs, similar to ours. Their CD4 clones, however, recognized F344 rat MHC class II molecules (RTIB or RTID) also via the direct pathway, and some mainly secreted IL-4, suggesting that they possessed Th2 characteristics. The reason that we did not obtain CD4 clones recognizing MHC class II molecules via the direct pathway or clones possessing Th2 characteristics is unclear. One possibility is that, if we had simply picked up more CD4 clones, we might have obtained some of this type too, responding to MHC class II molecules via the direct recognition pathway or possessing Th2 characteristics. Another possibility is that our clones were derived from a secondary MLR culture, in which responder cells were splenocytes from B6 mice immunized with WF rat thymocytes, and thymocytes were always used as a source of xeno-antigen in the presence of syngeneic splenocytes. This is a different protocol from that of Hirota et al. ; their CD4 clones were derived from primary MLR cultures, and only splenocytes of F344 rats were used as stimulator cells. The different conditions for establishing our CD4 clones might have diverted the xeno-reactive CD4 T-cell repertoire towards indirect recognition, and polarization to Thl. In the human-pig combination, Shishido et al. reported human CD8 clones which recognized S LAdd MHC class I via the direct pathway, as well as human CD4 clones that recognized pig xeno-antigens via the indirect pathway,22,23similar to our clones. In contrast, however, their CD4 clones responded only to SLAdd pig xenoantigen strain-specifically,23 suggesting that they recognized peptides derived from the polymorphic region of pig MHC class I or class II, associated with self-HLA class II molecules. Murphy et al. also described CD4 T cells specifically recognizing synthetic peptides spanning the polymorphic region of rat MHC class II molecules identical to those of the xenograft. These cells used the indirect recognition pathway, and had been induced in B10.A mice after the rejection of xenogeneic WF rat skin or heart grafts.4 In contrast, our CD4 clones recognize rat xenoantigens shared between several different strains, suggesting that the peptides recognized were derived from thymocyte-specific rat proteins, tissue non-specific proteins, or non-polymorphic regions of rat MHC molecules, which may behave like minor histocompatibility antigens. 108. Fine specificity analysis of our CD4 clones and determination of their epitopes needs to be addressed in the future. For the CD8 clones, we have analyzed their fine specificities, and determined them to be specific for the RTICu-allele. This is the first report elucidating the fine specificity of mouse CD8 clones that recognize rat MHC via the direct pathway. Because our CD8 clones were cytotoxic to WF rat cells, and because our CD4 clones produced large amounts of the proinflammatory cytokine IFN-y in response to rat xeno-antigens, it is likely that they would act as effector cells in vivo for the rejection of WF tissues or organs transplanted into B6 mice. The adoptive transfer of CD8 clones, CD4 clones, or both, into B6-SCID mice transplanted with WF tissues may facilitate the functional dissection of xeno-reactive CD8 T-cell direct recognition and CD4 T-cell indirect recognition mediating xenograft rejection, and, hence, the development of specific intervention strategies to control anti-xeno responses. Our Tcell clones can be further used as sources of TcR genes to establish xeno-reactive TcR-transgenic mice which will allow us to obtain naïve xenoreactive T cells, expected to be useful for studying the initiation of anti-xeno responses. Our xeno-reactive T-cell clones and methodology for establishing them will contribute to a more comprehensive understanding of xenotransplantation immunity. Acknowledgments Supported. in part by a grant. Diabetes Foundation, tents of this publication bility. of the author,. financial. and. or commercial. from the Juvenile. International. are solely the. author. conflict. The conthe responsideclares. no. of interest.. References. 1. Auchincloss H Jr., Sachs DH (1998) Xenogeneic transplantation. Annu Rev Immunol 16 : 433-470 2. Noguchi H (2007) Stem cells for the treatment of diabetes. Endocr J 54: 7-16 3. Yu PB, Holzknecht ZE, Bruno D, Parker W, Platt JL (1996) Modulation of natural IgM binding and complement activation by natural IgG antibodies: a role for IgG anti-Gal alphal-3 Gal antibodies. J Immunol 157 : 5163-5168 4. Murphy B, Auchincloss H, Jr., Carpenter CB, Sayegh MH (1996) T cell recognition of xeno-MHC peptides during concordant xenograft rejection. Transplanta-.
(9) Xeno-reactive. tion 61 : 1133-1137 5. Uchida T, et al. (1999) Roles of CD4+ and CD8 T cells in discordant skin xenograft rejection. Transplantation 68 : 1721-1727 6. Yin DP, et al. (2002) Role of CD4 and CD8 T cells in the rejection of concordant pancreas xenografts. Transplantation 74 : 1236-1241 7. Olack BJ, et al. (2002) Rejection of porcine islet xenografts mediated by CD4+ T cells activated through the indirect antigen recognition pathway. Xenotrans-. T-cell. clones. tion 13 : 224-232 15. Bernstein ID, Wright PW, Cohen E (1976) Generation of cytotoxic lymphocytes in vitro : response of immune rat spleen cells to a syngeneic gross virusinduced lymphoma in mixed lymphocyte-tumor culture. J Immunol 116 : 1367-1372 16. Reynolds CW, Bere EW Jr., Ward JM (1984) Natural killer activity in the rat. III. Characterization of trans-. production are dependent on CD4+ helper T cells and self antigen-presenting cells in the mouse. J Exp Med 172: 567-575 12. Watschinger B (1995) How T cells recognize alloantigen: evidence for two pathways of allorecognition. Nephrol Dial Transplant 10: 1556-1558 13. Benichou G, Valujskikh A, Heeger PS (1999) Contributions of direct and indirect T cell alloreactivity during allograft rejection in mice. J Immunol 162 : 352 -358. plantable large granular lymphocyte (LGL) leukemias in the F344 rat. J Immunol 132 : 534-540 17. Kunz HW, Gill TJ 3rd. (1996) Commonly used rat strains, In : Coligan JE, et al. (eds) : Current Protocols in Immunology, supplement 17. Hoboken, NJ, John Wiley & Sons, pp A.1 H. l -A.1 H.2 18. Hirota T, et al. (1996) Evidence of direct recognition subset in xeno-reactive helper T cells. Artif Organs 20 : 890-894 19. Hirota T, et al. (1997) Mouse CD4 and CD8 T-cell clones that recognize xeno-antigens directly on xenoantigen presenting cells in xeno MHC-restricted ways. Transplant Proc 29: 950-952 20. Hirota T, et al. (1997) Direct recognition of rat MHC antigens on rat antigen-presenting cells by mouse CD4+ and CD8+ T cells and establishment of T cell clones exhibiting a direct recognition pathway. Transplantation 63 : 705-710 21. Iida Y, et al. (2003) Helper T-cell type 1 or type 2 function of xeno-MHC-restricted T-cell clones in a direct xenoantigen recognition. Transpl Immunol 11 : 169-173 22. Shishido S, Naziruddin B, Howard T, Mohanakumar T (1997) Recognition of porcine major histocompatibility complex class I antigens by human CD8+ cytolytic T cell clones. Transplantation 64 : 340 -346. 14. Masaki H, et al. (2006) Anti-mouse CD154 antibody treatment facilitates generation of mixed xenogeneic rat hematopoietic chimerism, prevents wasting disease and. 23. Shishido S, Naziruddin B. Xu XC, Howard T. Mohanakumar T (1998) Indirect recognition of porcine xenoantigens by human CD4+ T cell clones. Trans-. plantation 9 : 393-401 8. Rayat GR, et al. (2003) The degree of phylogenetic disparity of islet grafts dictates the reliance on indirect CD4 T-cell antigen recognition for rejection. Diabetes 52 : 1433-1440 9. Koulmanda M, Laufer TM, Auchincloss H Jr., Smith RN (2004) Prolonged survival of fetal pig islet xenografts in mice lacking the capacity for an indirect response. Xenotransplantation 11 : 525-530 10. Singh NP, Guo L, Que X, Shirwan H (2004) Blockade of indirect recognition mediated by CD4+ T cells leads to prolonged cardiac xenograft survival. Xenotransplantation 11 : 33-42 11. Moses RD, Pierson RN 3rd, Winn HJ, Auchincloss H Jr. (1990) Xenogeneic proliferation and lymphokine. prolongs xenograft survival in mice. Xenotransplanta-. plantation. 65 : 706-712. 109.
(10)
図
関連したドキュメント
Projection of Differential Algebras and Elimination As was indicated in 5.23, Proposition 5.22 ensures that if we know how to resolve simple basic objects, then a sequence of
Keywords: continuous time random walk, Brownian motion, collision time, skew Young tableaux, tandem queue.. AMS 2000 Subject Classification: Primary:
The response of bodies to external stimuli is characterized by the many ways in which bodies store energy, how they release this energy that is stored, the various ways in which
This paper presents an investigation into the mechanics of this specific problem and develops an analytical approach that accounts for the effects of geometrical and material data on
Abstract. The backward heat problem is known to be ill possed, which has lead to the design of several regularization methods. In this article we apply the method of filtering out
While conducting an experiment regarding fetal move- ments as a result of Pulsed Wave Doppler (PWD) ultrasound, [8] we encountered the severe artifacts in the acquired image2.
L. It is shown that the right-sided, left-sided, and symmetric maximal functions of any measurable function can be integrable only simultaneously. The analogous statement is proved
But in fact we can very quickly bound the axial elbows by the simple center-line method and so, in the vanilla algorithm, we will work only with upper bounds on the axial elbows..
