東京湾内湾に造成された砂浜海岸と泥質干潟の魚類
相と多様性
著者
丸山 啓太, 河野 博, 竹山 佳奈, 中瀬 浩太
雑誌名
東京海洋大学研究報告
巻
17
ページ
1-17
発行年
2021-03-01
URL
http://id.nii.ac.jp/1342/00002035/
[論文]
東京湾内湾に造成された砂浜海岸と泥質干潟の魚類相と多様性
*1
丸山啓太
*2・河野 博
*3・竹山佳奈
*4・中瀬浩太
*4(Accepted November 26, 2020)
Fish Assemblages and Diversities in the Artificial Sandy Beach and Tidal Mud Flat in the
Inner Tokyo Bay, Central Japan
Keita MARUYAMA*2, Hiroshi KOHNO*3, Kana TAKEYAMA*4, Kota NAKASE*4
Abstract: The Furuhama Park, which was artificially improved and developed in 2000 at Heiwajima artificial
island of Ota City, is located at the innermost part of Tokyo Bay. The Park is composed of the sandy beach (1.2 ha) and tidal flat (1.0 ha) with a shallow water area (4.6 ha) between them, and separated from the outside canal by submerged dikes. Monthly samplings of fishes were carried out in both the sandy beach and tidal flat by using the small seine-net from May 2014 to April 2019. A total of 60,050 individuals of 40 species of 19 families and 44,807 individuals of 38 species of 19 families were collected from the sandy beach and tidal flat, respectively. Despite a difference was confirmed in the bottom sediment between the sandy beach and tidal flat, no remarkable differences were detected on the fish fauna. Comparisons with previous studies conducted at natural habitats in the inner Tokyo Bay revealed that the fish diversity of the Park were inferior to natural tidal flats. Our study suggests that the Park provides various habitats for estuarine gobiids, but the location of the Park secluded from the open sea through canals is likely to be unsuitable for marine fish habitats.
Key words: larvae, juvenile, fish fauna, tidal flat, Tokyo Bay
はじめに
明治時代後期までは東京湾内湾(房総半島の富津と三浦 半島の観音崎を結んだ線よりも北の海域)の沿岸域には連 続的な干潟や浅場が存在しており、有用な漁場であった1)。 しかし、1950 年代に入ると内湾への汚染負荷が増え始めて 水質問題が深刻化し、さらに 1960 年代より多くの干潟や 浅場が埋め立てられることにより、魚介類の減少が見られ るようになった2)。とくに埋め立てによる干潟と浅場の減 少は著しく、戦前の干潟の総面積は136 km2とされていた が、1973 年には 10 km2にまで減少した3)。このような状況 の中で、1970 年代より積極的に干潟や浅場を造成しようと する動きが見られるようになり4)、1997 年における干潟の 面積は16.4 km2と増加している5)。 失われた干潟や浅場などの浅い海域は魚類にとって重 要な生息場所として知られ,現在では東京湾内湾でも数少 ない干潟がその機能を担っていることが知られている6,7)。 東京湾内湾の各所では干潟や浅場での調査6,7,8)に加え、東 京都の葛西臨海公園の人工渚 9,10)、神奈川県の八景島海の 公園10)、千葉県の新浜湖11)などの人工の干潟や海浜、潟湖 でも調査が行われてきた。内湾の魚類相をまとめた河野12) は、失われた浅場や干潟域、あるいは海浜の再生計画を進 めていくうえで、人工的な環境が魚類にどのような場を提 供するのかを明らかにする必要性を指摘している。また、 東京都大田区の京浜島の干潟で魚類相を調査した茂木ら 13)は、人工物造成後の仔稚魚への影響を知るためには、1 年といった短期間ではなく、長期間のモニタリングが必要 であるとしている。 干潟や浅場を造成する事業が進められる中、2007 年に東 京都大田区に大森ふるさとの浜辺公園が開園した。本公園 は行政と地元のステークホルダーが20 年にわたる議論の すえに造成された、砂浜海岸(1.2 ha)と泥質干潟(1.0 ha)、 浅場(4.6 ha)を有する臨海公園である。周辺の深さ 4 m の運河域では、夏場の水深 2 m 以深で溶存酸素濃度が 3 mg/L 以下と、貧酸素化する14)。モニタリング調査として は、施工前より5 年以上ベントス、水質および底質の調査 が行われてきた4)。魚類相に関する調査については、年に 2-3 回の手網と投網による採集15)、2007 年 5 月から 11 月 まで行われた人工海浜における投網調査 16)、2012 年から *1 東京湾内湾に造成された干潟と海浜の魚類相Fish assemblage and diversity in the developed tidal flat and sandy beach at the Furuhama Park, Ota City, Tokyo, central Japan
*2 Laboratory of Ichthyology, Tokyo Univercity of Marine Science and Technology, 4-5-7 Konan, Minato-ku, Tokyo 108-8477, Japan(東京海洋大 学魚類学研究室)
*3 Department of Ocean Sciences, Tokyo University of Marine Science and Technology (TUMSAT), 4-5-7 Konan, Minato-ku, Tokyo 108-8477, Japan(東京海洋大学学術研究院海洋環境科学部門)
2013 年にかけて 9 か月間行われた人工干潟のタイドプー ルと浅場、またそれをつなぐ澪筋における手網と定置網調 査17)、2016 年 4 月から 2017 年 3 月に人工干潟のタイドプ ールにおける手網調査18)、2016 年 4 月から 2019 年 3 月に 行われた籠網による調査 19)が行われている。それに加え、 定量的な魚類群集調査として2014 年から 2015 年にかけて 小型地曳網を用いた調査20)が行われたが、干潟では9 月、 11 月と 1 月には調査が行われていない。 そこで本研究では村井ら 20)のデータも含めて、2014 年 から2019 年までの 5 年間、より長期にわたる定量的な調 査を実施した。そして、採集された魚類の生活史型や利用 様式を明らかにし、またそれらの結果を東京湾内湾の他の 人工海浜や自然干潟と比較することにより、大森ふるさと の浜辺公園が魚類にどのような場を提供しているのかを 検討・評価した。
調査地点の概要
調査地点は、東京湾内湾の西岸に位置する東京都大田区 の「大森ふるさとの浜辺公園」である(Fig. 1)。同地点で はかつて、土砂や周辺のガス工場より排出されたコークス 殻が明治時代に設置された東京ガスの波除堤防周囲に堆 積し、生物の生息場や鳥の餌場となる干潟や浅場を作り出 していた21)。しかし、産業構造や環境の変化によって遊休 化した結果として、2000 年に緑地、砂浜、浅場、磯場を有 する公園にするため、埋め立てが開始された。 その後、2007 年に同地区は 1.2 ha の砂浜海岸(千葉県君 津産のd50 = 0.2 mm の山砂で養浜:以下、海浜 Sandy beach とする)と1.0 ha の泥質干潟(在来の干潟を 200 m ほど沖 合に移設し、多孔質の礫や岩を配置:以下、干潟 Tidal flat とする)、さらにそれらの間には水深1.5 m、面積 4.6 ha の 浅場を有する公園として開園した4)。また、周辺運河との 境界部となる浅場の南北の端部にはそれぞれ、海底から高 さ約 4 m の砂留潜堤 Submerged dike が配置されている (Fig. 1)。同潜堤は、浅場からの土砂の流出や、周辺運河 の底層に広がる貧酸素水の流入を抑制している14)。同公園 は施工前より様々な目的で環境調査が行われている。今も なお、底質、水質およびベントスなどの調査が継続されて いる4)。調査方法
本研究の調査地点は、大森ふるさとの浜辺公園の海浜と 干潟の2 地点とした。採集期間は 2014 年 5 月から 2019 年 の4 月、月に 1 回、大潮前後の日中の干潮時に行った。た だし、干潟において2014 年 9 月と 11 月、2015 年 1 月と 6 月、2019 年の 4 月は未調査である。また、2016 年と 2017 には8 月に調査を行えなかったため、それぞれ 9 月に 2 回 行った。曳網回数は2 回(2014 年 5 月と 6 月のみ 3 回)と した。2 地点で調査を行った回数や曳網数が異なるため、 採集された個体数は総個体数(Table 1)を除いて、各月に 採集された個体数を1 曳網当たりの個体数に直してから算 出した値を用いた。 採集にはKANOU et al.22)を参考に作製された小型地曳網 (袖網部:長さ4.5 m、高さ 1.0 m、網目 2.0 mm;胴網部 から袋網部:網口の幅2.0 m、高さ 1.0 m、長さ 5.5 m、網 目0.8 mm)を用い、水深 1 m 以浅を汀線に沿って 25 m 曳 網した。また網口の幅を4.0 m になるように曳網し、1 回 の曳網で100 m2の魚類を採集した。採集物は現地で10% 海水ホルマリンを用い固定し、研究室に持ち帰った後に魚 類だけを抜き出し、種の同定、個体数の計数、体長の測定 および発育段階の決定を行った。 発育段階は加納ほか6)に従い4 つ(仔魚:鰭条が定数に 達していない時期のもの、稚魚:鰭条が定数に達し鱗も出 始めているが、体形や模様が成魚と異なる時期のもの、若 魚:鱗の分布や模様はほぼ完成し体形も成魚に近いが未成 熟のもの、成魚:性的に成熟しているもの)に区分した。 種の同定は中坊(編)23)と沖山(編)24)に従った。科の配 列と学名、和名は主に中坊(編)23)に従った。しかし、マ ル タ Tribolodon brandtii maruta の 学 名 は SAKAI andAMANO25)に、イシガレイ Platichthys bicoloratus とマコガレ イ Pseudopleuronectes yokohamae の学名は尼岡26)に、クサ
フグ Takifugu alboplumbeus の学名は MATSUURA27)に従った。 また、これら採集された魚類は主に加納ほか6)に従い、 生活史型を6 つ(河口魚:河口域もしくは河口域と海域の 境界で産卵を行い河口域で全生活史のほぼ全てを完結す る種(仔魚期に海域へ分散することもある)、海水魚:海 域で産卵を行う種で、基本的には河川淡水域では成長しな い、遡河回遊魚:産卵のために河川を遡り、淡水域で産卵 を行う種、降河回遊魚:産卵のために河川を降り、海域で 産卵を行う種、両側回遊魚:産卵を河川淡水域で行い、仔 魚は流下して海域で少し成長した後に河川へ戻り、河川で 成長・成熟する種、淡水魚:河川淡水域で産卵を行い、生 活史を淡水域で完結させる種)、利用様式を3 つ(滞在型: 仔魚もしくは稚魚から成魚まで出現し、本調査地点で生活 史をほぼ完結させる種、一時滞在型:仔魚から稚魚、稚魚 から若魚、仔魚から若魚と複数の発育段階が出現するが、 成魚までは滞在しない種、通過・遇来型:1 つの発育段階 のみ、もしくは不連続の発育段階で出現する種)に区分し た。 採集と同時に水温と塩分を YSI/Nanotech 社の EC-300 で、溶存酸素量(以下、DO)を HORIBA 社の OM-1 で 測定した。なお、2016 年からはこれら 3 つの環境項目は、 YSI/Nanotech 社の YSI Pro2030 で測定した[干潟では、 2014 年 5 月、9 月、11 月、2015 年 2 月(DO のみ)、2017 年9 月の 1 回目は未測定]。また、2 地点間で底質の比較を 行うため、2018 年の 3 月、6 月、9 月と 12 月に内径 4.5 cm、 高さ6.5 cm のコアを用いて底土を採集した。採取した底土
は河野ほか28)に従い処理を行い、各ふるい(6 段階:2000、 1000、500、250、125、63μm)に残留した試料の割合と中 央粒径値、泥分(粒径63 μm 以下)を求めた。 ま た 、 群 集 組 成 の 比 較 を 行 う た め に 地 点 毎 の Shannon-Wiener の多様度 H’と Pielou の均衡度 J’を求め た(ともに、木元29)より引用)。さらに、横軸に出現個体 数の第1 位種から順に第 n 位種をとり、縦軸に総個体数に 占める各種の割合を優占順にプロットした「簡易多様度判 定グラフ」を作成することで多様度を比較した。比較対象 は、本調査地点における1 年間の調査(村井ほか,20): 以下、 前調査)と、河野ほか28)と村瀬ほか30)の東京都大田区に位 置する羽田(本調査地点より東に約4 km の場所に位置し、 東京湾内湾に対して開放的; 羽田空港の北東の小規模な人 工前浜干潟: Fig. 1 の Haneda)、京浜島(本調査地点より東 に約2.5 km で、本調査地点と羽田との中間的な立地; 人工 島である京浜島の南東部に位置するつばさ公園に自然に できた干潟: Fig. 1 の Keihin-jima)、多摩川(本調査地点よ り南に約3.5 km で、海老取川と多摩川が合流する地点にで きた自然干潟: Fig. 1 の Tama-gawa R.)における約 6 年間の 調査とした。
結果
1.環境項目
水温、塩分およびDO は両地点で同じような値を記録し た(Fig. 2)。水温の最高は、海浜で 2018 年 7 月の 33.8℃、 干潟で2014 年 7 月の 33.6℃であった。最低は、海浜と干 潟ともに2018 年 1 月の 9.5℃であった。塩分はおおむね夏 季に低く冬季に高く、平均±SD は海浜で 17.4±4.38、干潟 で18.0±4.94 であった。最高は海浜で 2017 年 4 月の 25.7、 干潟で2017 年 1 月の 27.6 であった。最低は、海浜と干潟 ともに2016 年 9 月でそれぞれ 8.5 と 7.8 であった。DO の 平均±SD は海浜で 7.6±3.26 mg/L、干潟で 7.2±2.99 mg/L であった。最高は海浜と干潟ともに2016 年 7 月で、それ ぞれ18.2 mg/L と 16.9 mg/L、最低は海浜と干潟ともに 2018 年6 月で、それぞれ 2.9 mg/L と 3.1 mg/L であった。 底質は海浜と干潟で違いがみられ、海浜の方が粗かった (Fig. 3)。海浜(図中の S)における粒径別の割合は、3 月と 6 月は 250-500μm、9 月は 500-1000μm、12 月は 1000-2000μm の粒径が高かった。対して干潟(T)におけ る粒径別の割合は、全ての月で125-250μm の粒径が高か った。中央粒径値は、海浜(S)で 402.5-635.1μm で平均 は526.2μm、干潟(T)で 198.2-246.3μm で平均は 217.9 μm であった。泥分は、海浜で 0.2-0.6%、平均は 0.4%、 干潟で0.4-2.1%、平均は 1.0%であった。2.出現した魚類
本研究で採集された魚類は海浜で19 科 40 種 60,050 個体 (122 曳網、1 曳網あたり 492.2 個体)、干潟で 19 科 38 種 44,807 個体(109 曳網、411.1 個体)であった(Table. 1: た だし、ウグイ属不明複数種 Tribolodon spp. 、チチブ属不明 複数種 Tridentiger spp. とハゼ科不明複数種 Gibiidae spp. は種数に含めない)。科別の割合はハゼ科魚類が優占し、 種数では海浜で16 種(海浜の種数の 40%)、干潟で 15 種 (干潟の種数の39%)、個体数でも海浜で 52,122 個体(海 浜の総個体数の87%)、干潟で 41,269 個体(干潟の総個体 数の92%)であった。優占した上位 5 種は海浜でビリンゴ Gymnobobius breunigii (海浜の総個体数の 45.8%)、マハゼ Acanthogobius flavimanus (35.9%)、ボラ Mugil cephalus cephalus (9.0%)、スミウキゴリ Gymnogobius petschiliensis (3.7%)、アユ Plecoglossus altivelis altivelis (1.9%)であ った。干潟の優占種は、マハゼ(干潟の総個体数の42.3%)、 ビリンゴ(41.4%)、ボラ(4.2%)、スミウキゴリ(3.2%)、 エドハゼ Gymnogobius macrognathos (3.1%)であった。 これら5 種で総個体数のうち、海浜では 96.4%、干潟では 94.4%を占めた。それぞれの地点でのみ出現した種は、海 浜でウグイ Triborodon hakonensis(7 個体)、ウグイ属不明 複 数 種 Triborodon spp. ( 106 )、 ガ ン テ ン イ シ ヨ ウ ジ Hippichthys penicillus(1)、イケカツオ Scomberoides lysan (1)、シマイサキ Rhyncopelates oxyrhynchus(1)、マサゴ ハ ゼ Pseudogobius masago ( 1 )、 マ コ ガ レ イ Pseudopleuronectes yokohamae ( 2 )、 ク サ フ グ Takifugu alboplumbeus(2)の 8 種以上 121 個体で海浜の総個体数の 0.2 % で あ っ た 。 ま た 、 干 潟 で は マ イ ワ シ Sardinops melanostictus(2)、メジナ Girella punctata(2)、トサカギ ンポ Omobranchus fasciolatoceps(4)、イダテンギンポ Omobranchus punctatus(3)、チチブ属不明複数種 Tridentiger spp.(11)、ヒガンフグ Takifugu pardalis(3)の 6 種以上 25 個体で干潟の総個体数の 0.1%以下であった。3.種数と個体数の経月変化
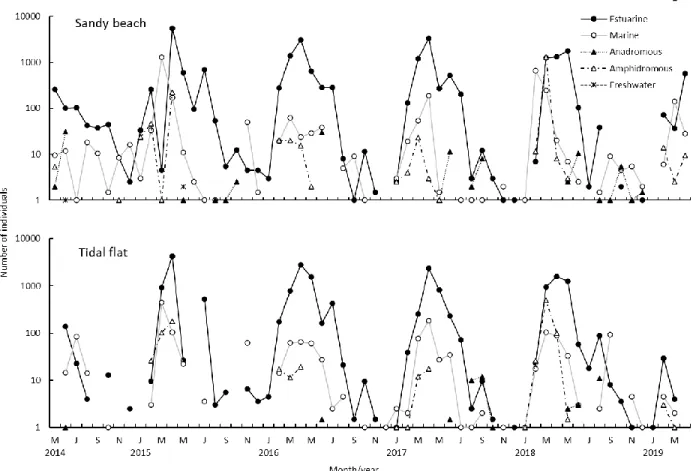
種数はおおむね、春季に多くなり、夏季から冬季にかけ て少なくなった(Fig. 4)。海浜における種数の最多は、2014 年8 月と 2015 年 4 月の 13 種であり、最少は 2016 年 12 月 と2019 年 1 月の 0 種であった。干潟における最多は、2015 年4 月の 13 種で、最少は 2015 年 10 月の 0 種であった。 個体数は両地点ともに3 月や 4 月に最多を記録し、春季 に多かった(Fig. 4)。春季以降は個体数が減少し、秋季と 冬季は少なかった。海浜における個体数の最多は、2015 年4 月で一曳網当たり 5947.5 個体であり、最少は 2016 年 12 月と 2019 年 1 月の 0 個体であった。干潟における個体 数の最多も2015 年 4 月で、一曳網当たり 4541 個体が採集され、最少は2015 年 10 月の 0 個体であった。
4.生活史型別の割合と出現の経月変化
ウグイ属不明複数種、チチブ属不明複数種およびハゼ科 不明複数種は生活史型の決定ができないため、ここではそ れらを除外した海浜40 種 29752 個体、干潟 38 種 22308.2 個体を対象とした。 海浜と干潟における生活史型別の割合は、次のように種 数と個体数ともにほぼ同じであった(Fig. 5)。海浜におけ る種数は、海水魚が最も多く22 種(海浜の総種数の 55.0%) であった。次いで、河口魚13 種(32.5%)、両側回遊魚 3 種(7.5%)、遡河回遊魚と淡水魚がそれぞれ 1 種(それそ れ 2.5%)であった。対して、干潟における種数も海水魚 が最も多く21 種(干潟の総種数の 55.3%)であった。次 いで、河口魚13 種(34.2%)、両側回遊魚 3 種(7.9%)、 遡河回遊魚1(2.6%)となり、淡水魚は採集されなかった。 海浜における個体数は、河口魚が最も多く24678.4 個体(海 浜の総個体数の 82.9%)であった。次いで海水魚 3240.8 個体(10.9%)、両側回遊魚 1718.3 個体(5.8%)、遡河回遊 魚111.5 個体(0.4%)、淡水魚 3 個体(0.1%以下)であっ た。干潟における個体数も河口魚が最も多く19588.9 個体 (干潟の総個体数の 87.8%)であった。次いで、海水魚 1673.1 個体(7.5%)、両側回遊魚 1003 個体(4.5%)、遡河 回遊魚43.2 個体(0.2%)であった。 月別の種数では、河口魚は両地点ともに年間を通して採 集され、海浜では2014 年 8-10 月、2016 年 4 月と 9 月に最 多の6 種、干潟では 2015 年 4 月と 2016 年 3 月に最多の 7 種が出現した(Fig. 6)。また、毎年 2-5 月に増加する傾向 が認められた。海水魚は、海浜では2014 年 8 月に最多の 7 種、干潟では2018 年 2 月に最多の 6 種が出現した。両地 点ともに、2-5 月に増加する傾向が認められた。遡河回遊 魚は両地点ともに5-12 月(ただし7 月を除く)に出現した。 両側回遊魚は両地点ともに毎年1-5 月に出現したが、海浜 においては2014 年 11 月に1 種のみ出現したことがあった。 淡水魚は、海浜において2014 年 6 月と 2015 年 5 月に 1 種 のみ確認された。 月別の個体数では、河口魚は3-5 月といった春に多く採 集され、2015 年 4 月に海浜で 5553 個体、干潟で 4258 個体 と最多を記録した(Fig. 7)。海水魚は 2-4 月に多く採集さ れ、2015 年 3 月に海浜で 1300 個体、干潟で 447 個体と最 多を記録した。遡河回遊魚は海浜で2014 年6 月に31 個体、 干潟で2017 年 9 月の 2 回目で 12 個体と最多を記録したが、 数個体の出現にとどまる月も多かった。両側回遊魚は3 月 か4 月に多く採集され、2018 年 3 月に海浜で 1279 個体、 干潟で488.5 個体と最多を記録した。淡水魚は海浜におい て2014 年 6 月に 1 個体と 2015 年 5 月に 2 個体採集された だけであった。5.利用様式別の割合と出現の経月変化
利用様式においても、ウグイ属不明複数種、チチブ属不 明複数種とハゼ科不明複数種を除外した海浜 40 種 29752 個体、干潟38 種 22308.2 個体を対象とした。 海浜における種数では、一時滞在型が最も多く19 種(海 浜の総種数の47.5%)であった(Fig. 8)。次いで、通過・ 偶来型17 種(42.5%)、滞在型 4 種(10.0%)であった。 しかし個体数では滞在型が最も多く24468.9 個体(海浜の 総個体数の82.2%)であった。次いで、一時滞在型 5185.9 個体(17.4%)、通過・偶来型 97.2 個体(0.3%)であった。 干潟における種数は一時滞在型と通過・偶来型が多く16 種(それぞれ干潟の総種数の42.1%)となった(Fig. 8)。 滞在型は6 種(15.8%)であった。個体数も海浜同様、最 も多く出現したのは滞在型で19572.6 個体(87.7%)であ った。次いで一時滞在型2687 個体(12.0%)、通過・偶来 型48.6 個体(0.2%)という結果になった。 月別の種数は、滞在型は年間を通して出現が確認され、 特に干潟においては3-5 月といった春季に多く記録された (Fig. 9)。一時滞在型は 2-5 月に多く出現し、海浜では 2015 年4 月の 10 種、干潟では 2018 年 2 月の 10 種が最多であ った。通過・偶来型は10-1 月といった秋から冬季にかけて は確認されないことが多かった。 月別の個体数は、滞在型は 3-5 月に多く出現した(Fig. 10)。最多は 2015 年 4 月に記録され、海浜で 5539.5 個体、 干潟で4254.5 個体であった。また、10-1 月の出現は少なか った。一時滞在型も3 月や 4 月に多く出現し、秋から冬は 少なかった。最多は2018 年 3 月に記録され、海浜で 1496.5 個体、干潟で593.5 個体であった。通過・偶来型に季節性 は見られず、海浜では2018 年 3 月の 31.5 個体、干潟では 2015 年 5 月の 15 個体が最多であった。6.生活史型別の利用様式
河口魚の種数では、海浜と干潟で違いがみられた(Fig. 11)。海浜においては、一時滞在型が 7 種(海浜に出現し た河口魚の総種数の53.8%)で多く、滞在型は 4 種(30.8%)、 通過・偶来型は2 種(15.4%)であった。対して干潟では 滞在型と通過・偶来型がそれぞれ6 種(それぞれ、干潟に 出現した河口魚の総種数の46.2%)で、一時滞在型は 1 種 (7.7%)であった。しかし、個体数は 2 地点で似た傾向が 見られ、滞在型が圧倒的に多く出現し、海浜で24468.9 個 体(海浜に出現した河口魚の個体数の 99.2%)、干潟で 19572.6 個体(干潟に出現した河口魚の個体数の 99.9%) であった。一時滞在型は海浜で208 個体(0.8%)と干潟で 10 個体(0.1%以下)、通過・偶来型は海浜で 1.5 個体と干 潟で6.3 個体(それぞれ、0.1%以下)と少なかった。 海水魚は両地点において似た傾向が見られた。両地点に おいて滞在型は確認されず、種数は海浜では通過・偶来型が多く14 種(海浜に出現した海水魚の種数の 63.6%)、次 いで一時滞在型が8 種(36.4%)であった。干潟では一時 滞在型が11 種(干潟に出現した海水魚の種数の 52.4%)で、 通過・偶来型は10 種(47.6%)であった。対して個体数で は一時滞在型が多く、海浜で3048.1 個体(海浜に出現した 海水魚の個体数の97.1%)、干潟で 1630.8 個体(干潟に出 現した海水魚の個体数の97.5%)であった。通過・偶来型 は海浜で92.7 個体(2.9%)、干潟で 42.3 個体(2.5%)と なった。 遡河回遊魚と両側回遊魚は両地点ですべて一時滞在型 であった。また、淡水魚は海浜のみで出現し、すべて通過・ 偶来型であった。
7.多様性
本調査地点の出現した種数は前調査の海浜は 33 種、干 潟は29 種であったことに対して本調査は海浜で 40 種、干 潟で38 種であった(Table 2)。簡易多様度判定グラフ(Fig. 12)に基づく第 1 位種の割合は前調査の海浜は 53.7%、干 潟は45.0%、本調査の海浜は 45.7%、干潟は 42.4%、全体 の80%に達する種順位は 3 種、2 種、2 種、2 種、全体の 95%に達する種順位は 4 種、6 種、5 種、6 種、多様度 H’ は1.30、1.37、1.34、1.38、均衡度 J’は 0.37、0.41、0.36、 0.38 であった(Table 2)。 比較地点に出現した種数は多摩川で50 種、京浜島で 52 種、羽田で45 種であった(Table 2)。簡易多様度判定グラ フ(Fig. 12)に基づく第 1 位種の割合は多摩川で 62.2%、 京浜島で43.9%、羽田で 34.1%、全体の 80%に達する種順 位は2 種、5 種、8 種、全体の 95%に達する種順位は 8 種、 13 種、15 種、多様度 H’は 1.34、1.96、2.40、均衡度 J’ は0.34、0.49、0.63 であった(Table 2)。考察
1.大森ふるさとの浜辺公園の海浜と干潟
大森ふるさとの浜辺公園の海浜と干潟は、水温や塩分、 DO などの環境項目には差がみられなかった。しかし底質 の粒径組成には違いがみられ、海浜の方が干潟よりも粗く、 2 地点で異なることが明らかとなった。 一方、海浜と干潟の2 地点における魚類相調査の結果、 多くの類似性が明らかとなった。出現した種数(海浜 40 種、干潟38 種)や個体数(海浜 492.2 個体/曳、干潟 411.1 個体/曳)、あるいは優占種(上位 4 種は一致)は同様の傾 向を示した。また、各地点のみで出現する種も5 年間の調 査の中で5 個体以下のものがほとんどで、これらは偶然出 現した可能性が高い。さらに、種数や個体数の経月変化(種 数・個体数ともに春をピークに秋から冬は少なくなる)、 生活史型別の出現(種数では海水魚、個体数では河口魚が 多く、各生活史型が出現する月の傾向もほぼ同じ)、利用 様式別の出現(個体数で滞在型が多く、出現する月の傾向 もほぼ同じ)、生活史型別の利用様式の割合のどれをとっ ても同様の傾向が確認された。これは、すでに村井ほか20) でも報告されており、水深4-5 m の運河域と砂留潜堤によ って隔てられた2 地点が水深 1.5 m より浅い浅場内に造成 されたこと21)、および6.8 ha という狭い海域にコンパクト に配置されたこと31)の結果であると考えることができる。2.大森ふるさとの浜辺公園の特徴
東京湾の内湾では、これまでに多くの地点において本調 査と同様の小型地曳網を用いた調査が行われ、各地点にお ける魚類群集の特徴が明らかになってきた。ここでは、桑 原ほか9)の東京都江戸川区に位置する葛西人工渚の西浜 (東京湾内湾で代表的な人工前浜干潟: fig. 1 の Kasai)に おける調査、および河野ほか28)と村瀬ほか30)の羽田、京浜 島、多摩川における調査と比較し、大森ふるさとの浜辺公 園の特徴を明らかにした。 1) 優占するハゼ科魚類 本研究の結果、大森ふるさとの浜辺公園の海浜と干潟で は種数では海水魚が多かったものの、個体数では河口魚が 優占した。河口魚は全てがハゼ科であり(Table 1)、それ らハゼ科河口魚は春に出現のピークを持つことが明らか となった。これは東京湾内湾の干潟のみならず、世界の多 くの内湾や河口汽水域における魚類群集の特徴として知 られ6,7,32)、人工環境であるふるはまも例外ではなかった。 これら河口魚のうち、海浜では4 種(マハゼ、アシシロハ ゼ Acanthogobius lactipes 、ビリンゴとチチブ Tridentiger obscurus )、干潟では 6 種(マハゼ、アシシロハゼ、ビリ ンゴ、エドハゼ、ニクハゼ Gymnogobius heptacanthus とチ チブ)が滞在型として認められた。羽田ではマハゼ、ビリ ンゴおよびヒメハゼ Favonigobius gymnauchen の 3 種、京 浜島ではマハゼ、アシシロハゼ、ビリンゴ、エドハゼ、ニ クハゼおよびヒメハゼの6 種、多摩川ではマハゼ、アシシ ロハゼ、ビリンゴ、エドハゼ、ヒメハゼおよびマサゴハゼ Pseudogobius masago が滞在型として認められている。本 研究の小型地曳網ではマサゴハゼは海浜で1 個体の採集の みであったが、本調査地点の干潟のタイドプールでは、季 節に合わせて異なる発育段階の個体が周年出現すること が知られている18)。また、葛西では滞在型の種が出現しな い。 これらの結果から、大森ふるさとの浜辺公園は、人工海 浜である羽田や葛西とは異なった生息場をハゼ科河口魚に提供しており、むしろ京浜島や多摩川に近い生息場を提 供していたと判断できる。村瀬ほか30)では、京浜島や多摩 川はハゼ科河口魚に定住・長期滞在の場を提供していると 結論付けている。大森ふるさとの浜辺公園も、自然干潟と 同様に、一部のハゼ科魚類には仔魚から成魚までの長期的 な成育場を提供できていることが明らかとなった。 2) 少ない海水魚 海水魚の種数は海浜で22 種、干潟で 21 種と生活史型別 の種数では優占したが、個体数はそれぞれ採集された総個 体数に対して10.9%と 7.5%で少なかった。比較地点の種数 は羽田で28 種、京浜島で 29 種、多摩川で 26 種と本調査 地点より多かったが、葛西では18 種であった(Table 3)。 しかし、どの地点も採集された総種数に対する割合は 55-64%で大きな違いはなかった。総個体数に対する海水魚 の割合は羽田で40.7%と高く、京浜島は 17.8%、多摩川で は4.2%、葛西では 18.3%である。また、一曳網あたりの海 水魚の採集個体数は本調査地点では少ない。これらのこと から、本調査地点は複雑な運河域の奥に立地することから、 海水魚の来遊は少ないことが明らかとなった。 しかし、東京湾内湾の干潟域あるいは砂浜海岸の特徴と して、海水魚にとっては一時的な滞在の場であることが示 唆されている12)。本調査地点でも、海水魚の個体数は海浜 で97.1%、干潟で 97.5%が一時滞在型であった。よって本 調査地点は、個体数は少ないが、来遊した海水魚にとって 成育場として機能していると判断できる。 3) アユの出現 東京湾内湾におけるアユの出現は冬季において顕著で あり、しばしば冬季の優占種になることが知られている30)。 しかし、大森ふるさとの浜辺公園は、アユが遡上する河川 が近くにないことや、入り組んだ運河域の奥に立地するこ とからアユの来遊は少ないとされてきた20)。実際にアユの 出現量の相違には、近隣に大きな河川の有無が関連してい る可能性が示唆されている33)。アユの採集個体数を見ると、 羽田では53.9 個体/曳網、京浜島で 4.0 個体/曳網、多摩川 で5.6 個体/曳網、葛西で 202.4 個体/曳網である。対して本 研究の海浜は9.5 個体/曳網、干潟は 4.7 個体/曳網であり開 放的な羽田や葛西に比べると個体数が少ないことが明ら かであった。よって大森ふるさとの浜辺公園はアユに成育 場を提供できていない可能性が考えられた。しかし、アユ は潮汐により移動をすることが知られる34,35,36)。そのこと を踏まえ丸山ほか37)は本調査地点で2018 年 3 月から 2019 年2 月に潮汐ごとの調査を行い、満潮時にはアユの個体数 が増加することを明らかにした。 以上のことから、本調査地点が海域に分散した仔魚が接 岸する機能は持たないが、ある程度成長し能動的な個体が 来遊する一つの場となっていることを明らかにしている。 東京湾内湾の小型地曳網による調査は干潮時のものがほ とんどであり6,10,11,20,28,30)、確かに干潮時の調査が種の多様 性や群集組成を素早く知る方法として有用であるが38)、満 潮時の調査を合わせて行うことで、より正確にその場の持 つ機能を明らかにできると考えられた。
3.過去または他地点との多様度比較
東京湾内湾の干潟域においては、地点間の多様度比較6) や、同一地点における年代別の多様度比較13)が行われてき た。しかし、人工的に造成された干潟や海浜の多様度を自 然干潟と比較した例はない。前調査20)は、本調査地点を多 摩川河口の天然干潟と比較したが、それは生活史や利用様 式の比較であり、多様度の比較は行われていない。 そこでまず、過去の本調査地点における1 年間の前調査 と5 年間の本調査で多様度を比較し、長期サンプリングの 必要性と効果を明らかにした。次に本調査を、河野ほか28) と村瀬ほか30)の東京都大田区に位置する羽田、京浜島およ び多摩川における調査と比較し、魚類にとっての人工環境 の評価をした。 1) 長期モニタリングの効果と必要性 出現した種数を比較すると、前調査の海浜は 33 種、干 潟は29 種であったことに対して本調査は海浜で 40 種、干 潟で38 種であった。一曳網あたりの個体数は前調査の海 浜が622 個体、干潟が 688 個体であったことに対して本調 査では海浜で492.2 個体、干潟で 411.1 個体であった。生 活史型別の割合や経月変化はほぼ同じであった。 しかし利用様式の割合は、種数では同様の傾向が認めら れたが、個体数では前調査では滞在型(総個体数に対して 海浜で54%、干潟で 45%)と一時滞在型(総個体数に対し て海浜で46%、干潟で 54%)が優占していたことに対して 本調査では滞在型(総個体数に対して海浜で82.2%、干潟 で87.7%)が優占した。 これにともなって、生活史型別の利用様式では、前調査 と異なり河口魚はほとんどが滞在型になった。また、前調 査の遡河回遊魚は通過・偶来型であったが、本調査では一 時滞在型に分類された。 以上のようにして、長期間調査を行うことで短期間の調 査とは異なる結果が得られた。 個体数で、河口魚のほぼすべてが滞在型に分類されたの は、本調査では優占種のマハゼが滞在型に分類されたこと に起因する。マハゼは、前調査では仔魚から若魚の出現で あったが、本調査では仔魚から成魚が確認された。大田区 による調査15)では、より大型の体長100 mm 以上の個体が 採集されている。また、遡河回遊魚の個体数が全て一時滞 在型に分類されたが、これは前調査では稚魚しか採集され なかったマルタが、本調査では稚魚と若魚が採集されたこ とに起因する。マルタも大田区の調査15)でより大型の個体 が採集されている。多様度という観点から見ると、1 年間の調査と 5 年間の 調査で大きな差が見られることはなかった。より長期間の 調査では、短期間の調査より多くの種が確認できたが、多 様度決定グラフ(Fig. 12)に基づく第 1 位種の割合、全体 の80%に達する種順位、全体の 95%に達する種順位、さら に多様度H’と均衡度 J’はほぼ同様の値を示した。 このように、長期間の調査は短期間の調査とはいくつか の点で異なる結果を得ることができ、より正確な魚類群集 を明らかにしたと判断できる。茂木ほか13)でも述べられて いるが、長期的な調査は魚類相の特徴を正確に得ることが でき、その結果は東京湾の環境保全や環境修復のために有 用であると結論付けることができる。 2) 人工干潟と自然干潟の多様度比較 出現した種数は、本調査の海浜が40 種、干潟が 38 種に 対して、多摩川で50 種、京浜島で 52 種、羽田で 45 種で あり、本調査地点では少ない傾向が確認できた。多様度判 定グラフ(Fig. 12)に基づく第 1 位種の割合(低い方が多 様度は高い)は海浜で45.7%、干潟で 42.4%に対して、多 摩川は62.2%と高い値を示したが、京浜島は 43.9%、羽田 は 34.1%であった。全体の 80%に達する種順位と全体の 95%に達する種順位(これらは遅い方が多様度は高い)は 海浜で2 種と 5 種、干潟で 2 種と 6 種に対して、多摩川で 2 種と 8 種、京浜島で 5 種と 13 種、羽田で 8 種と 15 種で あり、京浜島と羽田で多様度が高いことを示している。 多様度 H’(数値が大きいほど多様度が高い)と均衡度 J’ (数値が大きいほど多様度が高い)は本調査の海浜で1.34 と0.36、干潟で 1.38 と 0.38 という値に対して、多摩川で 1.34 と 0.34、京浜島で 1.96 と 0.49、羽田で 2.4 と 0.63 であ り、京浜島や羽田では高い値であった。 以上のことから、多様度判定グラフ、多様度H’と均衡度 J’に基づくと、本調査地点の多様度は多摩川とほぼ同等で あるが、京浜島や羽田よりは低い値であることが明らかに なった。ただし、塩分をみると、多摩川は河川水の影響を 直接受けるため平均10.9 と低い。対して京浜島は平均 14.3、 羽田は21.0 であり、本調査地点の海浜と干潟は 17.5 と 18.1 である。よってこの多様度の低さは塩分に起因するもので はなく、運河の奥に造成された立地の影響や、周辺が運河 域となっており他の干潟と連続性が無いこと、もしくは造 成された人工海浜・干潟の限界であるという可能性が考え られた。 今後は、多様度のみならず、各魚種の食性や発育段階ご との出現を天然干潟と比較し、より具体的に魚類が大森ふ るさとの浜辺公園をどのように利用しているのかを明ら かにしていくことが必要である。またその結果を用いるこ とで、今後の東京湾沿岸域のあり方や、人工環境造成事業 への指針を提示することが期待される。
謝辞
本研究を行うにあたり、採集調査の許可や傭船にご協力 をいただいた大田区の都市基盤整備部の方々および東京 湾遊漁船業協同組合の飯島正宏理事長と組合員の方々に お礼申し上げます。また、東京海洋大学魚類学研究室の学 生の皆さんのご協力により、5 年間という長期間の調査を 実行できたことを感謝いたします。本研究の一部は JSPS 科研費基盤(B)24310028 および基盤(C)15K00654、2019 年度笹川科学助成金(研究番号: 2019-4094)の助成を受け て実施しました。引用文献
1) 国土交通省港湾局・環境省自然環境局. 干潟ネットワークの 再生に向けて. 国立印刷局, 2004, 119pp. 2) 清水誠. 東京湾の魚介類(6)昭和 60 年代の生物相. 海洋と 生物, 1990, 68, 183-189. 3) 鎌谷明善. 海岸線の変貌. 東京湾―100 年の環境変遷, 小倉 紀雄(編), 恒星社厚生閣, 東京, 1993, 20-27. 4) 中瀬浩太. 人工干潟の施工およびモニタリング.建設の施工 企画,2008, 8, 42-47. 5) 環境庁自然保護局. 日本の干潟、藻場、サンゴ礁の現状. 海 中公園センター, 東京, 1997, 291pp. 6) 加納光樹・小池哲・河野博. 東京湾内湾の干潟域の魚類相と 多様性. 魚類学雑誌, 2000, 47, 115-129.7) Hermosilla, J.J., Y. Tamura, M. Moteki and H. Kohno. Distribution and community structure of fish in Obitsu-gawa River Estuary of inner Tokyo Bay, central Japan. AACL Bioflux, 2012, 5(4), 197-222. 8) 那須賢二・甲原道子・渋川浩一・河野博. 東京湾湾奥部京浜 島の干潟に出現する魚類. 東京水産大学研究報告, 1996, 82, 125-133. 9) 桑原悠字・土田奈々・元山崇・河野博・加納光樹・島田裕至・ 三森亮介. 葛西人工渚(東京湾湾奥部)の魚類相. La mer, 2003, 41, 28-36. 10) 山根武士・岸田宗範・原口泉・阿部礼・大藤三矢子・河野博・ 加納光樹. 東京湾内湾2地点(葛西臨海公園と八景島海の公 園)の仔稚魚相. La mer, 2004, 42, 35-42. 11) 河野博・横尾俊博・茂木正人・加納光樹. 東京湾岸に位置す る人工潟湖(新浜湖)の魚類相. 日本生物地理学会会報, 2008, 63, 133-142. 12) 河野博. 東京湾の魚類 研究史と自然誌. 江戸前の環境学 海を楽しむ・考える・学びあう12 章, 川辺みどり・河野博(編), 東京大学出版会, 2012, 85-106. 13) 茂木正人・安田健吾・山本佳子・横尾俊博・河野博・諸星一 信・鈴木信昭・松坂省一・有路隆一. 東京湾京浜島の魚類相 の季節変化と長期モニタリングの必要性. La mer, 2009, 46,
121-134. 14) 竹山佳奈・田中克彦・河野博・木村賢史・中瀬浩太・岩上貴 弘. 東京湾奥部の都市運河域に造成した干潟・海浜環境の長 期的な変遷. 土木学会論文集 B3(海洋開発), 2018, 74, I_510-I_515. 15) 大田区. 平成 30 年度 平和島運河環境調査報告書, 大田区都 市基盤整備部. 2019. 16) 小林麻里・佐々木剛. 大森ふるさとの浜辺公園を活用した水 圏環境教育の有効性の考察と魚類を用いた教材開発の基礎 調査. 水圏環境教育誌, 2008, 1(1), 18-52. 17) 竹山佳奈・木村賢史・上村了美・吉田潤・中瀬浩太・古河恵 太・鎌田弘行. 運河域の干潟上に造成したタイドプールの生 物生息効果. 土木学会論文集 B3(海洋開発), 2013, 69, I_1030-I_1035. 18) 酒井卓・竹山佳奈・中瀬浩太・河野博. 東京湾奥部に再生さ れた干潟域(大森ふるさとの浜辺公園)のタイドプールの魚 類相. 日本生物地理学会会報, 2018, 72, 98-112. 19) 小野寺暁・丸山啓太・竹山佳奈・河野博. 東京湾奥部の直立 護岸(大森ふるさとの浜辺公園)に設置した籠網に蝟集する 魚類の出現様式. La mer, 2020, 58, 59-69. 20) 村井俊太・村瀬敦宣・河野博・竹山佳奈・中瀬浩太・岩上貴 弘. 東京湾の湾奥に再生された干潟と人工海浜(大森ふるさ との浜辺公園)の魚類相. La mer, 2016, 54, 11-27. 21) 岡村知忠・中瀬浩太・佐藤正昭・小寺一宗. 人工干潟造成工 事にともなう干潟環境の変遷について. 海洋開発論文集, 2004, 20, 419-424.
22) Kanou, K., H. Kohno, P. Tongnunui and H. Kurorura. Larvae and juveniles of two engraulidid species, Thryssa setirostris and T.
hamiltoni, occurring in the surf zone at Trang, southern Thailand.
Ichthyol. Res., 2002, 49, 401-405.
23) 中坊徹次(編). 日本産魚類検索 全種の同定 第三版. 東 海大学出版会, 2013, 神奈川県秦野市.
24) 沖山宗雄(編). 日本産稚魚図鑑 第二版. 東海大学出版会, 2014, 神奈川県秦野市.
25) Sakai, H. and S. Amano. A new subspecies of Anadoromous Far Eastern Dace, Tribolodon brandtii maruta subsp. Nov. (Teleostei, Cyprinidae) from Japan. Bull. Natl. Mus. Nat. Sci., Ser. A, 2014, 40, 219-229.
26) 尼岡邦夫. 日本産ヒラメ・カレイ類. 東海大学出版部, 秦野,
2016, 229pp.
27) Matsuura, K. Taxonomic and Nomenclatural Comments on Two Puffers of the Genus Takifugu with Description of a New Species,
Takifugu flavipterus, from Japan (Actinopterygii,
Tetraodontiformes, Tetraodontidae). Bull. Natl. Mus. Nat. Sci., Ser. A, 2017, 43, 71-80. 28) 河野博・茂木正人・石丸隆・関沢和彦. 羽田空港滑走路建設 にともなう多摩川河口域の魚類への影響. 羽田空港周辺水域 環境調査研究委員会(編), 羽田周辺水域環境調査最終報告 書~研究の総括と今後の展望~: 152-166. 羽田周辺水域環境 調査研究委員会, 東京, 2014. URL: https://www.tbeic.go.jp/haneda-iinkai/view/iinkai/Download/2014 0317/07_3bu5syou.pdf 29) 木元新作. 集団生物学概説. 共立出版, 東京, 1993, 188pp. 30) 村瀬敦宣・角張ちひろ・加瀬喜弘・齊藤有希・河野博. 羽田 空港滑走路の建設は多摩川河口干潟域を利用する魚類にど のように影響するか? 日本生物地理学会会報, 2014, 69, 57-75. 31) 里見勇・藤沢康文・五十嵐美穂. 大森ふるさとの浜辺整備事 業-事業実施と合意形成のプロセス-. 海洋開発論文集, 2004, 20, 299-304. 32) 加納光樹. 干潟域の魚類. 河野博(監修)東京湾魚の自然誌, 平凡社, 東京, 2006, 73-81. 33) 内田肇・須田有輔・町井紀之. 土井ヶ浜海岸の砕波帯に出現 する魚類. 水産大学校研究報告, 1998, 46, 163-173. 34) 塚本勝己・望月賢二・大竹二雄・山崎幸夫. 川口水域におけ るアユ仔稚魚の分布・回遊・成長. 水産土木,1989, 25, 45-57. 35) 東健作. アユの海洋生活期における分布生態. 高知大学海洋 生物教育研究センター研究報告, 2005, 23, 59-112. 36) Kanou, K., M. Sano and H. Kohno. Larval and juvenile fishes
occurring with flood tides on an intertidal mudflat in the Tama River estuary, central Japan. Ichthyol. Res., 2005, 52, 158-164. 37) 丸山啓太・河野博. 東京湾奥部の運河域に造成された人工海
浜(大森ふるさとの浜辺公園)におけるアユ仔稚魚の出現. La mer, 2020, 58, 51-57.
38) Morrison, M. A., M. P. Francis, B. W. Hartill, and D. M. Parkinson. Diurnal and tidal variation on the abundance of the fish fauna of a temperate tidal mudflat. Estuarine, Coastal and Shelf Science, 2002, 54, 793-807.
東京湾内湾に造成された海浜と干潟の魚類相と多様性
丸山啓太*2・河野 博*3・竹山佳奈*4・中瀬浩太*4
*2東京海洋大学魚類学研究室 *3東京海洋大学学術研究院海洋環境科学部門 *4五洋建設株式会社環境事業部 東京湾内湾に造成された海浜と干潟が魚類にどのような場を提供しているのか明らかにすることを目的 として、月に一度の小型地曳網による調査を5 年間行った。調査地点は東京都大田区の「大森ふるさとの 浜辺公園」で、同公園は海浜(1.2 ha)と干潟(1.0 ha)、それらの間に浅場(4.6 ha)の海域を有する公園 である。周辺は水深約5 m の運河域であるが、運河域との境界に海底から高さ約 4 m の砂留潜堤が造成さ れたため、浅場の水深は最も深い部分で1.5 m 程度となっている。採集された魚類は海浜で 19 科 40 種 60,050 個体、干潟で19 科 38 種 44,807 個体であった。2 地点の底質には違いがみられたが、出現する種数や個体 数、または生活史型や利用様式から魚類相はほぼ同様であった。本調査地点はハゼ科河口魚が優占し、ま たそれらのハゼ科河口魚には仔魚から成魚までの長期的な成育場を提供していることが明らかとなった。 しかし、運河域の最奥に立地していることから、海水魚の出現は少なかった。また、多摩川河口域の3 つ の干潟と多様度の比較を行うと、本調査地点は劣ることが明らかとなった。 キーワード: 仔稚魚,魚類相,干潟,東京湾Family and Species No. of
individuals Size range(SL,mm)
Developmental Stage Life-style category Life-cycle category Clupeidae Konosirus punctatus 10 / 24 5.6-72.0 / 6.8-36.9 L, Y / L, Y P&S / P&S M Sardinops melanostictus 0 / 2 7.8-8.4 L P&S M Sardinella zunasi 8 / 55 7.0-9.5 / 7.7-17.6 L / L P&S / P&S M Cyprinidae
Tribolodon brandtii maruta 256 / 87 16.8-99.9 / 18.9-94.8 J-Y / J-Y T / T An Tribolodon hakonensis 7 / 0 38.6-54.8 J P&S F Tribolodon spp. 106 / 0 21.7-34.9 J P&S Plecoglossidae
Plecoglossus altivelis altivelis 1,160 / 507 11.5-40.5 / 8.8-50.6 L-J / L-J T / T Am Syngnathidae
Hippichthys penicillus 1 / 0 81.2 Y P&S M Mugilidae
Chelon haematocheilus 4 / 3 16.1-26.9 / 23.8-45.9 J / J P&S / P&S M Mugil cephalus cephalus 5,414 / 1,903 19.4-107.0 / 20.8-63.9 J-Y / J-Y T / T M Atherinidae
Hypoatherina valenciennei 11 / 187 8.4-44.7 / 6.6-34.5 L-Y / L-J T / T M Platycephalidae
Platycephalus sp. 2 15 / 1 8.0-76.8 / 37.3 J-Y / Y T / P&S M Lateolabracidae
Lateolabrax japonicus 555 / 186 11.3-104.2 / 10.6-44.1 L-Y / L-J T / T M Carangidae Scomberoides lysan 1 / 0 31.2 J P&S M Leiognathidae Nuchequula nuchalis 1 / 186 6.3 / 6.3-18.7 L / L-J P&S / T M Sparidae Acanthopagrus latus 162 / 176 10.6-26.6 / 9.7-33.2 L-J / L-J T / T M Acanthopagrus schlegelii 9 / 76 9.2-26.9 / 9.5-30.4 L-J / L-J T / T M Sillaginidae Sillago japonica 35 / 13 7.7-41.3 / 8.4-33.5 L-J / L-J T / T M Teraponidae Rhynchopelates oxyrhynchus 1 / 0 19.8 J P&S M Terapon jarbua 63 / 1 14.8-33.1 / 16.2 J / J P&S / P&S M Girellidae
Girella punctata 0 / 2 16.3, 16.4 J P&S M Stichaeidae
Dictyosoma burgeri 3 / 9 7.1-9.4 / 4.7-12.8 L / L-J P&S / T M Pholidae
Pholis crassispina 2 / 5 56.2-76.4 / 8.4-81.2 Y / L-Y P&S / T M Blenniidae
Omobranchus fasciolatoceps 0 / 4 3.2-4.4 L P&S M Omobranchus punctatus 0 / 3 3.7-5.1 L P&S M Gobiidae
Acanthogobius flavimanus 21,576 / 18,972 7.2-88.6 / 6.9-94.7 L-A / L-A R / R E Acanthogobius lactipes 94 / 195 4.8-51.9 / 4.1-49.0 L-A / L-A R / R E Chaenogobius gulosus 114 / 456 3.3-32.4 / 3.1- 61.77 L-Y / L-Y T / T M Eutaeniichthys gilli 2 / 2 3.9-8.4 / 6.8-9.8 L / L P&S / P&S E Favonigobius gymnauchen 83 / 20 8.6-51.1 / 10.1-45.2 J-Y / J-Y T / T E Glossogobius olivaceus 18 / 1 5.0-16.2 / 11.0 L-J / L T / P&S E Gymnogobius breunigii 27,505 / 18,558 4.1-53.8 / 4.3-54.4 L-A / L-A R / R E Gymnogobius heptacanthus 8 / 67 13.0-38.0 / 4.9-60.7 L-J / L-A T / R E Gymnogobius macrognathos 273 / 1,403 7.4-30.8 / 3.9-35.0 L-Y / L-A T / R E Gymnogobius petschiliensis 2,219 / 1,452 5.4-29.5 / 6.6-30.2 L-J / L-J T / T Am Gymnogobius urotaenia 76 / 48 6.6-27.5 / 6.6-27.3 L-J / L-J T / T Am Luciogobius guttatus 11 / 1 7.2-11.8 / 14.1 L-J / J T / P&S E Mugilogobius abei 8 / 1 3.9-12.5 / 11.5 L-J / L T / P&S E
Pseudogobius masago 1 / 0 11.9 J P&S E
Redigobius bikolanus 15 / 4 4.9-13.6 / 7.1-13.8 L-J / J T / P&S Am Tridentiger obscurus 119 / 89 6.3-46.6 / 6.1-42.6 L-A / L-A R / R E Tridentiger spp. 0 / 11 4.3-8.4 L
GOBIIDAE spp. 10 / 24 5.8-8.0 / 3.3-8.1 L -Pleuronectidae
Platichthys bicoloratus 84 / 67 9.0-30.0 / 9.2-48.9 J / J-Y P&S / T M Pseudopleuronectes yokohamae 2 / 0 10.1-11.2 J P&S M Triacanthidae
Triacanthus biaculeatus 6 / 3 6.2-40.1 / 5.3-6.9 J / J P&S / P&S M Tetraodontidae Takifugu alboplumbeus 2 / 0 15.2-40.7 J P&S M Takifugu pardalis 0 / 3 6.5-9.9 J P&S M No. of species 40 / 38 No. of individuals 60,050 / 44,807 No. of towings 122 / 109 No. of individuals/towing 492.2 / 411.1
Sandy beach / Tidal flat
Developmental stage (A, adult; J, juvenile; L, larva; Y, young)
Life-cycle category (Am, amphidromous fishes; An, anadromous fishes; E, estuarine fishes; F, freshwater fishes; M, marine fishes) Life-style category (P&S, passersby and stays; R, resident; T, transient)
Place Year No. of sp. Percentage of 1st ranked sp. Rank of sp. over 80% Ranked of sp. over 95% Species diversity (H') Evenness (J') Sandy beach 2014-20191) 40 45.7 2 5 1.34 0.36 Tidal flat 2014-20191) 38 42.4 2 6 1.38 0.38 Sandy beach 2014-20152) 33 53.7 3 4 1.30 0.37 Tidal flat 2014-20152) 29 45.0 2 6 1.37 0.41 Tama-gawa R. 2006-20133) 50 62.2 2 8 1.34 0.34 Keihin-jima 2006-20133) 52 43.9 5 13 1.96 0.49 Haneda 2006-20133) 45 34.1 8 15 2.40 0.63
1) Present Study, 2) Murai et al. (2016), 3) Kohno et al. (2014) and Murase et al. (2014).
Table 2. Comparisons of species diversities represented by number of species, the percentage of first ranked species, species rank over 80% and 95%, species diversity (H’) and Evenness (J’) of fishes collected at the sandy beach and tidal flat of Furuhama Park, Tama-gawa River, Keihin-jima and Haneda
Place No. of sp. Ratio of sp. (%) Number of ind.
per towing Ratio of ind. (%)
Sandy beach1) 22 55.0 54.0 10.9 Tidal flat1) 21 55.3 31.0 7.5 Tama-gawa R.2) 26 55.3 101.4 4.2 Keihin-jima2) 29 63.0 192.1 17.8 Haneda2) 28 63.6 195.9 40.7 Kasai3) 18 58.1 409.6 18.3
1) Present study, 2) Kohno et al. (2014) and Murase et al. (2014), 3) Kuwabara et al. (2003).
Table 3. Comparisons of number of species and individuals of marine fishes collected at the sandy beach and tidal flat of Furuhama Park, Tamagawa River, Keihin-jima, Haneda and Kasai in the inner Tokyo Bay
Fig. 1 Map showing the sampling sites in the present study (solid circles) and reference sites (open circles) in the inner Tokyo Bay.
Fig. 2 Monthly changes of water temperature (upper), salinity (middle) and dissolved oxygen (lower) in the sandy beach (open circles) and tidal flat (solid circles) in the inner Tokyo Bay.
Fig. 3 Particle size ratios (bars) and median particle sizes (solid circles) of bottom sediment in the sandy beach (S) and tidal flat (T) in the inner Tokyo Bay.
Fig. 4 Monthly changes of the numbers of species (upper) and individuals (lower) in the sandy beach (open circles) and tidal flat (solid circles) in the inner Tokyo Bay.
Fig. 5 Percentage of numbers of species and individuals by life-cycle categories in the sandy beach and tidal flat in the inner Tokyo Bay.
Fig. 6 Monthly changes of number of species by life-cycle categories in the sandy beach (upper) and tidal flat (lower) in the inner Tokyo Bay.
Fig. 7 Monthly changes of number of individuals by life-cycle categories in the sandy beach (upper) and tidal flat (lower) in the inner Tokyo Bay.
Fig. 8 Percentage of numbers of species and individuals by life-style categories in the sandy beach and tidal flat in the inner Tokyo Bay.
Fig. 9 Monthly changes of number of species by life-style categories in the sandy beach (upper) and tidal flat (lower) in the inner Tokyo Bay.
Fig. 10 Monthly changes of number of individuals by life-style categories in the sandy beach (upper) and tidal flat (lower) in the inner Tokyo Bay.
Fig. 11 Percentage of numbers of species (upper) and individuals (lower) by life-style categories (P&S, passersby and stays), shown by each life-cycle categories (Am, amphidromous fishes; An, anadromous fishes; E, estuarine fishes; F, freshwater fishes; M, marine fishes) and sampling sites (S, sandy beach; T, tidal flat) in the inner Tokyo Bay.
Fig. 12 Relationships between the accumulated percentages of individual number and the species rank of fishes collected in the sandy beach and tidal flat, Tama-gawa River, Keihin-jima and Haneda in the inner Tokyo Bay. Arrows with letters a-g indicate the number of species in each sampling.