Japanese Psychological Research
1995, Vol.37, No.4, 240-246 Short Report
Temporal
distribution
of drinking
in variable
time
schedule-induced
drinking
in rats1
YOSHIHISA
HAMAMURA2
Department of Liberal Arts, National Defence Academy, 1-10-20 Hashirimizu, Yokosuka,
Kanagawa
239
TAKASHI
OKADA
Department
of Psychology,
Faculry
of Letters,
University
of Tokyo,
Hongo,
Bunkyo-ku,
Tokyo
113
KAZUYUKI
YAMADA
Branch
of Business
Management,
Tokyo
Metropolitan
College
of Commerce,
1-2-1
Harumi,
Chuo-ku, Tokyo
104
The aim of this experiment
is to study the effect
of the length
of the inter-food
interval
(IFI) on
immediately
following
adjunctive
drinking in rats, under variable
time 112.5
s schedule. The
U-shaped relation
was found between
the length
of the IFI and the latency
of immediately
following
licking. The animals
began to drink earlier
and stopped
drinking
earlier following
intermediate
IFI
than following
shorter
or longer IFIS.
No systematic
relation
was observed
between
the length
of
the IFI and the number
of immediately
following
licking responses. It was suggested
that under
FT schedules
in previous
studies
the expected
length of the succeeding
IFI affected
the amount
of
drinking,
whereas
under the VT schedule
of the present
experiment
the length
of the preceding IFI
affected
the latency
of drinking.
Key words: schedule-induced
drinking, wheel-running
activity,
variable-time
schedule,
rats.
Under periodic schedules of food deliv-ery. the food-deprived rats drink excessive amounts of water immediately after each
food delivery. This behavior, usually called 'schedule -induced drinking'
, is viewed as psychogenic drinking, because it typically occurs in the absence of water deprivation
(Falk, 1961, 1966, 1969).
Amount of schedule-induced drinking is affected by inter-food interval (IFI) length;
drinking per food pellet is at a maximum level when IFI of fixed time (FT) schedules is set at 120 or 240 s, and decreases with longer or shorter IFIs (Rosellini & Burdette,
1980; Segal, Oden, & Deadwyler, 1965).
However, there is some discrepancy among the findings about the effect of IFI length on
temporal distributions of drinking. Cohen,
Looney, Campagnoni, and Lawler (1985)
compared findings in the previous studies
and concluded that schedule-induced
drink-ing generally reached at a maximum level within 15 or 20 s after each food delivery, regardless of the IFI length. In contrast, oth-ers showed that the first lick (Segal et al., 1965) as well as peak licking (Rosellini & Burdette, 1980) were delayed as a function of IFI length. Rosellini (1985) suggested that the IFI effect demonstrated by Rosellini and Burdette (1980) was due to timing response
which occurred in anticipation of the next
1
This research was supported in part by a Grant-in
Aid for Scientific researches, Japan Ministry of Education, Science and Culture, No.62710078, awarded to the first author. A part of this study was reported at the 52nd Annual Convention of the Japanese Psychological Association in October 1988.
2
Correspondence and requests for reprint should
be sent to Yoshihisa Hamamura, Department of Liberal Arts, National Defence Academy, 1-10-20 Hashirimizu, Yokosuka, Kanagawa 239, Japan.
Temporal distribution of drinking in VT schedule-induced drinking 241
food delivery. According to this view, the
discrepancy among the findings in the previ-ous studies is readily explained in terms of whether such timing responses occurred or did not.
In the present study, we analysed
tempo-ral distribution of drinking under a single
variable time (VT) schedule. Because the
animals are unable to anticipate the interval to the next coming food delivery under VT schedules, it is possible to eliminate, if any, the effect of anticipatory "timing" responses.
According to Staddon (1977), schedule-induced behaviors such as drinking under a time schedules typically occur early in the
IFI intervals. In contrast, facultative
behav-iors such as running response occur in the
middle of intervals, followed by terminal
behaviors such as food anticipation response at the end of intervals. Temporal distribution of drinking showed its close relation to the occurrence of facultative responses. So, both drinking and running response were recorded and analyzed in the present study.
Method Subjects.
The subjects were 12 male Wistar albino rats (Nippon BIO-SUPP Center), approxi-mately 22 weeks of age at the time of the experiment. They were housed individually
in a wirenetting cage with a 12:12 hr
day/night cycle and maintained at 80 % of
their free-feeding weights by postsession
feeding (Oriental Yeast: MF). Water was
freely available at all times in the home cage. Apparatus
Two running wheels (Muromachi-Kikai: 30 cm in diameter and 10 cm wide) were used. Each running wheel was equipped
with an adjoining cage measuring 11×18×
11.5 cm. A food cup was located on the left side of the adjoining cage and a stainless steel water spout on the right side. Food
pel-lets (Muromachi-Kikai: 50 mg) were
deliv-ered to the food cup by an automatic pellet dispenser (BRS/LVE 114-20). A
stainless-steel water spout was positioned 2 mm
behind a 20-mm-diam access hole, which was 8.6 cm above floor level. Contact with the tip of the drinking spout was sensed by a
contact-relay drinkometer. The running
wheel was adjusted so as to rotate in both directions and a running response was electri-cally recorded by a magnetic switch every half turn of the wheel. A microcomputer (NEC PC-9801UV2) controlled the experi-mental events.
Procedure
On Day 1-3, each subject was habituated to the running wheel for 10 min. On Day
4-5, baseline water intake was recorded.
Twenty-one pellets were massed in the food cup at the start of the session, and each rat was allowed to remain in the box for 42 min. The number of licking responses and the number of running responses per second were recorded as the baseline water-intake and the baseline wheel-running, respectively. On Day 6-35, the rats were exposed to
thirty 42-min-sessions. In each session, 21
food pellets were delivered according to vari-able time (VT) 112.5 s schedule. VT 112.5 s schedule was composed of four different IFIs (30, 60, 120 and 240 s), and the sequence of IFIs was randomly varied from session to
session. For each session, the number of
licking responses and the number of running responses were recorded every second.
Results
One animal did not develop schedule-induced drinking and thus was excluded from the following analyses. The number of lick-ing and runnlick-ing responses of the remainlick-ing 11 animals steadily increased over sessions, and reached asymptotic levels after 20
ses-Y. Hamamura, T. Okada and K. Yamada
sions (Day 26-35) were used for analyses. The animals began to drink shortly after each food delivery and stopped drinking
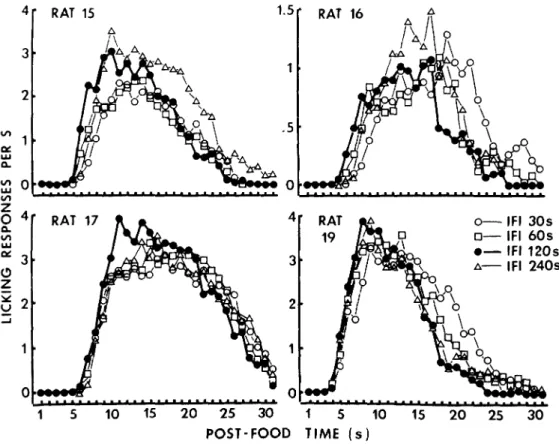
within the first 30 s of the IFIs. Figure 1
shows the typical temporal distribution of
licking over the first 30 s of the IFIs for four conditions of the preceding IFI length in four rats. Since the data shown in Figure 1 and those from the remaining rats suggested the IFI effect, temporal distribution of licking was analyzed in more details as follows.
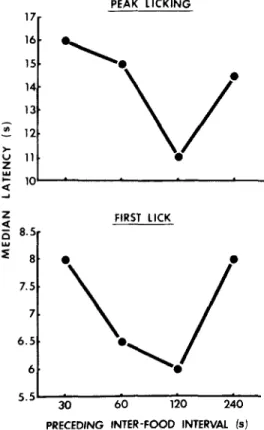
The upper panel of Figure 2 shows the median latency of peak licking (time between food delivery and peak licking) as a function of the preceding IFI length for 11 subjects. Friedman two-way analysis of variance indi-cated that there was a significant difference
among the 4 preceding IFI conditions
(X2=9.35, df=3, p<.025). Wilcoxon sign-rank test revealed that the latency of peak licking following 120-s IFI was significantly shorter than those following 30-s IF1 (T=1, n=11, p<.0025), 60-s IFI (T=6, n=11, p<.01), and
240-s IFI (T=6.5, n=9, p<.05). The latency
of peak licking following 240-s IFI was sig-nificantly shorter than that following 30-s IFI (T=8, n=10, p<.025).
The lower panel of Figure 2 shows the median licking latency of the first lick for 9 rats. The data from the remaining 2 rats were excluded because their median latencies were longer than 30 s. The median latencies for each of these 9 rats are shown in Figure 3. There was a significant difference among the conditions of preceding IFI length (t=10.5, df=3, p<.025). The licking latency following
Figure t. Mean number of licking responses as a function of post-food time over the first 30 s of the inter-food intervals for 4 conditions of the preceding inter-inter-food interval length.
Temporal distribution of drinking in VT schedule-induced drinking 243
120-s IFI was significantly shorter than that following 30-s IFI (T=0, n=6, p<.025) and 240-s IFI (T=0, n=6, p<.025). The licking latency following 60-s IFI was significantly shorter than that following 30-s IFI (T=0,
n=8, p<.005) and 240-s IFI (T=0, n=5,
p<,05).
As it was shown in Figure 1, the subjects stopped drinking early following 120-s IFI. The median number of licking responses dur-ing the last 4 s of the first 30 s of IFIs (27-30 s after food delivery) for the nine rats was .98 following 30-s IFI, .76 following 60-s IFI, .16 following 120-s IFI, and .54 following 240-s IFI. There was a significant difference among the four IFI conditions (X1=8.2, df=3,
p<.05). The number of licking responses
during this period following 120-s IFI was
Figure 2. Median latency of the first lick (lower panel) and peak licking (upper panel) as a function of the preceding inter-food interval length.
significantly less than that following 30-s IFI (T=4, n=8, p<.05), 60-s IFI (T=7, n=9, p<.05), and 240-s IFI (T=4, n=8, (p<.025).
Length of IFIs did affect the temporal pattern of drinking, but did not the number of licking. The group median of the mean num-ber of licking responses within the first 30 s after pellet delivery was 30.0 following 30-s
I FI, 28.7 following 60-s IFI, 30.6 following 120-s IFI, and 33.3 following 240-s IFI. There was no significant difference among the IFI conditions.
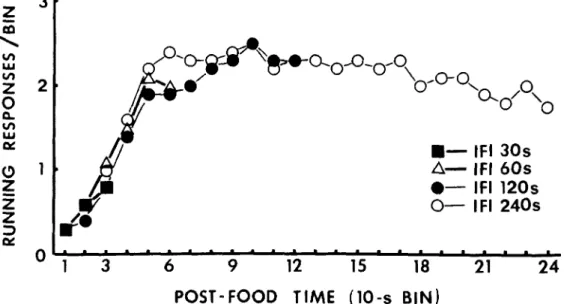
Figure 4 shows the temporal distributions of wheel running responses in the succesive 10-s bins of 30, 60, 120, and 240-s IFIs. A sign test revealed that the number of running responses during the last 10-s bin of 120-s
FI was significantly higher than that during the last 10-s bin of 30-s IFI (p<.01), 60-s IFI (p<.05), and 240-s IFI (p<.05).
No specific correlation was to be found between the number of running responses during the last 10-s bin of the IFIs and the licking latency in the following interval. The median latency of the first lick was 7 s when 0, 1, 2 or 3 wheel running responses occurred during the last 10-s bin of the preceding IFIs; 6 s when 4, 5, 6, 7, 8 or 9 running responses
Figure 3. Median latency of the first lick as a func-tion of the preceding inter-food interval length for individual subjects.
244 Y. Hamamura, T. Okada and K. Yamada
occurred; and 6 s when more than 10 running responses occurred.
Discussion
It was shown that the animals began and stopped drinking earlier following 120-s IFI than following 30, 60, and 240-s IFIs, A U-shaped relation was found between the length of IFI and the latency of immediately follow-ing lickfollow-ing under VT schedule (lower panel of Figure 2). In the previous studies conduct-ed by using FT schedules, IFI effect was explained as due to superstitious (Segal et al., 1965) or timing response (Rosellini, 1985). However, the findings in the present experi-ment cannot be explained by such anticipato-ry responses because animals are unable to expect the time of the next food delivery in the VT schedule.
The present study also showed that the length of IFIs did not affect the number of licking behavior but changed the temporal pattern of licking. This suggests that "the expected length of IFI" available only in FT
schedules and "the length of preceding IFI" might have different effects on drinking. That is, the expected length of the next IFI changes the amount of drinking under FT schedules, but the length of preceding IFI changes the temporal pattern of drinking such as licking latency.
There are at least three possible accounts of licking latency based on which the U-shaped functions shown in Figure 2 may be explained. (1) The length of post-food time has some effects on facultative behaviors such as 'running', which may compete with the immediately following eating and licking behavior, (2) The length of post-food time has some effects on the amount of running, and 'running-induced thirst' may affect the immediately following licking latency. (3) The length of the preceding IFI rather than any facultative behavior conducted by post-food time directly affects the immediately following licking latency.
The first explanation expects a negative correlation between the amount of running and the following licking latency. In the
Figure 4. Mean number of running responses as a function of post-food time (10-s bin) in the food inter-vals of 30, 60, 120 and 240 s.
Temporal distribution of drinking in VT schedule-induced drinking 245
present experiment, the amount of running just before the food delivery was highest in
120-s IFI (Figure 4), whereas the licking
latency was shortest following 120-s IFI
(Figure 2). This finding is not consistent with this explanation.
The second explanation expects short
latencies of licking following the IFIs, during which the rats showed the highest levels of running (the 120-s IFIs in the present study). This was supported by the findings shown in Figures 2 and 4. It was revealed, however, that the difference in the amount of running during the last 10-s bin of IFIs did not pro-duce systematic difference in the latency of following licking, a finding inconsistent with this explanation.
The third explanation seems more consist-ent with the findings in the presconsist-ent study. Then, why does the preceding IFI directly affect the following licking latency so as to
produce the U-shaped function shown in
Figure 2? One possibility is that wheel run-ning and licking are controlled by a common
motivational state (Levitsky & Collier,
1968), and the strength of this motivation is affected by post-food time. Food delivery at the end of interval interrupts wheel running
that has been enhanced by the increased
motivational state. The interruption of
enhanced running may induce the rats to
engage in licking responses. In the present
experiment, the amount of running just
before food delivery was highest in the 120-s IFIs (Figure 4). That is, the motivational state might be highest following the 120-s IFI. This notion readily explains the shortest licking latency in the 120-s IFIs.
The common motivational view is
sup-ported by the findings in the previous studies (Segal, 1969; Roper, 1978) that the amount of drinking increased when running was pre-vented, and by the inverted U-shaped
func-tions obtained between the IFI and the
amount of running (Riley, Wetherington,
Delamater, Peele, & Dacanay, 1985) or drinking (Rosellini & Burdette, 1980; Segal et al., 1965) under FT schedules of different IFI lengths.
Similar inverted U-shaped relations have been observed between the IFI and wood-chewing time under fixed interval schedules in rats (Roper, 1983), and between the IFI and the number of aggressive responses under FT schedules in pigeons (Flory, 1969). Similar relation was also observed between the meal size and the amount of drinking in rats (Reid & Staddon, 1987), with the num-ber of drinking responses being maximal under intermediate food size. These findings consistently support the notion of common motivational state.
Another possible explanation as to why the preceding IFI directly affects the follow-ing lickfollow-ing latency is that animals expect averaged length of IFIs and then be surprised by unexpected food deliveries in the longer or shorter IFIs. Unexpected food deliveries delay the following licking. In this case, the U-shaped function does not reflect the moti-vational state of drinking. It should be noted that this 'surprise' explanation does not pre-dict the U-shaped function under FT sched-ules of different IFI lengths. This explanation predicts the shortest licking latency following the IFI of average length under VT sched-ules, and not always following the 120-s IFIs regardless of the VT values. Further studies are needed to assess the underlying process of the U-shaped function obtained under a VT schedule in the present study.
References
Cohen, P.S., Looney, T.A., Carnpagnoni, F.R., & Lawler, C.P. 1985 A two-state model of rein-forcer-induced motivation. In F.R. Brush, & J.B. Overmier (Eds.), Affect, conditioning, and cog-nition: Essays on the determinants of behavior. Hillsdale, NJ: Lawrence Erlbaum Associates. Pp. 281-297.
246 Y. Hamamura, T. Okada and K. Yamada
Falk, J.L. 1961 Production of polydipsia in normal rats by an intermittent food schedule. Science,
133, 195-196.
Falk, J.L. 1966 Schedule-induced polydipsia as a function of lived interval length. Journal of the
Experimental Analysis of Behavior, 9, 37-39. Falk, J. L. 1969 Conditions producing psychogenic
polydipsia in animals. Annuals of the New York Academy of Science, 157, 569-593.
Flory, R.K. 1969 Attack behavior as a function of minimum inter-food interval. Journal of the Experimental Analysis of Behavior, 12, 825-828. Levitsky, D., & Collier, G. 1968 Schedule-induced wheel running. Physiology and Behavior, 3, 571-573.
Reid, A.K., & Staddon, J.E.R. 1987 Within-session meal-size effects on induced drinking. Journal of the Experimental Analysis of Behavior, 48,
289-301.
Riley, A.L., Wetherington, C.L., Delamater, A.R., Peele, D.B., & Dacanay, R.J. 1985 The effects of variations in the interpellet interval on wheel running in the rat. Animal Learning and Behavior, 13, 201-206.
Roper, T.J. 1978 Diversity and substitutability of adjunctive activities under fixed-interval
sched-ules of food reinforcement. Journal of the Experimental Analysis of Behavior, 30, 83-96. Roper, T.J. 1983 Schedule-induced behavior. In R.L.
Mellgren (Ed.), Animal cognition and behavior, The Netherlands: North Holland. Pp.127-164. Rosellini, R.A. 1985 An application of
opponent-process theory to adjunctive behavior. In F.R. Brush & J.B. Overmier (Eds.), Affect, condition-ing, and cognition: Essays on the determinants of behavior. Hillsdale, NJ: Lawrence Erlbaum Associates. Pp.263-280.
Rosellini, R.A., & Burdette, D.R. 1980 Meal size and intermeal interval both regulate schedule-induced water intake in rats. Animal Learning and Behavior, 8, 647-652.
Segal, E.F., Oden, D.L., & Deadwyler, S.A. 1965 Determinants of polydipsia: IV. Free-reinforce-ment schedule. Psychonomic Science, 3, 11-12. Segal, E.F. 1969 The interaction of psychogenic polydipsia with wheel running in rats. Psychonomic Science, 14, 141-144.
Staddon, J.E.R. 1977 Schedule-induced behavior. In W.K. Honig & J.E.R. Staddon (Eds.), Handbook of operant behavior. Englewood Cliffs, NJ: Prentice-Hall. Pp.125-152.