Rho/myocardin関連転写因子A(MRTF-A)経路は,ヒト網膜色素上皮細胞でTGF-β2による上皮間葉移行において重要な働きをする
14
0
0
全文
(2) Umi Tanaka, et al.. 218. biomarker 8), and produce extracellular matrix 2, 3, 9). 2, 3). 32,34,35). . Thus, TGF-β-induced. .. nuclear accumulation of MRTF-A is Rho-. This transdifferentiation is referred to as the. associated protein kinase (ROCK)-dependent.. components. such as type I collagen. translocation. 10). epithelial-mesenchymal transition (EMT) .. This Rho/MRTF pathway plays an important. The deposition of these extracellular matrices. role in the TGF-β-induced EMT of various. 2, 9). leads to fibrotic membrane formation. . In. PVR, the contraction of these membranes often results in the failure of retinal detachment 1,11-14). surgery. . In AMD, these fibrotic membranes 9,15). cells, such as cardiac. 36). 39). , lung. scleral , and dermal fibroblasts. 37). 40). and lens epithelial cells (LECs). , colonic. 38). and kidney 29). ,. 36). . However,. the role of the Rho/MRTF pathway in the. . Thus,. TGF-β-induced EMT of RPE cells is unknown.. the prevention of EMT in RPE cells has. Y-27632, a Rho kinase inhibitor, inhibited the. therapeutic implications.. EMT of various TGF- β -induced cells because. Transforming growth factor-β (TGF-. it inhibits F-actin stress fiber formation, which. β ) is a major factor during the EMT of. in turn increases G-actin expression32,34), thereby. RPE cells 9,16) and renal tubular 17-19), lens 20,21),. reducing the nuclear translocation of MRTF-A.. cause severe visual disturbances. 18,19,22-24). epithelial cells. Notably,. Although Y-27632 inhibits the induction of COL1. TGF-β induces α-SMA and collagen production. mRNA via TGF-β in RPE cells 33), the role of. and mammary in these cells. 17,22,25,26). . Recent studies have. MRTF activity on ROCK inhibition is unknown.. indicated that myocardin-related transcription. To determine whether the Rho/MRTF. factor A (MRTF-A) plays an important role in. pathway is involved in the EMT of TGF-β -. 21,22,27,28). ,. induced RPE cells, we examined the effects. epithelial cells.. of Y-27632 on α-SMA and type I collagen. MRTF-A is a transcriptional co-activator. expression levels in TGF-β -induced RPE cells,. TGF-β -induced EMT in renal tubular lens. 20,29). , and mammary. 18,19,23,24). that is sequestered in the cytoplasm. 30). ,. and has been recently reported to play an important role in the EMT of RPE cells. 31). after which MRTF-A localization was linked to said expression levels.. .. Upon TGF-β stimulation, MRTF-A trans-. II. Materials and methods. locates from the cytoplasm into the nucleus. ARPE-19 cell culture: Human retinal. and interacts with the serum response factor. pigmented epithelium cells (ARPE-19). 32). and. were purchased from the American Type. type I collagen . The localization of MRTF-A. Culture Collection (Rockville, MD, USA). To. is controlled by monomeric globular actin. authenticate the cell line, a short tandem. (SRF) to promote the expression of α-SMA 33). 32,34,35). . Decreased cytoplasmic G-actin. repeat (STR) analysis was performed by. expression promotes G-actin dissociation from. the BEX Co., Ltd., (Tokyo, Japan) using the. MRTF-A, which translocates MRTF-A into. GenePrint 10 System (Promega, Madison,. (G-actin). 32,34,35). . TGF-β promotes filamentous. WI, USA). ARPE-19 cells were cultured in. actin (F-actin) stress fiber formation via Rho. Dulbecco’ s modified Eagle’ s medium (DMEM). the nucleus. 32). . This decreases cytoplasmic. (Nacalai Tesque, Inc., Kyoto, Japan) containing. G-actin and promotes MRTF-A nuclear. 10% (v/v) fetal bovine serum (FBS) (GE. activation.
(3) Original: Rho/ MRTF-A pathway in EMT in RPE cells. 219. Table 1. Primer sets Protein. Oligonucleotide. Expected PCR product size. human smooth muscle actin. Sense ATTGCCGACCGAATGCAGAAG Antisense AGAGGCCAGGATGGAGCCAC. 120bp. human collagen type 1A. Sense TGGACCAGCAGACTGGCAAC Antisense TCGTGCAGCCATCGACAGTGGAC. 120bp. GAPDH. Sense AGGTCATCATCCATGACAACTTTG Antisense TTCAGCTCAGGGATGACCTT. 190bp. Healthcare Life Sciences, South Logan, UT,. RNA preparation and RT-PCR analysis:. USA) in a humidified atmosphere of 5% CO2 at. Total RNA from ARPE-19 cells was prepared. 37 ℃. Confluent ARPE-19 cells were cultured. using ISOGEN reagent (NIPPON GENE,. in DMEM containing 0% FBS. Then, the cells. Toyama, Japan). cDNAs were prepared from. were treated with TGF-β2 (12.5 ng/ml; Sigma,. 1 µg of total RNA using PrimeScript reverse. St. Louis, MO, USA) for 24 h. CGG-203971 was. transcriptase (Takara Bio, Kyoto, Japan). dissolved in dimethyl sulfoxide (DMSO) and. according to the manufacturer’s instructions.. diluted with DMEM to 3, 10 or 30 µM (0.1%. PCR amplification was performed using. DMSO). Either 3, 10 or 30 µM CGG-203971. ExTaq polymerase (Takara Bio) for 30 cycles;. or 0.1% DMSO (vehicle) or 3, 10, and 30 µM. each cycle consisted of denaturation at 94 ℃. Y-27632 was added to confluent ARPE-19 cells. for 0.5 min, annealing at 60℃ for 0.5 min, and. 1 h after TGF-β2 treatment.. extension at 72℃ for 1 min. Quantitative real-. siRNA oligonucleotides: The following. time-PCR (qRT-PCR) was performed using the. 21-nucleotide siRNA duplexes were synthesized. EcoTM RT-PCR System (illumina, San Diego,. by Nippon Gene Material Co, Ltd. (Toyama,. CA, USA) according to the manufacturer’s. Japan): 5 ′ -AAGAACATCCTTCCTGTTGAG-. protocol. The primers used for qRT-PCR are. 3 ′for human MRTF a siRNA. The target. listed in Table 1.. sequence for the control Photinus pyralis. Immunocytochemistry: Immunocytochemical. luciferase siRNA was 5-AAGCCATTCTAT. analysis was performed using the anti-. CCTCTAGAG-3 ′ , which has no significant. MRTF-A antibody (ab49311, abcam). All cell-. homology to any mammalian gene sequence. culture media were from Life Technologies,. by BLAST searches.. Inc. An Alexa 488-conjugated anti-rabbit. siRNA transfection: Cells were transfected. antibody and DAPI were obtained from. with siRNA oligonucleotides using the. Molecular Probes (Eugene, OR, USA) and. Lipofectamine 2000 reagent (Invitrogen,. used for nuclear staining. Confluent ARPE-19. Carlsbad, CA, USA) according to the. cells were cultured in DMEM containing 0%. manufacturer’ s protocol and cultured for 48 h. FBS. Then, the cells were treated with TGF-. before subsequent analyses, such as real-time-. β (12.5 ng/ml; Sigma, St. Louis, MO, USA). PCR (RT-PCR) and western blotting.. for 24 h. Leptomycin B (LMB 50 nM) was.
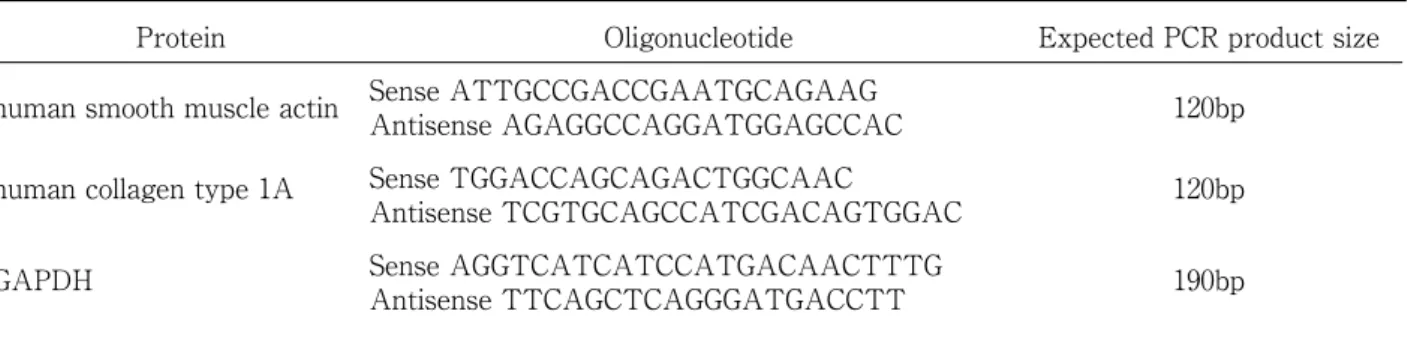
(4) Umi Tanaka, et al.. 220. Relative MRTF-A levels TGF-β2. Protein expression analysis: Sample preparation. **. *. for western blotting, gel preparation, and. **. electrophoretic conditions were performed as described previously 41). Western blot analyses were performed using an anti-MRTF-A antibody (ab49311; Abcam) and anti-GAPDH −. +. siLuc. −. + siMRTF-A. Fig. 1. Effect of MRTF-A siRNA (siMRTF-A) and TGF-β2 on the MRTF-A mRNA expression in ARPE-19 cells (p < 0.0001, ANOVA). Transfection with siMRTF-A decreased MRTF-A mRNA expression in ARPE-19 cells compared with that in control siRNA (siLuc) transfectants. In siLuc transfectants, the expression of MRTF-A mRNA increased slightly by treatment with 12.5 ng/ml TGFβ2. Expression levels were normalized by GAPDH. Data are presented as means ± SEMs of 3 independent experiments. *p < 0.001, ** p < 0.0001 (post-hoc Tukey's multiple comparison tests).. antibody (Chemicon International, Temecula, CA, USA). Band intensity in the immunoblot was semi-quantified using Image J1.38x. Statistics: Data are expressed as means ± standard error of 3 or 6 independent experiments. Statistical analyses were performed using the unpaired Student’ s t-test and one-way analysis of variance (ANOVA) followed by post-hoc Tukey's multiple comparison tests. Statistical tests were performed using Kaleida Graph version 4.1 (Synergy Software, Reading, PA, USA).. III. Results To determine whether MRTF-A plays an added to inhibit nuclear excretion factor for. important role in the EMT of TGF- β -induced. 1 h before cell fixation. After fixation with. ARPE-19 cells, MRTF-A mRNA was knocked. 4% paraformaldehyde solution, cells were. down in the ARPE-19 cells. ARPE-19 cells. incubated with anti-MRTF-A antibody at 4 ℃. transfected with MRTF-A siRNA exhibited. for 6 h. Following incubation with the primary. lower MRTF-A mRNA (Fig. 1) and protein. antibody, the cells were further incubated. expression levels (Fig. 2A, B) than those in. for 1.5 h with Alexa Fluor 488 conjugate goat. control siRNA transfectants (mRNA: 0.55 ±. anti-rabbit secondary antibody (Molecular. 0.04 vs. 1.00 ± 0.03; protein: 0.41 ± 0.05 vs. 1.00. Probes). Immunofluorescent pictures of the. ± 0.00, respectively; p < 0.0001 and p < 0.05).. cells were analyzed using ImageJ software.. TGF-β2 treatment resulted in slightly higher. Integrated intensities were measured from. MRTF-A mRNA and protein expression levels. a specified rectangular area. Measurements. in control siRNA transfectants than those. were obtained from three random cytoplasmic. without TGF-β2 treatment (mRNA: 1.34. sites and the nucleus of each cell, and 300–500. 0.02 vs. 1.00 ± 0.03; protein: 1.58 ± 0.18 vs. 1.00. cells were enumerated per category. The. ± 0.00, respectively; p < 0.001 and p < 0.05).. ±. ratio of the average nuclear and cytoplasmic. To examine whether MRTF-A affects α -. intensities was determined and categorized as. SMA and type I collagen expression, ARPE-. previously reported. 20). .. 19 cells were transfected with control or.
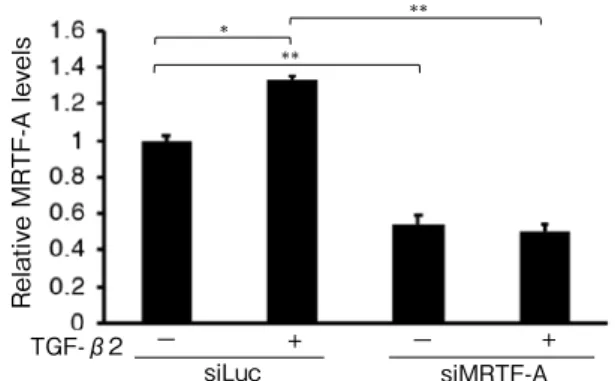
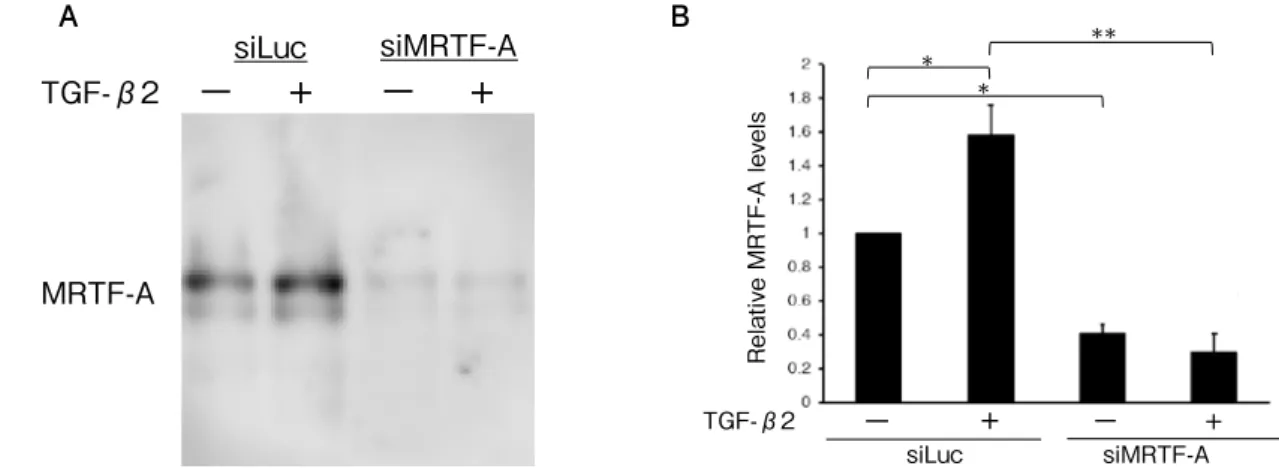
(5) Original: Rho/ MRTF-A pathway in EMT in RPE cells. TGF-β2. MRTF-A. −TGF-β2 +. B. siMRTF-A siLuc. −− +. * *. siMRTF-A. +−. +. MRTF-A. −. TGF-β2. **. **. Relative MRTF-A levels. siLuc. Relative MRTF-A levels. . A. 221. siLuc. +. **. −. +. siMRTF-A. Fig. 2. Effect of MRTF-A siRNA (siMRTF-A) and TGF-β2 TGF-β2on MRTF-A -in ARPE-19 + − protein+expression− cells. siLuc siMRTF-A (A) Transfection with siMRTF decreased MRTF-A protein expression in ARPE-19 cells compared with that in control siRNA (siLuc) transfects by western blotting. In ARPE-19 cells transfected with siLuc, MRTF-A protein expression was increased with treatment of 12.5 ng/ml TGF-β2. (B) Densitometric quantification of MRTF-A/GAPDH (p < 0.05, ANOVA). Densitometric quantification was normalized by GAPDH. Data are presented as means ± SEMs of 3 independent experiments. *p < 0.05, ** p < 0.0001 (post-hoc Tukey's multiple comparison tests). A. * *. B. B Relative Type 1 collagen levels. *. **. Relative Type 1 collagen levels. Relative smooth muscle actin levels. *. Relative smooth muscle actin levels. A. A. *. * *. B *. − − +− − + + + − −+ − + + + TGFTGF TGF-β2 siL siMRT siLuc-β2 siMRTF-A si siMRTF-A siLuc siMR β2 uc F-A Lu TF-A Fig. 3. Effects of MRTF-A on cα -SMA and type I collagen expressions in ARPE-19 cells treated with TGF-β2 −. TGF-β2 (p < 0.0001, p < 0.0001, ANOVA, respectively). ARPE-19 cells transfected with control siRNA (siLuc) or MRTF-A siRNA (siMRTF-A) were cultured with or without 12.5 ng/ml TGF-β2 for 24 h. TGF-β2 treatment increased α -SMA (A) and type I collagen (B) expression in ARPE-19 cells transfected with control siRNA. This increase was partially inhibited by treatment with MRTF-A siRNA. Expression levels were normalized by GAPDH. Data are presented as means ± SEMs of 3 independent experiments. *p < 0.0001, **p < 0.001 (post-hoc Tukey's multiple comparison tests).. MRTF-A siRNA. Then, they were cultured. 0.13 vs. 1.00 ± 0.02; type I collagen expression:. with or without 12.5 ng/ml TGF-β2 for 24 h.. 2.87 ± 0.18 vs. 1.00 ± 0.02, respectively; all. TGF-β2 increased α-SMA and type I collagen. p < 0.0001). These increases were partially. expression levels in ARPE-19 cells transfected. inhibited by treatment with MRTF-A siRNA. with control siRNA ( α -SMA expression: 1.63. (α-SMA expression: 1.10 ± 0.09 vs. 1.63 ± 0.13;.
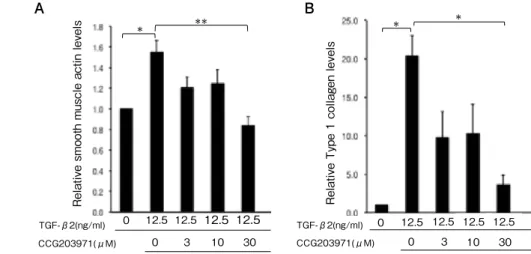
(6) Umi Tanaka, et al.. 222. A. A. CCG203971(μM). *. * Relative Type 1 collagen levels. Relative smooth muscle actin levels TGF-β2(ng/ml). **. *. B. B. 0. 12.5 12.5 12.5 12.5 0. 3. 10. 30. TGF-β2(ng/ml) CCG203971(μM). 0. 12.5 12.5 12.5 12.5 0. 3. 10. 30. Fig. 4. Effect of CCG-203971 on α-SMA and type I collagen expressions in ARPE-19 cells treated with TGF-β2 (p < 0.001, p < 0.0001, ANOVA, respectively). Treatment with 12.5 ng/ml TGF-β2 increased α-SMA (A) and type I collagen (B) expression in ARPE-19 cells, and CCG203971 abolished these increases in a dose-dependent manner. Expression levels were normalized by GAPDH. Data are presented as means ± SEMs of 3 independent experiments. *p < 0.05, **p < 0.001 (post-hoc Tukey's multiple comparison tests).. type I collagen expression: 1.62 ± 0.18 vs. 2.87. To examine whether TGF-β2 promotes the. ± 0.18, respectively; p < 0.001 and p < 0.0001,. nuclear translocation of MRTF-A and CCG-. Fig. 3A, B).. 203971 inhibits this promotion, ARPE-19 cells. To determine whether CCG-203971, a. were cultured with or without 12.5 ng/ml TGF-. MRTF-A inhibitor, could also reduce the. β2 or a combination of TGF-β2 and 30 µM. expression of α-SMA and type I collagen,. CCG-203971 for 24 h. Thereafter, the cells were. we cultured TGF-β2-treated (12.5 ng/ml). treated with LMB, a nuclear export inhibitor,. ARPE-19 cells in the presence or absence. for 1 h. MRTF-A remained in the cytosol. of CCG-203971 (3, 10, and 30 µM). Although. without TGF-β2 treatment (Fig. 5A) but was. treatment with TGF-β2 increased expressions. translocated to the nucleus after TGF-β 2. of α-SMA and type I collagen in ARPE-19. treatment (Fig. 5B). However, joint treatment. cells relative to the control group (α-SMA. with TGF-β2 and 30 µM CCG-203971 inhibited. expression: 1.55 ± 0.11 vs. 1.00 ± 0.00; type. this nuclear translocation (Fig. 5C).. I collagen expression: 20.38 ± 2.63 vs. 1.00. To quantify the changes in MRTF-A. ± 0.00, respectively; all p < 0.01), CCG-. compartmentalization, the fluorescent intensity. 203971 diminished these increases in a dose-. of MRTF-A in the cytoplasm and nucleus. dependent manner ( α -SMA expression: 1.55. was measured. In controls treated only with. ± 0.11, 1.21 ± 0.11, 1.25 ± 0.13, and 0.84 ± 0.09,. LMB, the number of cells with cytoplasmic. for 0, 3, 10, and 30 µM CCG-20397; type I. MRTF-A was 57.0 ± 3.32%; however, this. collagen expression: 20.38 ± 2.63, 9.76 ± 3.40,. number significantly decreased upon TGF-β2. 10.30 ± 3.73, and 3.65 ± 1.21, for 0, 3, 10, and. treatment (12.5 ng/ml) for 24 h to 11.9 ± 1.91%. 30 µM CCG-20397, respectively, Fig. 4A, B).. (p < 0.05). Correspondingly, the number of cells.
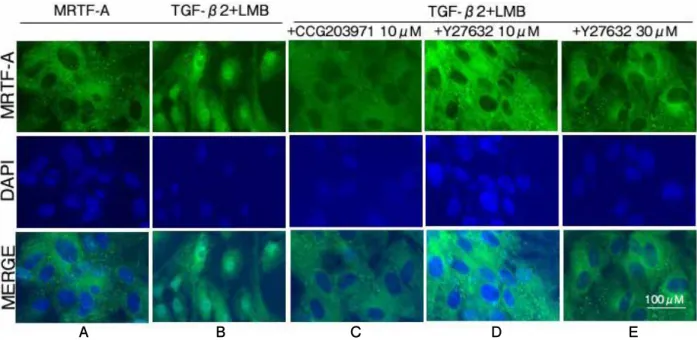
(7) Original: Rho/ MRTF-A pathway in EMT in RPE cells. A. B. C. D. 223. E. Subcellular localization of MRTF-A (%). Fig. 5. Translocation of MRTF-A in ARPE-19 cells stimulated by TGF-β2. ARPE-19 cells were cultured with or without 12.5 ng/ml TGF-β2 for 24 h. Either 30 µM CGG203971 or 10 and 30 µM Y27632 was added 1 h after TGF-β2 treatment. Leptomycin B (LMB 50 nM) was added to inhibit nuclear excretion factor for 10 mins before cell fixation. ARPE-19 cells were immunostained using an MRTF-A-specific antibody (green) and a nuclear stain (DAPI, blue). MRTF-A remained in the cytosol in the absence of TGF-β2 (A), but MRTF-A was translocated to the nucleus after TGF-β2 treatment (B). This nuclear translocation was inhibited by CGG203971 (C). Y27632 also inhibited nuclear translocation of MRTF-A at each concentration (D, E). Scale bar = 100 μm. 100%. *. *. *. *. 80%. *. 60%. カテゴリ 3 Nuclear. * 40%. カテゴリ 2 Pan-cellular カテゴリ 1 Cytoplasmic. *. 20%. * 0%. TGF-β2 (ng/ml). 0. 系列 1. 系列 2 12.5. 系列 3 12.5. 系列 4 12.5. 系列 5 12.5. CCG203971(μM). 0. 30. 0. 0. Y27632 (μM). 0. 0. 10. 30. Fig. 6. Quantification and comparison of intracellular MRTF-A translocation in ARPE-19 cells stimulated by TGF-β2. MRTF-A is primarily cytoplasmic in the control. Nuclear MRTF-A significantly increased after TGF-β2 treatment but significantly decreased upon treatment with 30 µM CCG203971 relative to the TGF-β2-treated cells. Similarly, Y27632 treatment significantly decreased nuclear MRTF-A in a dose-dependent manner. Gene expression levels were normalized to those of GAPDH. Data are presented as the mean ± SEMs of 6 independent experiments. *p < 0.05 (post-hoc Tukey's multiple comparison tests).. with nuclear MRTF-A significantly increased. with 30 µM CCG-203971, this increase was. relative to the control (68.0 ± 3.29% vs. 18.4. significantly attenuated (4.42 ± 1.72% vs. 68.0. ± 1.95%, p < 0.05). However, upon treatment. ± 3.29%, p < 0.05, Fig. 6)..
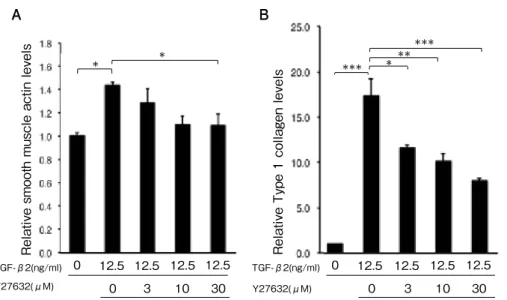
(8) Umi Tanaka, et al.. 224. TGF-β2(ng/ml) Y27632(μM). B *. *. 0. 12.5 12.5 12.5 12.5 0. 3. 10. 30. TGF-β2(ng/ml) Y27632(μM). *. ***. Relative Type 1 collagen levels. Relative smooth muscle actin levels. A. 0. 12.5 0. **. ***. 12.5 12.5 12.5 3. 10. 30. Fig. 7. Effect of Y27632 on α -SMA and type I collagen expressions in ARPE-19 cells treated with TGFβ2 (p < 0.05, p < 0.001, ANOVA, respectively). Treatment with 12.5 ng/ml TGF-β2 increased α-SMA (A) and type I collagen (B) expression in ARPE-19 cells, and Y27632 abolished these increases in a dose-dependent manner. Expression levels were normalized by GAPDH. Data are presented as means ± SEMs of 3 independent experiments. *p < 0.05, **p < 0.01, ***p < 0.001 (post-hoc Tukey's multiple comparison tests).. To determine whether Y-27632, a ROCK. MRTF-A were examined. ARPE-19 cells were. inhibitor, inhibits the EMT of TGF- β. cultured with or without 12.5 ng/ml TGF- β. -induced RPE cells, TGF-β2-treated (12.5 ng/. 2 or a combination of TGF-β2 and 10 or 30µM. ml) ARPE-19 cells were cultured with or. Y-27632 for 24 h. ARPE-19 cells were then. without Y-27632 (3, 10, and 30 µM). Although. treated with LMB for 1 h. MRTF-A remained. treatment with TGF-β2 increased the. in the cytosol without TGF-β2 treatment. expression levels of α-SMA and type I collagen. (Fig. 5A) but was translocated to the nucleus. in ARPE-19 cells relative to the control (α-. after TGF-β2 treatment (Fig. 5B). However,. SMA expression: 1.43 ± 0.03 vs. 1.00 ± 0.02;. treatment with a combination of TGF-β2 and. type I collagen expression: 16.99 ± 1.91 vs. 1.00. 10 or 30µM Y-27632 inhibited this nuclear. ± 0.00, respectively; p < 0.05 and p < 0.0001),. translocation (Fig. 5D, E).. Y-27632 diminished these increases in a dose-. Furthermore, we investigated the effect. dependent manner (α-SMA expression: 1.43 ±. of Y-27632 on the changes in MRTF-A. 0.03, 1.29 ± 0.11, 1.10 ± 0.08 and 1.01 ± 0.09, for. c o m p a r t m e n t a l i z a t i o n . A g a i n , T G F - β2. 0, 3, 10, and 30 µM Y-27632; type I collagen. treatment significantly increased the number. expression: 16.99 ± 1.91, 11.34 ± 0.40, 9.97 ±. of cells with nuclear MRTF-A relative to the. 0.80, and 7.82 ± 0.28, for 0, 3, 10, and 30 µM. control (68.0 ± 3.29% vs. 18.4 ± 1.95%, p < 0.05);. Y-27632, respectively, Fig. 7A, B).. however, Y-27632 treatment (10 and 30 µM). To determine whether the Rho/MRTF. significantly attenuated this increase in a dose-. pathway is involved in the EMT of TGF-. dependent manner (30.7 ± 3.04% and 20.7 ±. β-induced RPE cells, the localizations of. 3.92%, respectively; p < 0.05, Fig. 6)..
(9) Original: Rho/ MRTF-A pathway in EMT in RPE cells. 225. IV. Discussion. bleb injections were required for the first 9. In this study, TGF-β2 led to increases in α-. days post-surgery followed by twice-weekly. SMA and type I collagen mRNA expression. injections for 3 weeks. Kobayashi et al.. in ARPE-19 cells, which were repressed. reported that intravitreal CCG-1423 injection. by MRTF-A knockdown. CCG-203971, an. markedly suppressed the development of. MRTF-A inhibitor, also diminished the. photocoagulation-induced subretinal fibrosis.. promotion of α-SMA and type I collagen. Although the residence time of the drug in. mRNA expressions in TGF-β2-treated ARPE-. the vitreous cavity was longer than usual 44),. 19 cells with a reduction of MRTF-A nuclear. CCG-1423 had to be injected into the vitreous. translocation. Y-27632, a ROCK inhibitor,. cavity twice a week. In this study, Y-27632,. also inhibited α-SMA and type I collagen. a ROCK inhibitor, also inhibited α-SMA and. expression and suppressed the translocation. type I collagen expression and suppressed. of MRTF-A to the nucleus in TGF-β2-treated. the translocation of MRTF-A to the nucleus. ARPE-19 cells. These results suggest that the. in TGF-β2-induced ARPE-19 cells. Moreover,. Rho/MRTF pathway plays an important role. Y-27632 also inhibited α-SMA and type I. in the EMT of TGF-β-induced RPE cells.. collagen expression in TGF-β2-induced LECs. 31). also. Our results also indicated that CCG-203971,. and topical Y-27632 administration prevented. a MRTF-A inhibitor, represses α-SMA and. anterior subcapsular cataract induced by. type I collagen mRNA expression and TGF-. UV-B irradiation, which had been developed. β2-induced MRTF-A nuclear translocation in. by the EMT of LECs 45). A topically instilled. ARPE-19 cells. These results are consistent. radiolabeled ROCK inhibitor can be detected. 31). with recent work by Kobayashi et al. , who. in the retina and choroid up to 4 h later 46).. reported that CCC-1423, a first-generation. Ripasudil, another ROCK inhibitor applied as. MRTF-A inhibitor, repressed the expression. eye drops, was used clinically in Japan for the. of α-SMA and type I collagen mRNA and. treatment of glaucoma 47,48). Although topical. TGF- β2-mediated nuclear translocation of. administration of ripasudil is required twice a. MRTF-A in RPE-1 cells, another cell line of. day via eye drops, this administration method. RPE cells. CCG-203971 is a second-generation. is easier and safer than a subconjunctival or. inhibitor with far less cytotoxicity than CCG-. intravitreal injection, which in rare cases can. 1423. 30,42). . Notably, CCG-222740, a novel MRTF. cause endophthalmitis, a complication that. inhibitor, was recently developed and has. can have devastating visual outcomes. These. exhibited a greater inhibitory effect on α-SMA. findings highlight the promising potential. 43). expression than CCG-203971 .. of topical ROCK inhibitor administration. Local administration of these MRTF-A. as a novel PVR and AMD treatment.. inhibitors prevented ocular fibrosis in vivo.. In this study, MRTF-A knockdown repressed. Particularly, local administration of CCG-. α-SMA mRNA expression induced by TGF-. 222740 prevented subconjunctival scarring. β2 in ARPE-19 cells. MRTF-A in conjunction. in rabbits after in vivo glaucoma filtration. with SRF. surgery. 43). . However, daily subconjunctival. 49,50). can induce α -SMA mRNA. expression after MRTF-A translocation from.
(10) Umi Tanaka, et al.. 226. . In ARPE-. hepatocyte growth factor 22), epidermal growth. 19 cells, TGF-β2 promoted MRTF-A nuclear. factor 1,9,12,15), and other members of the TGF-. transport. Moreover, TGF-β2 increased the. β superfamily such as activin 57) can induce. expression of MRTF-A mRNA and protein in. EMT in RPE cells. Vascular endothelial. ARPE-cells. In MS-1 endothelial cells, TGF-. growth factor (VEGF) is a key factor in. β promoted α -SMA expression not only by. choroidal neovascularization, which is a main. stimulating the nuclear transport of MRTF-A. pathological AMD lesion 5,58,59). VEGF activates. but also by enhancing MRTF-A transcription,. the nuclear translocation of MRTF-A in mouse. which promoted MRTF-A accumulation in. retinal endothelial cells 60), while PDGF post-. the nucleus 51). Further research is required. transcriptionally activates MRTF-A expression. to determine the effect of enhanced MRTF-A. in cultured hepatic stellate cells 61). Therefore,. transcription on TGF-β2-induced α -SMA. cells and growth factors other than RPE cells. expression in ARPE-19 cells.. and TGF-β may potentially play critical roles. the cytoplasm to the nucleus. 32). In this study, the Rho/MRTF pathway was. in PVR or AMD. Further investigations using. found to play an important role in the EMT. in vivo models are required to elucidate the. of TGF-β-induced RPE cells. However, Smad. role of MRTF-A in these diseases.. pathways are canonical TGF-β signaling. In conclusion, our results demonstrate that. pathways and play important roles in. TGF-β2 promoted the expression of α-SMA. 52,53). EMT. . Its cross-talk with the Rho/MRTF. and type I collagen in ARPE-19 cells via the. pathway has also been shown to be critical. Rho/MRTF pathway. These results suggest. during TGF-β induced EMT. TGF-β-induced. that MRTF-A may be a critical mediator of. translocation of MRTF-A in LECs was. EMT in human RPE cells.. recently found to be inhibited in the presence. . of SIS3, a Smad3-specific inhibitor. 54). . Such. cross-talk with the Rho/MRTF pathway may also play an important role in the EMT of RPE cells. In this study, we examined the activity of MRTF-A in TGF-β2-treated ARPE-19 cells. Morphological and histochemical studies of proliferative membranes in PVR 2,3,12,13,55) and AMD. 5,9,15,56). showed that a variety of cells,. such as RPE cells, fibroblasts, glial cells, and inflammatory cells, participate in the pathogenesis of these diseases. Although TGFβ plays a critical role in the EMT of RPE cells. 1,22). , various growth factors, such as. connective tissue growth factor 9,13), plateletderived growth factor (PDGF) 1,9,12,13,15,22) ,. Acknowledgements We thank Ms. Rieko Higashio for technical assistance with the real-time PCR analysis. This work was supported by a Japan Society for the Promotion of Science KAKENHI grant (no. 17K11465). Conflict of interest: The authors have no conflict of interest to declare.. .
(11) Original: Rho/ MRTF-A pathway in EMT in RPE cells. 227. References 1) Shu DY and Lovicu FJ: Myofibroblast transdifferentiation: The dark force in ocular wound healing and fibrosis. Prog Retin Eye Res 60, 44-65, 2017. 2) Hiscott P, Hagan S, Heathcote S, et al.: Pathobiology of epiretinal and subretinal membranes: possible roles for the matricellular proteins thrombospondin 1 and osteonectin (SPARC). Eye 16, 393-403, 2002. 3) Hiscott P, Sheridan C, Magee RM, et al.: Matrix and the retinal pigment epithelium in proliferative retinal disease. Prog Retin Eye Res 18, 167-190, 1999. 4) Hirasawa M, Noda K, Noda S, et al.: Transcriptional factors associated with epithelial-mesenchymal transition in choroidal neovascularization. Mol Vis 17, 1222-1230, 2011. 5) Lopez PF, Sippy BD, Lambert HM, et al.: Transdifferentiated retinal pigment epithelial cells are immunoreactive for vascular endothelial growth factor in surgically excised agerelated macular degeneration-related choroidal neovascular membranes. Invest Ophthalmol Vis Sci 37, 855-868, 1999. 6) Watanabe D, Takagi H, Suzuma K, et al.: Expression of connective tissue growth factor and its potential role in choroidal neovascularization. Retina 25, 911-918, 2005. 7) Feist RM Jr, King JL, Morris R, et al.: Myofibroblast and extracellular matrix origins in proliferative vitreoretinopathy. Graefes Arch Clin Exp Ophthalmol 252, 347-357. 2014. 8) Sappino AP, Schürch W and Gabbiani G: Differentiation repertoire of fibroblastic cells: expression of cytoskeletal proteins as marker of phenotypic modulations. Lab Invest 63, 144-161, 1990. 9) Little K, Ma JH, Yang N, et al.: Myofibroblasts in macular fibrosis secondary to neovascular age-related macular degeneration-the potential sources and molecular cues for their recruitment and activation. EBioMedicine 38, 283-291, 2018. 10) Yang S, Li H, Li M, et al.: Mechanisms of epithelial-mesenchymal transition in proliferative vitreoretinopathy. Discov Med 20, 207-217, 2015. 11) Chen Z, Shao Y and Li X: The roles of signaling pathways in epithelial-to-mesenchymal transition of PVR. Mol Vis 21, 706-710, 2015. 12) Khan MA, Brady CJ and Kaiser RS: Clinical management of proliferative vitreoretinopathy: an. update. Retina 35, 165-175, 2015. 13) Pennock S, Haddock LJ, Eliott D, et al.: Is neutralizing vitreal growth factors a viable strategy to prevent proliferative vitreoretinopathy? Prog Retin Eye Res 40, 16-34, 2014. 14) Sadaka A and Giuliari GP: Proliferative vitreoretinopathy: current and emerging treatments. Clin Ophthalmol 6, 1325-1333, 2012. 15) Ishikawa K, Kannan R and Hinton DR: Molecular mechanisms of subretinal fibrosis in age-related macular degeneration. Exp Eye Res 142, 19-25, 2016. 16) Saika S: TGFb pathobiology in the eye. Lab Invest 86, 106-115, 2006. 17) Cao Y, Hu J, Sui J, et al.: Quercetin is able to alleviate TGF-β-induced fibrosis in renal tubular epithelial cells by suppressing miR-21. Exp Ther Med 16, 2442-2448, 2018. 18) O'Connor JW, Riley PN, Nalluri SM, et al.: Matrix rigidity mediates TGFβ1-induced epithelial-myofibroblast transition by controlling cytoskeletal organization and MRTF-A localization. J Cell Physiol 230, 1829-1839, 2015. 19) O'Connor JW and Gomez EW: Cell adhesion and shape regulate TGF-beta1-induced epithelialmyofibroblast transition via MRTF-A signaling. PLoS One 8, e83188, doi: 10.1371/journal.pone. 0083188, 2013. 20) Gupta M, Korol A and West-Mays JA: Nuclear translocation of myocardin-related transcription factor-A during transforming growth factor betainduced epithelial to mesenchymal transition of lens epithelial cells. Mol Vis 19, 1017-1028, 2013. 21) Lovicu FJ, Shin EH and McAvoy JW: Fibrosis in the lens. Sprouty regulation of TGFβ-signaling prevents lens EMT leading to cataract. Exp Eye Res 142, 92-101, 2016. 22) Chen Q, Yang W, Wang X, et al.: TGF-β 1 induces EMT in Bovine mammary epithelial cells through the TGF-β1/Smad signaling pathway. Cell Physiol Biochem 43, 82-93. 2017. 23) Morita T and Hayashi K: Tumor progression is mediated by thymosin-β4 through a TGFβ / MRTF signaling axis. Mol Cancer Res 16, 880893, 2018. 24) O’ Connor JW, Mistry K, Detweiler D, et al.: Cell-cell contact and matrix adhesion promote α SMA expression during TGF-β 1-induced epithelial-myofibroblast transition via Notch.
(12) 228. Umi Tanaka, et al.. and MRTF-A. Sci Rep 6, 26226, doi: 10.1038/ srep26226, 2016. 25) Urakami C, Kurosaka D, Tamada K, et al.: Lovastatin alters TGF-β-induced epithelialmesenchymal transition in porcine lens epithelial cells. Curr Eye Res 37, 479-485, 2012. 26) Xiao W, Chen X, Liu X, et al.: Trichostatin A, a histone deacetylase inhibitor, suppresses proliferation and epithelial-mesenchymal transition in retinal pigment epithelium cells. J Cell Mol Med 18, 646-655, 2014. 27) Morita T, Mayanagi T and Sobue K: Dual roles of myocardin-related transcription factors in epithelial–mesenchymal transition via slug induction and actin remodeling. J Cell Biol 179, 1027-1042. 2007. 28) Xu H, Wu X, Qin H, et al.: Myocardin-related transcription factor A epigenetically regulates renal fibrosis in diabetic nephropathy. J Am Soc Nephrol 26, 1648-1660, 2015. 29) Korol A, Taiyab A and West-Mays JA: RhoA/ ROCK signaling regulates TGF β -induced epithelial-mesenchymal transition of lens epithelial cells through MRTF-A. Mol Med 22, 713-723, 2016. 30) Watanabe B, Minami S, Ishida H, et al.: Stereospecific inhibitory effects of CCG-1423 on the cellular events mediated by myocardinrelated transcription factor A. PLoS One 10, e0136242, doi: 10.1371/journal.pone.0136242, 2015. 31) Kobayashi M, Tokuda K, Kobayashi Y, et al.: Suppression of epithelial-mesenchymal transition in retinal pigment epithelial cells by an MRTF-A inhibitor. Invest Ophthalmol Vis Sci 60, 528-537, 2019. 32) Olson EN and Nordheim A: Linking actin dynamics and gene transcription to drive cellular motile functions. Nat Rev Mol Cell Biol 11, 353365, 2010. 33) Itoh Y, Kimoto K, Imaizumi M, et al.: Inhibition of RhoA/Rho-kinase pathway suppresses the expression of type Icollagen induced by TGFbeta2 in human retinal pigment epithelial cells. Exp Eye Res 84, 464-472, 2007. 34) Tsou PS, Haak AJ, Khanna D, et al.: Cellular mechanisms of tissue fibrosis. 8. Current and future drug targets in fibrosis: focus on Rho GTPase-regulated gene transcription. Am J Physiol Cell Physiol 307, 2-13, 2014. 35) Yu-Wai-Man C, Treisman R, Bailly M, et al.: The role of the MRTF-A/SRF pathway in ocular. fibrosis. Invest. Ophthalmol Vis Sci 55, 4560-4567, 2014. 36) Small EM: The Actin-MRTF-SRF Gene Regulatory Axis and Myofibroblast. J Cardiovasc Trans Res 5, 794-804, 2012. 37) Sandbo N, Lau A, Kach J, et al.: Delayed stress fiber formation mediates pulmonary myofibroblast differentiation in response to TGFβ. Am J Physiol Lung Cell Mol Physiol 301, 656666, 2011. 38) Laura AJ, Eva SR, Andrew JH, et al.: Novel Rho/MRTF/SRF inhibitors block matrix-stiffness and TGF-β -induced fibrogenesis in human colonic myofibroblasts. Inflamm Bowel Dis 20, 154-165, 2014. 39) Yuan Y, Li M, To CH, et al.: The role of the RhoA/ROCK signaling pathway in mechanical strain-induced scleral myofibroblast differentiation. IOVS 59, 3619-3629, 2018. 40) Akhmetshina A, Dees C, Pileckyte M, et al.: Rho-associated kinases are crucial for myofibroblast differentiation and production of extracellular matrix in scleroderma fibroblasts. Arthritis Rheum 58, 2553-2564, 2008. 41) Ogasawara S, Hashizume K, Okuno T, et al.: Effect of geranylgeranylacetone on ultraviolet radiation type B-induced cataract in heat shock transcription factor 1 heterozygous mouse. Curr Eye Res 42, 732-737, 2017. 42) Bell JL, Haak AJ, Wade SM, et al.: Optimization of novel nipecotic bis(amide) inhibitors of the Rho/ MKL1/SRF transcriptional pathway as potential anti-metastasis agents. Bioorg Med Chem Lett 23, 3826-3832, 2013. 43) Tagalakis AD, Madaan S, Larsen SD, et al.: In vitro and in vivo delivery of a sustained release nanocarrier-based formulation of an MRTF/SRF inhibitor in conjunctival fibrosis. J Nanobiotechnology 16, doi: 10.1186/s12951-0180425-32018, 2018. 44) García-Quintanilla L, Luaces-Rodríguez A, Gil-Martínez M, et al.: Pharmacokinetics of intravitreal anti-VEGF drugs in age-related macular degeneration. Pharmaceutics 31, 11, 8, doi: 10.3390/pharmaceutics11080365, 2019. 45) Imaizumi T, Kurosaka D, Tanaka U, et al.: Topical administration of a ROCK inhibitor prevents anterior subcapsular cataract induced by UV-B irradiation. Exp Eye Res 181, 145-149, 2019. 46) Isobe T, Mizuno K, Kaneko Y, et al.: Effects of.
(13) Original: Rho/ MRTF-A pathway in EMT in RPE cells. K-115, a rho-kinase inhibitor, on aqueous humor dynamics in rabbits. Curr Eye Res 39, 813-822, 2014. 47) Garnock-Jones KP: Ripasudil: first global approval. Drugs 74, 2211-2215, 2014. 48) Terao E, Nakakura S, Fujisawa Y, et al.: Time course of conjunctival hyperemia induced by a Rho-kinase inhibitor anti-glaucoma eye drop: ripasudil 0.4%. Curr Eye Res 42, 738-742, 2017. 49) Esnault C, Stewart A, Gualdrini F, et al.: Rho-actin signaling to the MRTF coactivators dominates the immediate transcriptional response to serum in fibroblasts. Genes Dev 28, 943-958, 2014. 50) Wang DZ, Li S, Hockemeyer D, et al.: Potentiation of serum response factor activity by a family of myocardin-related transcription factors. Proc Natl Acad Sci USA 99, 14855-14860, 2002. 51) Mihira H, Suzuki HI, Akatsu Y, et al.: TGFβ -induced mesenchymal transition of MS-1 endothelial cells requires Smad-dependent cooperative activation of Rho signals and MRTF-A. J Biochem 151, 145-156, 2013. 52) Saika S, Yamanaka O, Okada Y, et al.: TGF β in fibroproliferative diseases in the eye. Front Biosci (Schol Ed) 1, 376-390, 2009. 53) Flanders KC: Smad3 as a mediator of the fibrotic response. Int J Exp Pathol 85, 47-64, 2004. 54) Taiyab A, Holms J and West-Mays JA: β Catenin/Smad3 interaction regulates transforming growth factor-β -induced epithelial to mesenchymal transition in the lens. Int J Mol. 229. Sci 20, 9, doi: 10.3390/ijms20092078, 2019. 55) Oberstein SY, Byun J, Herrera D, et al.: Cell proliferation in human epiretinal membranes: characterization of cell types and correlation with disease condition and duration. Mol Vis 17, 17941805, 2011. 56) Grossniklaus HE, Hutchinson AK, Capone A Jr, et al.: Clinicopathologic features of surgically excised choroidal neovascular membranes. Ophthalmology 101, 1099-1111, 1994. 57) Saika S, Kono-Saika S, Tanaka T, et al.: Smad3 is required for dedifferentiation of retinal pigment epithelium following retinal detachment in mice. Lab Invest 84, 1245-1258, 2004. 58) Kvanta A, Algvere PV, Berglin L, et al.: Subfoveal fibrovascular membranes in agerelated macular degeneration express vascular endothelial growth factor. Invest Ophthalmol Vis Sci 37, 1929-1934, 1996. 59) Miller JW, Le Couter J, Strauss EC, et al.: Vascular endothelial growth factor a in intraocular vascular disease. Ophthalmology 120, 106-114, 2013. 60) Weinl C, Riehle H, Park D, et al.: Endothelial SRF/MRTF ablation causes vascular disease phenotypes in murine retinae. J Clin Invest 123, 2193-2206, 2013. 61) Han X, Hao C, Li L, et al.: HDAC4 stimulates MRTF-A expression and drives fibrogenesis in hepatic stellate cells by targeting miR-206. Oncotarget 29, 47586-47594, 2017..
(14) 岩手医誌 72 巻,5 号(令和 2 年 12 月)217-230 頁.. Rho/myocardin 関連転写因子 A(MRTF-A)経路は, ヒト網膜色素上皮細胞で TGF-β2 による上皮間葉移行において重要な働きをする 田中うみ 1),黒坂大次郎 1), 村井憲一 1),橋爪公平 1),福田一央 1), 五日市そら 1),三部 篤 2) 1). 2). 岩手医科大学医学部,眼科学講座 岩手医科大学薬学部,病態薬理学講座薬剤治療学分野. (Received on January 20, 2020 & Accepted on February 18, 2020). 要旨 網膜色素上皮細胞( RPE )における上皮間葉移行 ( EMT )は,増殖硝子体網膜症や滲出型加齢黄斑変性 に関与する.トランスフォーミング増殖因子β( TGFβ)刺激により,Rho 活性化を介し myocardin 関連 転写因子 A( MRTF-A )が細胞質から核内に移行し EMT が生じるが,RPE における Rho/MRTF 経路の 働きは知られていない.本研究で,TGF-β2 はヒト RPE( ARPE- 19)において EMT の指標であるα平滑. 筋アクチン(α-SMA )と I 型コラーゲン発現を促進 し,これは MRTF のノックダウンにより阻害された. MRTF 阻害薬である CCG- 203971 は MRTF-A の核内 移行を阻害し,両者の発現を阻害した.Rho 関連タ ンパクキナーゼ阻害薬である Y- 27632 も,同様に核 内移行と両者の発現を阻害した.Rho/MRTF 経路が TGF-β2 による RPE での EMT で重要な役割を果た すことが示唆された.. 230.
(15)
図
+3
関連したドキュメント
However, VPA prevented the morphological changes characteristic for activation and inhibited the expres- sion of collagen type 1 α 1 (COL1A1) and TGF- β 1 in activated LI90 cells
Treatment with ONO-1301 increased hepatic HGF mRNA expression, but decreased the expressions of TGF-β1, connective tissue growth factor, α-smooth muscle actin, and type-I and
内輪面の凹凸はED注射群程ではないが,粘膜上皮の
Standard domino tableaux have already been considered by many authors [33], [6], [34], [8], [1], but, to the best of our knowledge, the expression of the
An example of a database state in the lextensive category of finite sets, for the EA sketch of our school data specification is provided by any database which models the
尿路上皮癌、肉腫様 Urothelial carcinoma, sarcomatoid subtype 8122/3 尿路上皮癌、巨細胞 Urothelial carcinoma, giant cell subtype 8031/3 尿路上皮癌、低分化
We present sufficient conditions for the existence of solutions to Neu- mann and periodic boundary-value problems for some class of quasilinear ordinary differential equations.. We
Replace the previous sum by a sum over all partitions in S c × DD Check that coefficents of x n on both sides are polynomials in t, and conclude that the formula is true for
