ContentslistsavailableatScienceDirect
Journal of Neuroscience Methods
j o ur na l h o me p a g e :w w w . e l s e v i e r . c o m / l o c a t e / j n e u m e t h
Generation of a neurodegenerative disease mouse model using lentiviral vectors carrying an enhanced synapsin I promoter
Yasunori Matsuzaki, Miho Oue, Hirokazu Hirai
∗DepartmentofNeurophysiology,GunmaUniversityGraduateSchoolofMedicine,Maebashi,Gunma371-8511,Japan
h i g h l i g h t s
•Wedevelopedanewhybridpromoterhavinghighneuronalspecificity.
•Thepromotershowedefficientandspecificneuronaltransductioninthecerebellum.
•Spinocerebellarataxiamodelmicewerelentivirallygeneratedusingthepromoter.
•Themodelmiceshowedneuron-specifictransgeneexpressionthroughouttheCNS.
•Themodelmiceshowedataxiaandneuronalinclusionscharacteristictothedisease.
a r t i c l e i n f o
Articlehistory:
Received11July2013 Receivedinrevisedform 26November2013 Accepted5December2013
Keywords:
SynapsinIpromoter Cerebellum Lentiviralvector Transgenicmouse Spinocerebellarataxia Ataxin-1
a b s t r a c t
Background:Certaininheritedprogressiveneurodegenerativedisorders,suchasspinocerebellarataxia (SCA),affectneuronsinlargeareasofthecentralnervoussystem(CNS).Theselectiveexpressionof disease-causingandtherapeuticgenesinsusceptibleregionsandcelltypesiscriticalforthegenerationof animalmodelsanddevelopmentofgenetherapiesforthesediseases.Previousstudieshavedemonstrated theadvantagesoftheshortsynapsinI(SynI)promoter(0.5kb)asaneuron-specificpromoterforrobust transgeneexpression.However,theshortSynIpromoterhasalsoshownsomepromoteractivityinglia andalackoftransgeneexpressioninsignificantareasoftheCNS.
Newmethods:ToimprovetheSynIpromoter,weusedaSynIpromoterthatistwiceaslong(1.0kb) astheshortSynIpromoterandincorporatedaminimalCMV(minCMV)sequence.
Results:Weobservedthatthe1.0kbratSynIpromoterwithminCMV[rSynI(1.0)-minCMV]exhibited robustpromoterstrength,excellentneuronalspecificityandwide-rangingtransgeneexpressionthrough- outtheCNS.
Comparisonwithexistingmethods:Comparedwiththetwopreviouslyreportedshort(0.5kb)promo- ters,thenewpromoterwassuperiorwithrespecttoneuronalspecificityandmoreefficientlytransduced neurons.Moreover,transgenicmiceexpressingthemutantproteinATXN1[Q98],whichcausesSCAtype 1(SCA1),underthecontroloftherSynI(1.0)-minCMVpromotershowedrobusttransgeneexpression specificallyinneuronsthroughouttheCNSandexhibitedprogressiveataxia.
Conclusion:rSynI(1.0)-minCMVdrivesrobustandneuron-specifictransgeneexpressionthroughoutthe CNSandisthereforeusefulforviralvector-mediatedneuron-specificgenedeliveryandgenerationof neuron-specifictransgenicanimals.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Previous studies comparing several neuron-specific promo- tershave shownthatthesynapsinI(SynI)promoterissuperior in terms of promoter strengthand neuronal specificity (Boulos et al., 2006; Hioki et al., 2007; Kugler et al., 2003; Nakagawa etal.,2006).BecausetheSynIpromoterisphylogeneticallywell
∗Correspondingauthor.Tel.:+81272207930;fax:+81272207936.
E-mailaddress:[email protected](H.Hirai).
conserved, promotersfromratsand humansare oftenusedfor the generation of neuron-specific transgenic mice (Guo et al., 2002;Heumannetal.,2000;Hoesche etal.,1993;Holzeretal., 2001; Morimoto et al., 2002; Nakagawa et al., 2006; Sargeant etal.,2012;ScottandLois, 2005)orviralvector-mediatedgene transfer into rodent neurons (Dittgen et al., 2004; Hiokiet al., 2007;Kugleretal.,2003).Thefirsttransgenicmiceexpressinga transgeneundercontroloftheSynIpromotercarriedaconstruct containing∼4300nucleotidesofthe5-flankingsequenceoftherat synapsingene(Hoescheetal.,1993).ThesizeoftheSynIpromoter inthesetransgenicmicewasgraduallyshortenedinsubsequent 0165-0270/$–seefrontmatter© 2013 Elsevier B.V. All rights reserved.
http://dx.doi.org/10.1016/j.jneumeth.2013.12.004
studies(Heumannetal.,2000;Morimotoetal.,2002;Nakagawa etal.,2006;Sargeantetal.,2012;ScottandLois,2005).However, recentstudiesusingshorter versionsoftheSynIpromoter have failedtodemonstrate robusttransgeneexpression inthe hind- brain.Forexample,Nakagawaetal.(2006)generatedtransgenic miceexpressingglutamatereceptorinteractingprotein1(GRIP1), apostsynapticprotein,underthecontrolofa450-bphumanSynI promoter;however,thesemicedidnotshowtransgeneexpression inthecerebellum,althoughtransgeneexpressionwasobservedin thebrainstem,olfactorybulb,forebrainandhippocampus. More recently,transgenicmicecarryinga585-bphumanSynIpromoter showedminimalexpressionofthetransgeneinseveralregions, includingthebrainstemandspinalcord,incontrasttotheabundant expressionobservedinthecerebrumandhippocampus(Sargeant etal.,2012).Theseresultssuggestthatthedeletedregionmight berequiredforstabletransgeneexpressionthroughoutthecentral nervoussystem(CNS).
Amajordisadvantageof celltype-specificpromotersistheir weakertranscriptionalactivitywhencomparedwithconstitutive viralpromoterssuchasthecytomegalovirus(CMV)promoterand themurinestemcellvirus(MSCV)promoter(Boulosetal.,2006;
Nakagawaetal.,2006).Hiokietal.(2007)increasedtranscriptional activity2-to-3-foldafterfusingtheCMVenhancertothehuman SynIpromoter(401bp),asverifiedthroughthelentiviralexpres- sionofGFPintheratneostriatum,thalamusandneocortexinvivo.
However,theadditionoftheCMVenhancersignificantlydecreased theneuronalspecificityofneuron-specificpromoters,includingthe SynIpromoter(Hiokietal.,2007).Theseresultssuggestthataddi- tionoftheCMVenhancerisbeneficialforincreasingtranscriptional activitybutmightpotentiallycompromiseneuronalspecificity.
Spinocerebellarataxias(SCAs)areprogressiveneurodegenera- tivediseasesofmultipletypes.SCAsimpairawiderangeofCNS regions,includingthecerebralcortex,thalamus,midbrain,cere- bellum,brainstemandspinalcord(Seideletal.,2012).Neuronal nuclearinclusionsareahallmarkofmanytypesofSCAs,suchasSCA type1,type3andtype7(Seideletal.,2012),indicatingthatneurons aretheprimarycelltypesaffectedbytheseSCAtypes.Theselective deliveryofdisease-causinggenestosusceptibleregionsandcell typesiscriticalforthegenerationofanimalmodels.Althoughmany differenttypesofSCAmousemodelshavebeengeneticallyengi- neered,therearenoSCAmousemodelsshowingdiffuseexpression ofdisease-causinggenesexclusivelyinneuronsintheCNS(Ingram etal.,2012).AlthoughtheSynIpromoterisanidealcandidatefor thegenerationoftheseanimals,previousstudiesusingshortSynI promotershavefoundeithernotransgeneexpressionorscarce expressioninthecerebellumandbrainstem(Nakagawaetal.,2006;
Sargeantetal.,2012).Theaimofthepresentstudywastoobtain aSynIpromoterwithhighneuronalspecificityandrobusttrans- criptionalactivityextensivelyintheCNS.ThebehavioroftheSynI promoterinthecerebellumhasnotbeenextensivelystudied;thus, usinglentiviralvectors,weinitiallyexaminedthetransductionpro- fileofthreedifferentSynIpromotersofdifferentlengthscarrying CMV-derivedenhancersinthecerebellumsofmice.Asaresult,we generatedanSCAmousemodelusing1.0kboftheratSynIpro- moterfusedwiththeminimalCMVpromoter(minCMV)sequence.
2. Materialsandmethods 2.1. Animals
C57BL/6J(BL6)micewerepurchasedfromCLEA(Tokyo,Japan), andICRmicewereobtainedfromCharlesRiverLaboratories(Kana- gawa,Japan)andJapanSLC(Sizuoka,Japan).NineBL6miceand23 ICRmicewereusedinthisstudy.Allproceduresforthecareand treatmentofanimalswereperformedaccordingtotheJapanese
ActontheWelfareandManagementofAnimalsandtheGuidelines fortheProperConductofAnimalExperimentsissuedthroughthe ScienceCouncilofJapan.Theexperimentalprotocolwasapproved throughtheInstitutionalCommitteeofGunmaUniversity(no.09- 062).Alleffortsweremadetominimizesufferingandreducethe numberofanimalsused.
2.2. ConstructionofmodifiedSynIpromoters
For the construction of rSynI(1.0)-minCMV (Fig. 1A and B), 1.0kboftheratSynIpromoterregion(nucleotides1–1046;Gen- Bank:DD464830.1)wasamplifiedbypolymerasechainreaction (PCR)ofratgenomicDNA(1stPCRinSupplementaryFig.1A).The minCMVsequence(nucleotides1091–1218;GenBank:X03922.1) wasamplifiedbyPCRoftheCMVIE1promoter(1stPCRinSup- plementary Fig. 1A). The 5 end of the forward primer for the minCMVfragmentamplificationcontainedasequenceoverlapping the3 endoftheratSynIpromoter;thus,theratSynIpromoter and minCMV fragments were directly combined through over- lapping PCR (Overlapping 2nd PCR in Supplementary Fig. 1A).
rSynI(0.5)-minCMV(containingnucleotides511–1046;GenBank:
DD464830.1)wasobtainedfromrSynI(1.0)-minCMVbythe3rdPCR depictedinSupplementaryFig.1A.
CMVe-hSynI(0.4)wasconstructedaspreviouslyreported(Hioki et al., 2007). Briefly, using PCR, the CMV enhancer (CMVe, nucleotides1–364;GenBank:X03922.1)and0.4kbofthehuman synapsinIpromoterregion(hSynI,nucleotides1889–2289;Gen- Bank: M55301.1) were amplified from the CMV promoter and humangenome,respectively,andthetwoPCRproductswerelig- atedafterrestrictionenzyme(PstI)digestion(SupplementaryFig.
1B).Thesequencesofallprimersusedforgenerationofthethree hybridpromoterconstructsareshowninSupplementaryFig.1C.
Thegeneratedpromoters wereinsertedintotheMluI/EcoRI- digestedsiteofthepCL20c-GFPlentiviralvectorplasmidprovided bySt.JudeChildren’sResearchHospital(Memphis,TN,USA)(Fig.1A andB).
2.3. Constructionofalentiviralvectorplasmidexpressing ATXN1[Q98]
P2A, a ‘self-cleaving’ peptide sequence (ATNFS- LLKQAGDVEENPGP), was inserted between GFP and HA- ATXN1[Q98], which freed GFP from HA-ATXN1[Q98] by ribo- somalskippingbetweenglycineandproline(Donnellyetal.,2001;
Iizukaetal.,2009;Kuzmichetal.,2013).Thewoodchuckhepatitis virus posttranscriptional regulatory element (WPRE) sequence wasinserted following ATXN1[Q98]at theClaI-digested site of pCL20c(Donelloetal.,1998;Zuffereyetal.,1999).
Thehumanataxin-1 genewasamplifiedusing reversetran- scriptionPCR,andthehumaninfluenzahemagglutinin(HA)tag sequence was attached to the 5 end of the ataxin-1 gene. To enhancethepathology when expressedintransgenicmice,the numberofCAGrepeatsintheataxin-1genewasexpandedto98 repeats using two-step PCR (Laccone et al., 1999). Briefly, two ataxin-1fragmentsfromthe5 endtotheCAGrepeatregionand fromtheCAGrepeatregiontothe3endwereproducedthrough PCR using two pairs of primers: ATXN1-F (forward primer), 5-CGACGCGTGCCACCATGTACCCATACGAT-3/CTG7-R (reverse primer) 5-CTGCTGCTGCTGCTGCTGCTG-3 and CAG7-F (forward primer), 5-CAGCAGCAGCAGCAGCAGCAG-3/ATXN1-R (reverse primer), 5-CCATCGATCTACTTGCCTACATTAGACC-3. The 5- and 3-endfragmentsobtainedfromthe1stPCRreactionweremixed togetheraftergelpurification,anda2ndPCR reactionwasper- formedusingATXN1-F(aforwardprimer)andATXN1-R(areverse primer) toobtainthe full-lengthataxin-1 gene. The conditions usedforthe1stand2ndPCRreactionswere40cyclesof98◦Cfor
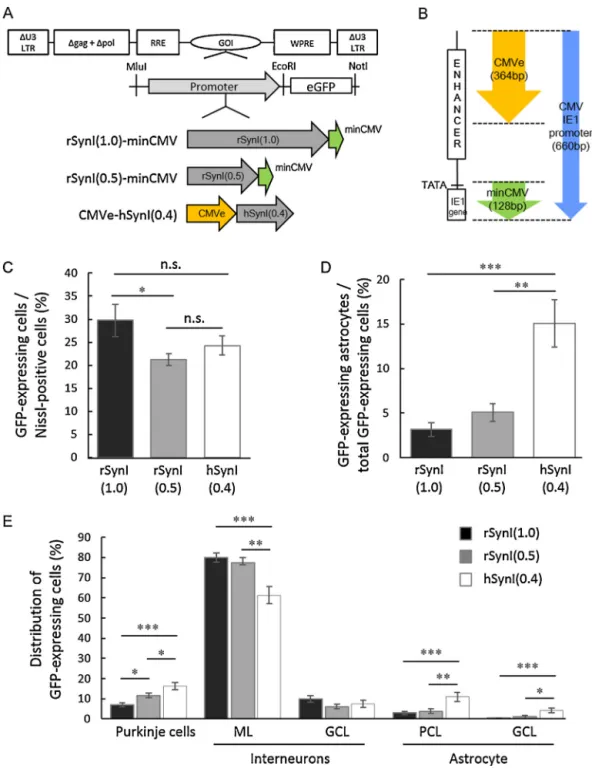
Fig.1.ComparisonofthreealteredhumanorratsynapsinI(SynI)promotersofdifferentsizes.(A)SchemadepictingtheproviralregionflankedbyU3-deletedlongterminal repeats(U3LTRs)andthecompositionofthreemodifiedSynIpromoters.EachofthemodifiedSynIpromotersandtheenhancedgreenfluorescentprotein(GFP)gene wereinsertedintolentiviralvectorsastheGOI(geneofinterest)andpositionedupstreamofthewoodchuckhepatitisvirusposttranscriptionalregulatoryelement(WPRE).
ThethreepromotersincludethehumanSynIpromoter(0.4kb)andtworatSynIpromoters(0.5kband1.0kb).ThehumanandratSynIpromoterswerefusedwiththe 364-bpcytomegalovirus(CMV)enhancer(CMVe,yellowarrow)atthe5end[CMVe-hSynI(0.4)]orwitha128-bpminimalCMV(minCMV,greenarrow)atthe3end [rSynI(1.0)-minCMVandrSynI(0.5)-minCMV],respectively.gag,deletedgagsequence;pol,deletedpolsequence;RRE,revresponseelement.(B)Adiagramshowingthe componentsoftheCMVIE1promoterandtherelativepositionsofCMVeandminCMVintheCMVIE1promoter(bluearrow).TheCMVIE1promoterincludestheenhancer region(ENHANCER),TATAboxandimmediateearly1(IE1)gene.(C)PercentageofneuronstransducedbylentiviralvectorscontainingoneofthemodifiedSynIpromoters.
Threemicewereusedfortheanalysisofeachpromoter(atotalofninemiceforthreepromoters).WecountedthenumberofGFP-expressingcellsinmorethan2000 Nissl-positiveneuronsinlobule4/5fromthreemice,andthepercentageratiosareexpressedinthegraph.(D)DecreaseintheneuronalspecificityofthreemodifiedSynI promoters.Threemicewereusedfortheanalysisofeachpromoter.WecountedthenumberofS-100-positiveastrocytesinmorethan500GFP-expressing-cellsinlobule 4/5,andthepercentageratioisexpressedinthegraph.(E)CelltypestransducedbythemodifiedratorhumanSynIpromoters.Threemicereceivedaninjectionofalentiviral vectorcontainingoneofthemodifiedSynIpromoters(atotalofninemiceforthreepromoters).CerebellarsectionswerelabeledforS-100andNissl.ThenumberofPurkinje cells,interneuronsandastrocytesinmorethan500GFP-positivecellsinlobule4/5fromthreemicewerecounted,andtheirratiosweredetermined.Purkinjecellswere distinguishedfrominterneuronsbytheiruniquemorphologyanddiameterofthesoma.ML,molecularlayer;GCL,granulecelllayer;PCL,Purkinjecelllayer.Thedataare themeans±S.E.M.Statisticalanalysiswasperformedbyone-wayANOVAwithTukey’sposthoctest:*p<0.05,**p<0.01,***p<0.001.n.s.,notsignificant.(Forinterpretation ofthereferencestocolorinthisfigurelegend,thereaderisreferredtothewebversionofthisarticle.)
10s,60◦Cfor 5sand 72◦C for90s. AfterexpansionoftheCAG tract,theataxin-1genewasinsertedintotheNotIrestrictionsite ofthepCL20c-rSynI(1.0)-minCMV-GFP-P2A-(NotI)-WPREplasmid usingKOD-Plus-NEOpolymerase(Toyobo,Osaka,Japan)andthe InFusionHDcloningkit(TakaraBio,Shiga,Japan)(Fig.4A).
2.4. Productionandtitrationoflentiviralvectors
The vesicular stomatitis virus-glycoprotein (VSV-G)- pseudotyped lentiviral vector was produced in HEK293T cells (Thermo Fisher Scientific, Kanagawa, Japan), as previously described(Torashimaetal.,2008).Theplasmidsweretransfected intoHEK293Tcellsusingthecalciumphosphatemethod.After48h, thesupernatant,containingviralparticles,washarvested,ultra- centrifuged(CP80WX;HitachiKoki,Tokyo,Japan)andresuspended in70lofDulbecco’sphosphate-bufferedsaline(−).Theresultant lentiviralsolutionwasstoredat4◦Candusedwithin3weeks.The viraltitersweremeasuredusingthequantitativereal-timeRT-PCR method.We usedanRNeasyMiniKit (Qiagen,Tokyo,Japan)to preparegenomicRNAfrom1loftheviralsolution.Reversetran- scriptionwasperformedusingtheReverTraAceqPCRRTMaster MixwithgDNARemover(Toyobo).Forviraltitration,quantitative real-timePCRwasperformedusingtheTakarathermalcyclerDice TP800system(TakaraBio)andSYBRPremixExTaqII(TakaraBio) withcyclesat95◦Cfor5sand60◦Cfor30s.Thefollowingprimer pairswere used:EGFP-F, 5-CGACCACTACCAGCAGAACAC-3 and EGFP-R,5-TGTGATCGCGCTTCTCGTTGG-3.
2.5. Generationoftransgenicmice
Transgenicmiceweregeneratedafterlentiviralinjectioninto the perivitelline space of 2-cell mouse embryos, as previously described(Oueet al.,2012).The2-cell embryoscollectedfrom superovulatedICRfemalemiceweretreatedwithserotropin(5IU;
ASKA Pharmaceutical Co., Tokyo, Japan) and gonatropin (5IU;
ASKAPharmaceuticalCo.).Thelentiviralmicroinjectionwasper- formedusingaFemtoJetmicroinjector(EppendorfAG,Hamburg, Germany)equippedwithaFemtoTip(EppendorfAG)andathree- axishangingjoystickoilhydraulicmicromanipulator(Narishige, Tokyo,Japan). Theinjectedembryosweretransplantedintothe oviductsofpseudo-pregnantfemales.Fluorescent imagesofthe brainandspinalcordofnewbornpupswereobtainedusingaCCD camera(VB-7010;Keyence,Osaka,Japan) attachedtoafluores- cencestereoscopicmicroscope(VB-L10;Keyence).Thegenotyping wasperformed by PCR amplification of genomic DNA samples fromthetoetips of weanedpupsusing thefollowing primers:
minCMV-F, 5-TTTTGACCTCCATAGAAGACACC-3 and EGFP-R,5- TGGTGCAGATGAACTTCAGGG-3.
2.6. Histologicalanalysis
The mice were deeply anesthetized with a combination of ketamine(150mg/kgbodyweight)andxylazine(15mg/kgbody weight)andtranscardiallyperfusedwithPBSandafixativecon- taining4%paraformaldehydein0.1Mphosphatebuffer.Thebrains werecutinto50msagittalsectionsusing amicroslicer(DTK- 1000; Dosaka, Kyoto, Japan). Images of GFP fluorescence were obtainedusingafluorescencemicroscope(BZ-9000;Keyence)or aconfocallaser-scanningmicroscope(LSM5PASCAL;CarlZeiss, Oberkochen,Germany).
2.7. Immunohistochemistry
Floatingbrainsectionswereimmunostainedinblockingsolu- tion (2%bovine serum albumin,2% normal donkey serum and 0.4%TritonX-100inphosphatebuffer)for1dayat4◦Cwiththe
followingprimaryantibodies: ratmonoclonalanti-GFP(1:1000;
04404-84; Nacalai Tesque, Kyoto, Japan), rabbit monoclonal anti-GFP(1:1000;GFP-Rb-Af2020-1;FrontierInstitute,Hokkaido, Japan), mouse monoclonal anti-S100 (1:1000; S2532; Sigma- Aldrich, St. Louis, MO, USA), rat monoclonal anti-HA (1:1000;
1867423;RocheDiagnostics,Basel,Switzerland)andrabbitpoly- clonal anti-calbindin D-28K (1:500; AB1778; Merck Millipore, Billerica,MA).Theboundprimaryantibodieswerevisualizedafter incubationwithAlexaFluor488donkeyanti-ratIgG(1:1000;Life Technologies,Gaithersburg,MD,USA),AlexaFluor568donkeyanti- mouseIgG(1:1000;LifeTechnologies)andAlexaFluor568donkey anti-ratIgG(1:1000;LifeTechnologies)inblockingsolutionfor2h atroomtemperature.Afterthesecondaryantibodyreaction,Nissl bodieswerestainedusingNeuroTrace530/615(1:200;LifeTech- nologies)orNeuroTrace640/660(1:200;LifeTechnologies)inPBS for40minatroomtemperature.
2.8. Measurementofthethicknessofthemolecularlayerinthe cerebellarcortex
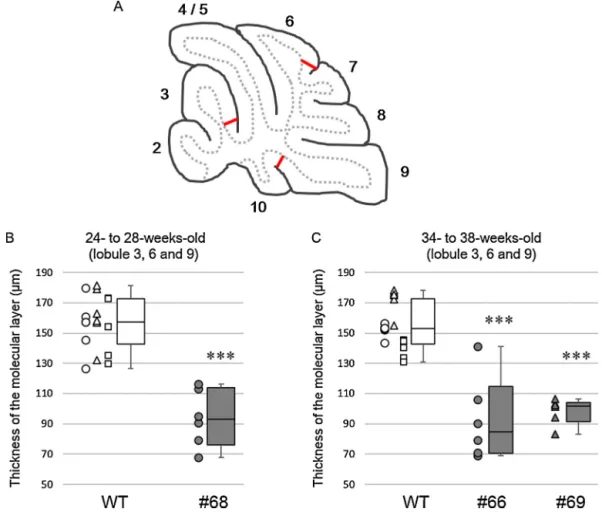
Thethicknesses of themolecularlayers from3 SCA1 model founders(no. 66, no.68 and no.69)were measuredand com- paredwiththatofwild-typemice.BecausetheSCA1modelfounder no.68wassacrificed at 26weeks ofage,three wild-typemice at26±2weeksofagewereusedforcomparison,while34–38- week-old wild-type mice were used as controls for the SCA1 modelfoundersno.66 and no.69, which weresacrificedat 38 weeksofage.Sagittalsections(50mthickness)wereprepared fromthecerebellarvermis(within0.4mmfromthemidline)and immunolabeledforcalbindin-D28K.Immunofluorescenceimages of the cerebellar cortex were obtained with a confocal laser- scanningmicroscope(LSM5PASCAL,CarlZeiss).Thethicknessof themolecularlayerwasmeasuredatlobule3,lobule6andlobule 9usingtwocerebellarsectionsfromonemouse.Theexactloca- tionsatwhichthethicknesswasmeasuredaredepictedinFig.7A.
Themolecularlayerwasdefinedfromtheoriginoftheprimary dendrites of Purkinjecells tothedistal end just below the pia mater.
3. Results
3.1. Assessmentofthepromoterstrengthandneuronalspecificity ofthethreeSynIpromoters
WepreparedthreeSynIpromotersofdifferentlengthsfused withCMV-derivedenhancers(Fig.1A).Onepromoter,comprising the401-bphumanSynIpromoterwiththeupstream364-bpCMVe fromtheCMVE1promoter[CMVe-hSynI(0.4)](Fig.1B),hasbeen previouslyreportedasbeingsuperiortootherneuron-specificpro- motersintermsofneuronalspecificityandtranscriptionalactivity (Hiokietal.,2007).Theothertwopromoterscomprised0.5or1.0- kbratSynIpromoterswithadownstreamminCMVregionfrom theCMVE1promoter[rSynI(0.5)-minCMVorrSynI(1.0)-minCMV, respectively](Fig.1AandB).Eachofthepromoters,togetherwith theenhancedgreenfluorescentprotein(GFP)gene,wassubcloned intothelentiviralplasmid(Fig.1A).TheWPREwasplaceddown- streamoftheGFPgenetoenhancegeneexpression(Donelloetal., 1998;Zuffereyetal.,1999).
Thepromoterstrengthandneuronalspecificityofthemodified SynI promoters were examined through the lentiviral vector- mediatedexpressionofGFPinthecerebellarcortex.Afteradjusting thegenometitersofthelentiviralvectors,10loflentiviralsolu- tionwasinjectedintothesubarachnoidalspaceoverlobules4/5of thecerebellaof3C57BL/6miceatpostnatalday(P)21–24.The micewereexaminedat7 daysafterviralinjection. Fluorescent
stereoscopicexaminationshowedbrightfluorescenceinthecere- bella treated with lentiviral vectors expressing GFP under the controlofthemodifiedSynIpromoter(SupplementaryFig.2A–C).
Toassessthetransductionareaandcelltypesinthecerebellum, sagittalsliceswereimmunostainedforNisslsubstance,aneuronal marker,orS-100,anastrocytemarker.Low-magnificationimages ofthe sagittalsectionsofthe cerebellumshoweda fairly simi- larrange ofGFP expression,whichwascentered atlobules4/5 andextendedtothecortexoflobule9throughthewhitematter (SupplementaryFig.2D–F).TheseresultssuggestthatrSynI(0.5)- minCMV and rSynI(1.0)-minCMV exhibit strong transcriptional activitysimilartoCMVe-hSynI(0.4),whichwaspreviouslyreported tohavestrongpromoteractivity(Hiokietal.,2007).Theenlarged imagesaroundlobules4/5showedthatallthreepromotersinduced GFPexpressionprimarilyinneurons,whichweremorphologically distinguishedasPurkinjecellsandinterneuronssuchasstellate, basketandGolgicells(SupplementaryFig.2G–I).Asreportedpre- viously (Torashima et al., 2006), the lentiviral vectors did not transducegranulecells.Toquantitatetheefficiencyoflentiviral transductionofneurons,wecountedneuronsthatweredouble- positiveforGFPandNisslinthecortexoflobule4/5anddivided thisnumberbythenumberofNissl-positivecells,omittinggran- ulecellsbecausenogranulecells weretransducedbylentiviral vectors.rSynI(1.0)-minCMVcausedasignificantlyhigherneuronal transductionfrequency (29.7%)than rSynI(0.5)-minCMV(21.3%) (Fig.1C).
SomeGFP-expressingcellswereimmunolabeledfor S-100,a glialmarker,whichrevealedthatthemodifiedSynIpromoterswere not 100% specificfor neurons(Supplementary Fig.2J–L, arrow- heads).Toassesstheneuronalspecificityofthethreepromoters, wedeterminedtheratioofastrocytesthatweredouble-positivefor GFPandS-100tothetotalnumberofGFP-positivecellsinlobule4/5 (Fig.1C).CMVe-hSynI(0.4)showedsignificantlygreaterexpression inastrocytes(15.1%)thandid1.0kband0.5kbratsynapsinI-based promoters(3.1%and5.1%,respectively)(Fig.1D).
Wenextexaminedthecorticalcelltypestransducedbythethree promoters.Weanalyzedmorethan500GFP-expressingcellsinthe cerebellarcorticesfromthreemicethatweretreatedwithlentiviral vectorscontainingone ofthemodified SynIpromoters,and the percentratioofPurkinjecells,interneuronsandastrocytestototal GFP-expressingcellswascalculated(Fig.1E).Consistentwiththe resultsshowninFig.2D,theastrocyteratiowassignificantlyhigher incellstransducedwithCMVe-hSynI(0.4)(15.1%)comparedwith thosetransducedwiththetwoothermodifiedratSynIpromoters (3–5%).CMVe-hSynIwasmorelikelytotransducePurkinjecells (16.1%)thanthemodifiedratSynIpromoters(7.0–11.7%);however, themodifiedratSynIpromoterstransducedinterneuronsinthe molecularlayer(namely,stellatecellsandbasketcells)significantly moreefficientlythanCMVe-hSynI.
We further examinedthe transduction profiles of the three promotersonthebasisofGFPintensityinPurkinjecells,interneu- rons and astrocytes, which was measured in the somata of
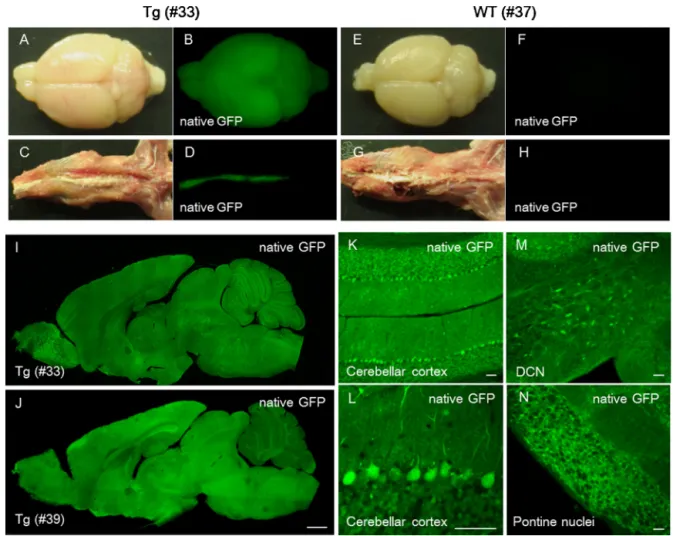
Fig.2. Lentiviral-generatedtransgenicmiceexpressingGFPunderthecontrolofrSynI(1.0)-minCMVintheCNS.(A–H)NativeGFPfluorescenceandbrightfield(BF)images ofthebrainandspinalcordfromaP35transgenicmouse(no.33)(A–D)andanage-matchedwild-typelittermate(E–H).(IandJ)GFPfluorescenceimagesofsagittalsections ofwholebrainsfromatransgenicmouse(no.33,P35)andtransgeniclittermate(no.39,P378).(K–N)NativeGFPfluorescenceimagesofthecerebellarcortex(KandL),deep cerebellarnuclei(DCN)(M)andpontinenuclei(N)fromatransgenicmouse(no.39).Anenlargedimageofthecerebellarcortex(L)showstheefficientexpressionofGFPin Purkinjeandgranulecells.Scalebars=1mm(J)and50m(K–N).
GFP-expressingcellsusingImageJ.GFPintensityinPurkinjecells transduced by lentiviral vectors containing rSynI(1.0)-minCMV wassignificantlylowerthanthatincellstransducedwiththeother twopromoters(SupplementaryFig.3).Ontheotherhand,theGFP intensityininterneuronsofthegranulecelllayertransducedby lentiviralvectorscontainingCMVe-hSynI(0.4)wasapproximately halfthatobservedin cellstransducedby lentiviralvectorscon- taining themodified rat SynI promoters. Taken together,these dataindicatethatrSynI-minCMVissuperiortoCMVe-hSynI(0.4)in termsofneuronalspecificity,andrSynI(1.0)-minCMVsignificantly moreefficientlytransducedcerebellarcorticalneuronscompared withrSynI(0.5)-minCMV.Thus,itseemedthatrSynI(1.0)-minCMV wassuperiortotheothertwopromotersforefficientandspecific neuronaltransduction.
3.2. GenerationoftransgenicmicethatexpressGFPunderthe controlofrSynI(1.0)-minCMV
ToexaminethetransgeneexpressionprofileoftherSynI(1.0)- minCMV in the CNS, transgenic mice expressing GFP under thecontrolof therSynI(1.0)-minCMVpromoter were produced throughthelentiviralvector-mediatedtransferofthetransgene intothechromosomesoffertilizedeggs.Intotal,8ofthe13pups generatedpossessedthetransgene.Thebrain(SupplementaryFig.
4AandB,arrows)andspinalcord(SupplementaryFig.4AandB, arrowheads)of thelivetransgenicpupsemittedbrightfluores- cencethatwasvisiblethroughthebonesandskinunderfluorescent stereoscopicmicroscopy.Thetransgenicmice,togetherwiththeir wild-typelittermates,were sacrificedat P35 (no.33) andP378 (no.39).Examinationoftransgenicmouseno.33revealedrobust GFPexpressionthroughouttheCNSunderfluorescentstereotaxic microscopy (Fig. 2A–D).The observed signal wasnot autofluo- rescencebecausealmost nofluorescentsignal wasobserved in thebrain and spinal cordof an age-matched wild-type mouse (Fig.2E–H).Thesagittalsectionsofthebrainfromtwotransgenic mice(Fig.2IandJ) showedbrightGFPfluorescencethroughout thebrains,evenat1yearafterbirth(no.39,Fig.2J).Althoughthe expressionofGFPinthecerebellumwaslowerthanthatinthe otherbrainregionsintransgenicmouseno.39(Fig.2J),theenlarged imagesofthecerebellumandassociatedbrainstemregionsshowed abundantGFPexpressioninPurkinjeandgranulecellsaswellasin neuronsinthedeepcerebellarandpontinenuclei(Fig.2K–N).
Toconfirmneuron-specificGFPexpression,brainsectionswere immunostainedforNisslsubstance.GFPwasexpressedextensively inmostneuronsinthecerebellumandassociatedbrainstemnuclei, suchaspontine,vestibularandinferiorolivarynuclei(Fig.3).Sim- ilarlevelsofGFPweredetectedinneuronselsewhereinthebrain, includingthecerebralcortex,hippocampus,thalamusand basal ganglia(datanotshown).Quantitativeanalysisofthetransgenic mice(no.33andno.39)revealedthatmorethan99.5%ofGFP- expressingcellswereneurons(Table1).
3.3. GenerationofSCA1modelmiceexpressingmutantataxin-1 andGFPviarSynI(1.0)-minCMV
WenextexaminedwhetherrSynI(1.0)-minCMVcouldbeused forthegenerationofamousemodelofdiseasethataffectsneurons diffuselyintheCNS.Inthisstudy,weselectedSCAtype1(SCA1), whichimpairsthecerebralcortex,basal ganglia,thalamus,hip- pocampus,brainstemandcerebellum(Seideletal.,2012).SCA1was inducedthroughtheabnormalexpansionofCAGrepeatsinthesca1 gene(Orretal.,1993),leadingtotheproductionofataxin-1pro- tein(ATXN1)withanabnormallyexpandedpolyglutaminetract.
Mutant ATXN1forms nuclearaggregates with nuclearproteins thatare criticalfor neuronalfunction, resultinginneurodegen- eration and cell death. Using lentiviral vectors, we generated
Fig.3.Neuron-specifictransgeneexpressionintransgenicmiceexpressingGFP underthe controlofrSynI(1.0)-minCMV.Sagittalbrain sectionswere double- immunolabeledforNisslsubstance(red)andGFP(green).Theimagesontheright aremagnificationsoftheimagesshownontheleft.(AandB)Cerebellarcortex;(C andD)deepcerebellarnuclei(DCN);(EandF)pontinenuclei;(GandH)vestibular nucleus;(IandJ)Inferiorolivarynucleus.Scalebar=20m.(Forinterpretationof thereferencestocolorinthisfigurelegend,thereaderisreferredtothewebversion ofthisarticle.)
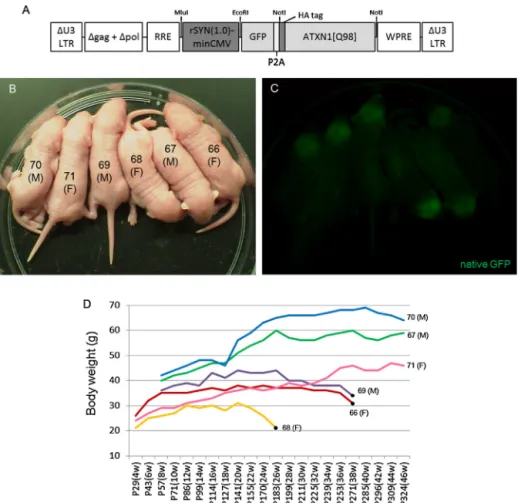
transgenicmiceexpressingGFPandATXN1containingastretch of98polyglutamineresidues(ATXN1[Q98])underthecontrolof rSynI(1.0)-minCMV(Fig.4A).
The17embryosthatreceivedlentiviralinjectionswerereturned toa pseudo-pregnantfemalemouse.Sixpups(no.66–71)were obtained,allofwhichweregenotypepositive.Thebrainsandspinal cordsof theP3 pups emittedbright fluorescencethat wasvis- iblethrough thebonesand skinunder fluorescentstereoscopic microscopy(Fig.4Band C). TheSCA1 model micegained body weightsteadilywithincreasinggrowth.However,no.68started
Table1
Neuronalspecificityofthetransgenicmicegeneratedinthecurrentstudythat expresstransgenesviatherSynI(1.0)-minCMVpromoter.Nisslwaslabeledinneu- ronswithNeuroTrace.ThepercentratiosofneuronsamongtheGFP-expressing cellsweredeterminedinthevariousbrainregions.Ineachregion,200–450GFP- expressingcellswereexamined.DCN,deepcerebellarnuclei.
GFP GFP-P2A-ATXN1(Q98)
No.33 No.39 No.66 No.68 No.69
Cerebellarcortex 99.5 100 98.8 100 99.3
DCN 100 100 100 100 100
Pontinenuclei 99.6 99.6 100 100 99.7
Hippocumpus 100 100 100 100 100
Cerebralcortex 99.7 99.6 99.7 100 100
Total 99.8 99.9 99.7 100 99.8
PercentratioofGFP-andNissl-double-positive-cellsingeneratedtransgenicmice.
toloseweightat20weeksofage(Fig.4D).Theothertwomice (no.66and no.69)alsostartedtoloseweightgraduallybegin- ningat27 weeks.Consistently,thethree miceshowedclasping (SupplementaryFig.5A)andataxicbehavior(SupplementaryFig.
5BandSupplementarymovie).Ataxiabecamemoreseverewith time;thus,thethreemicewereeuthanizedat26(no.68)and38 (no.66andno.69)weeksofage.Transgenicmouseno.71died spontaneouslyat49weeksofage.
ThewholebrainsandspinalcordsfromtheSCA1modelmice (no.68,no.66andno.69)emittedbrightGFPfluorescenceunder
fluorescentstereoscopicmicroscopy(Fig.5AandBandSupplemen- taryFig.6A–D).SagittalsectionsshoweddiffuseGFPexpression throughoutthebrain(Fig.5CandSupplementaryFig.6EandF).
Enlargedimagesofasagittalsectionfrommouseno.68showed efficientGFPexpressioninPurkinjeandgranulecellsinthecere- bellarcortex(Fig.5D)aswellascellsinthedeepcerebellar,pontine (datanotshown)andinferiorolivarynuclei(Fig.5E).Thedisrupted arrangementofPurkinjecellsomatawasclearlyvisible(Fig.5D).
Wenextexaminedneuronalspecificityinthetransgenicmice bymeasuringtheratioofneurons(cellsdouble-positiveforGFP andNissl)intransducedcells(GFP-positivecells)inthecerebellar cortex,deepcerebellarnuclei,pontinenuclei,CA3ofthehippocam- pusandcerebralcortex.Wecounted200–450GFP-positivecellsin eachregionfromeachmouse.Theresultshowedthattheneuronal specificitywasgreaterthan99.3%inallregionsofthetransgenic miceexamined(Table1).
TovisualizetheexpressionandlocalizationofATXN1[Q98],we immunostainedbrainsectionsfromtheSCA1modelmiceanda control wild-typemouse (28 weeksold) forHA, which was N- terminallyfusedwithATXN1[Q98],andNisslsubstance(Fig.6).The miceusedforimmunohistochemistrywererelativelyolder(26- and38-week-old)mice;therefore,non-specificgranularimmuno- labelingoflipofuscinwasobservedinthecytoplasmbutnotinthe nuclei(BrunkandTerman,2002;Nandy,1971).Inadditiontothe non-specificimmunolabeling,all3SCA1mousemodels(no.68,no.
66andno.69)showedATXN1[Q98]aggregateslocalizeddiffusely inthenucleiofneuronsinthecerebellarcortex,deepcerebellar
Fig.4. Lentiviral-generatedtransgenicmiceexpressingGFPandATXN1withanabnormallyexpandedpolyglutaminestretch.(A)Adiagramdepictingtheprovirusflanked byU3LTRs.TranscriptionofGFPandanataxin-1cDNAcontaining98CAGrepeats(ATXN1[Q98])drivenbyrSynI(1.0)-minCMV.Ahumaninfluenzahemagglutinin(HA)tag wasattachedtothe5endofATXN1[Q98].Aself-cleaving2Asequence(P2A)wasinsertedbetweenGFPandmutantataxin-1cDNAtoexpressATXN1[Q98]freedfromGFP.
(BandC)Brightfield(B)andGFPfluorescent(C)stereoscopicimagesofthetransgenicpups.Thenumberandsex(maleorfemale)ofeachmouseisindicatedonthebody.
(D)Agraphshowingchangesinthebodyweightsoftransgenicmice.Becausetransgenicmiceno.68,no.69andno.66developedsevereataxiawithdecreasedbodyweight, theanimalsweresacrificedtoexaminethepathologicalchangesinthebrainatthepointsindicatedwithblackdots.
Fig.5.NativeGFPinthebrainofatransgenicmouse(no.68,P185)expressingGFPandATXN1[Q98].Thetransgenicmouse(no.68)didnotgainweightuntil12weeksafter birthandbegantoloseweightat20weeksafterbirth.Inparallel,aprogressiontowardsevereataxiawasnotedamongthelittermates.ThemousewassacrificedatP185 forpathologicalexamination.(AandB)Brightfield(A)andGFPfluorescent(B)stereoscopicimagesofthewholebrainofthetransgenicmouse.(C)Asagittalsectionofthe brain.NotethatGFPwasexpressedthroughoutthetransgenicbrain,similartothepatternobservedintransgenicmiceexpressingGFPalone.(DandE)Enlargedimagesof theregionsprimarilyaffectedbySCA1,includingthecerebellarcortex(D)andtheinferiorolivarynucleus(E).Scalebars=1mm(C),20m(D)and50m(E).
nuclei,pontinenuclei,hippocampusandcerebralcortex(Fig.6).
Theseaggregateswerenotobservedinthecontrolwild-typemice (Fig.6P–T).Inthecerebellarcortex,strongimmunoreactivitywas observedinPurkinjecellnuclei(Fig.6A,FandK).Thethickness ofthemolecularlayerwassignificantlydecreasedinSCA1model micecomparedwiththecontrolwild-typemouse(arrowsindicate thetopandbottomofthelayer,Fig.6A,F,KandP)(Fig.7).Inthe deepcerebellarnuclei,immunoreactivitywasprimarilyobserved inlargeglutamatergicneurons.Thus,althoughtheexpressionof thetransgenedifferedamongthetransgenicindividuals,thetrans- genewasstably expressedthroughoutthebrain, indicatingthe reliabilityandstabilityofrSynI(1.0)-minCMVfordiffusetransgene expressioninneuronsoftheCNS.
4. Discussion
Inthisstudy,wegeneratedamousemodelofaneurodegen- erativedisease,SCA1,usinglentiviralvectorscontaininga1.0-kb ratSynIpromoterfusedwithminCMVatthe3 end.Transgenic miceexpressedmutantataxin-1specificallyinneuronsthrough- outtheCNS.Adetailedexaminationusingimmunohistochemistry revealednuclear aggregates of mutant ataxin-1 in the primary regionsaffectedbySCA1,includingthecerebralcortex,cerebel- lar cortex, deep cerebellar nuclei and brainstem (Seidel et al., 2012).
Variousneuron-specifictransgenicmicehavebeenproduced forstudyingmoleculesofunknownfunctionandforexaminingthe pathologyofanddevelopingnoveltherapiesforneuronaldiseases (Boy et al.,2010; Burrightetal., 1995;Gendron and Petrucelli, 2011;Nuberetal.,2008;Torashimaetal.,2008;Wenetal.,2002).
Although transgenic mice expressingtransgenes driven by the SynI promoter have been generated (Hoesche et al., 1993), its weaktranscriptionalactivity,whichisacommonfeatureofcell
type-specific promoters, often yields ambiguous outcomes. To avoidambiguity,atetracycline-controlledtransactivationsystem (Gossen and Bujard,1992;Odeh et al.,2011)hasbeenusedto compensate for the insufficient transcriptional activity of the SynI promoter (Chen et al., 2013; Kuhn et al., 2012; Sargeant etal.,2012).However,aseveralstudieshavedemonstratedthe toxicityofthetetracyclinetransactivator(McCloskeyetal.,2005;
MorimotoandKopan,2009;Sissonetal.,2006),whichsometimes influences the phenotypes of the resultant transgenic rodents, includingtheirbehavior (Barton etal., 2002; Han etal., 2012).
Using lentiviral vector-mediated transgene expression in the mousebraininvivo,arecentstudydemonstratedthattheaddition of the CMV enhancer to the SynI promoter strengthened pro- moteractivity(Hiokietal.,2007).Thus,theviralvector-mediated expressionsystemis apowerfulapproach forscreeningvarious promoterconstructs. However,thetransgeneexpressionprofile ofapromoterintransgenicanimalsdoesnotalwayscorrespond tothatobservedinviralvector-mediatedexpression(Oueetal., 2012),aspromoter constructsinsertedintothechromosomeof transgenicmicemightundergovariousdevelopmentalmodifica- tions,suchasDNAmethylationandacetylation,therebysilencing or enhancing the promoter activity depending on the specific areaandcelltype.Therefore,afterscreeningapromoterconstruct throughviralvector-mediatedexpressioninthebraininvivo,the transgeneexpressionprofileof thepromoter shouldbe further examinedintransgenicmice.
Themost commonly acceptedmethodfor the generationof transgenicmiceispronuclearinjectionoflinearizedDNAintofer- tilizedoocytes(Gordonetal.,1980).Amajordisadvantageofthis methodistherelativelylowefficiency,particularlyinspeciesother than mice(Hirabayashi etal., 2001; Park, 2007).Althoughdis- easemodelsof non-humanprimatesare crucial fortheclinical translation of resultsfrom basicresearch,there is a significant
Fig.6.ATXN1[Q98]aggregatesinneuronsofvariousregionsprimarilyaffectedbySCA1.SagittalsectionsfromthetransgenicmiceexpressingGFPandATXN1[Q98](no.66, no.68andno.69)andthecontrolwild-typemouseweredouble-immunostainedforNisslsubstance(blue)andHA(magenta)fusedtotheamino-terminusofATXN1[Q98].
Diffusecytoplasmicgranularspotsimmunolabeledwiththeanti-HAantibodyshowednon-specificimmunolabelingforlipofuscin,whichaccumulatesinpostmitoticcells withincreasingage.(A,F,KandP)Cerebellarcortex;(B,G,LandQ)deepcerebellarnuclei(DCN);(C,H,MandR)pontinenuclei;(D,I,NandS)hippocampus;(E,J,Oand T)cerebralcortex.ATXN1[Q98]waslocalizedspecificallyintheneuronsinthebrainareasexamined.Notethatthethicknessofthemolecularlayer,indicatedwiththe verticalarrows,ismarkedlythinnerintransgenicmiceexpressingATXN1[Q98]andGFP(A,FandK)comparedwiththecontrolwild-typemouse(P).Scalebar=20m.(For interpretationofthereferencestocolorinthisfigurelegend,thereaderisreferredtothewebversionofthisarticle.)
obstacletothegenerationoftransgenicnon-humanprimatesusing conventionalmethods.Thelentiviralvector-mediatedinsertionof atransgeneintothechromosomeisanotherapproachforthegen- erationoftransgenicanimals(Oueetal.,2012;Park,2007).Indeed, transgenicmarmosetsexpressingGFPunderthecontrolof con- stitutiveviralpromotershavepreviouslybeendeveloped(Sasaki etal.,2009).Lentiviraltransgenesishasseveraladvantagesover conventionalpronuclearinjection,whichincludestheproduction oftransgenicanimalswithhigherefficiency,particularlyinspecies otherthanmice(Park, 2007).Moreover,thelow transcriptional activityofcelltype-specificpromoterscanbeboostedthroughthe additionoftheWPREdownstreamofthegeneofinterest.Forthe lentiviraltransferofatransgene,thesizeofthetransgeneflanked
bythelongterminalrepeatsshouldbelessthan8kbbecauseof packaginglimitations;ifthetransgenesizeis withintheselim- its,itcanbeefficientlyexpressed.Inthisstudy,weuseda1.0-kb ratSynIpromoter,whichshowedincreasedtranscriptionalactivity withoutcompromisedneuronalspecificitywhena128-bpminCMV sequence was fused to its 3 end. Approximately 14 of the 19 animalsbornduring thisstudypossessed transgenes,and most oftheanimalsrobustlyexpressedthetransgenethroughoutthe CNS.Thediscrepancybetweenourresultsandthoseofprevious reportsshowingabsentorfainttransgeneexpressioninthecere- bellumorbrainstem(Nakagawaetal.,2006;Sargeantetal.,2012) mightreflecttheuseofalongerSynIpromoter.Alternatively,the appendedminCMVmighthavestrengthened theSynIpromoter
Fig.7.DecreaseinthethicknessofthemolecularlayersinSCA1modelmice.(A)Schemadepictingthelocationsatwhichthethicknessofthemolecularlayerwasmeasured.
(BandC)MolecularlayerthicknessofSCA1modelmiceno.68(26weeksold),no.66andno.69(both38weeksold)andage-matchedwild-typemice(WT).Themolecular layerthicknesswasmeasuredatthreelobulesusingtwocerebellarslicespermouse.Circles,trianglesandsquaresarevaluesobtainedfromdifferentanimals.Foreachbox, theboxboundariesindicatethefirstandthirdquartiles,themiddlelinesintheboxesindicatethemedianandthewhiskersrepresentthelowestandhighestvalues.Asterisks abovethewhiskersindicatesignificantdifferencesbetweenthewild-typeandSCA1modelmiceasdeterminedbytheWilcoxon–Mann–Whitneytest(***p<0.001).
activitysufficientlysuchthatthetransgenewasalsoexpressedin thehindbrain.
The transgenic mice expressing mutant ataxin-1 displayed ataxicbehavior.Immunohistochemistryrevealedmutantprotein aggregates consistently in the nuclei of neurons in the major brainregionsaffectedbySCA1.Thus,the1.0-kbratSynIpromoter attachedtominCMVexhibitedgreatpotentialforthestableand robustexpressionofatransgenethroughouttheCNS.Becausethe SynIpromoter ishighlyhomologousacrossspecies,atransgene expressionprofile similarto that observed in this study might beobtainedusingthesameSynIpromoterindifferentanimals, includingnon-humanprimates.Therefore,lentiviraltransgenesis usingthismodifiedSynIpromotermaybeemployedinthegen- erationofvariousanimalmodels,includingnon-humanprimates, fordiseases thataffect theCNS,suchasseveralmajortypes of SCA.
Conflictofintereststatement
Theseauthorsdeclarenoactualorpotentialconflictsofinterest concerningtheworkpresentedinthismanuscript.
Acknowledgments
Thisworkwas supportedbya grant fromtheFunding Pro- gramforNextGenerationWorld-LeadingResearchers(LS021)(to H.Hirai).
AppendixA. Supplementarydata
Supplementarydataassociatedwiththisarticlecanbefound, in the online version, at http://dx.doi.org/10.1016/j.jneumeth.
2013.12.004.
References
BartonMD,DunlopJW,PsaltisG,KulikJ,DeGennaroL,KwakSP.ModifiedGFAP promoterauto-regulatestet-activatorexpressionforincreasedtransactivation andreducedtTA-associatedtoxicity.BrainResearchMolecularBrainResearch 2002;101:71–81.
BoulosS,MeloniBP,ArthurPG,BojarskiC,KnuckeyNW.AssessmentofCMV,RSVand SYN1promotersandthewoodchuckpost-transcriptionalregulatoryelementin adenovirusvectorsfortransgeneexpressionincorticalneuronalcultures.Brain Research2006;1102:27–38.
BoyJ,SchmidtT,SchumannU,GrasshoffU,UnserS,HolzmannC,SchmittI,Karl T,LacconeF,WolburgH,IbrahimS,RiessO.Atransgenicmousemodelof spinocerebellarataxiatype3resemblinglatediseaseonsetandgender-specific instabilityofCAGrepeats.NeurobiologyofDisease2010;37:284–93.
BrunkUT,TermanA.Lipofuscin:mechanismsofage-relatedaccumulationandinflu- enceoncellfunction.FreeRadicalBiologyandMedicine2002;33:611–9.
BurrightEN,ClarkHB,ServadioA,MatillaT,FeddersenRM,YunisWS,DuvickLA, ZoghbiHY,OrrHT.SCA1transgenicmice:amodelforneurodegenerationcaused byanexpandedCAGtrinucleotiderepeat.Cell1995;82:937–48.
ChenY,CaoL,LuoC,DitzelDA,PeterJ,SprengelR.Genetransferofreversibly controlledpolycistronicgenes.MolecularTherapyNucleicAcids2013;2:e85.
DittgenT,NimmerjahnA,KomaiS,LicznerskiP,WatersJ,MargrieTW,Helmchen F,DenkW,BrechtM,OstenP.Lentivirus-basedgeneticmanipulationsofcor- ticalneuronsandtheiropticalandelectrophysiologicalmonitoringinvivo.
ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica 2004;101:18206–11.
DonelloJE,LoebJE,HopeTJ.Woodchuckhepatitisviruscontainsatripartitepost- transcriptionalregulatoryelement.JournalofVirology1998;72:5085–92.

![Fig. 5. Native GFP in the brain of a transgenic mouse (no. 68, P185) expressing GFP and ATXN1[Q98]](https://thumb-ap.123doks.com/thumbv2/123deta/6228647.1091336/8.918.193.707.83.501/fig-native-gfp-brain-transgenic-mouse-expressing-atxn.webp)
![Fig. 6. ATXN1[Q98] aggregates in neurons of various regions primarily affected by SCA1](https://thumb-ap.123doks.com/thumbv2/123deta/6228647.1091336/9.918.167.766.78.816/fig-atxn-aggregates-neurons-various-regions-primarily-affected.webp)