ISSN 0916-4804 総 説
ヒ ト単 球 ・マ ク ロ フ ァ ー ジ の 分 化 と機 能
赤 川 清 子 国立感染症研究所 免疫部 要 旨 ヒ ト単 球 をGM-CSFま た はM-CSF存 在 下 に 培 養 す る と, 形 態, 表 面 マ ー カ ー, 機 能(貧 食 能, 活 性 酸 素 産 生 能, 抗 原 呈 示 能, HIV感 染 感 受 性 な ど)の 異 な る2種 類 の マ ク ロ フ ァ ー ジ(Mφ)に 分 化 す る こ と, GM-CSFで 分 化 誘 導 し たMφは, ヒ トの 肺 胞Mφに 似 て い る こ と が 知 ら れ た. ま たCSFに よ る ヒ ト単 球 のMφへ の 分 化 はIL-4に よ り修 飾 さ れ, GM-CSFとIL-4に よ りCD 1陽 性 の 樹 状 細 胞(DC)に, ま たM-CSFとIL-4に よ りTRAP陽 性 の 破 骨 細 胞 様 多 核 巨 細 胞(MGC)へ の 分 化 が 誘 導 さ れ る こ と が 知 ら れ た. 単 球 由 来DCは, 既 にGM-CSFに よ りMφへ 変 換 す る 能 力 は 有 し て い な い が, M-CSFの レ セ プ タ ー で あ るc-fmsを 有 しM-CSFに 反 応 し てMφに 分 化 可 能 で あ る. し か しTNF-α で 処 理 す る こ と に よ りc.fmsの 発 現 が 抑 制 さ れM.CSFに よ るMφへ の 分 化 能 を 失 う こ とが 知 ら れ た. ま た 単 球 由 来MGCの 形 成 に は 内 在 性 のIL-1とIL-6が 重 要 な 役 割 を 果 た し て お り, CD4/HLA-DR, LFA-1/ICAM-1 及 びCD 14と そ の リ ガ ン ドの 相 互 作 用 が 必 要 な こ と が 示 唆 さ れ た.Key words: 単 球(monocytes), マ ク ロ フ ァ ー ジ (macrophages), サ イ トカ イ ン (cytokine), 樹 状 細 胞 (dendritic cells), 多 核 巨 細 胞 (multi-nucleated giant cells)
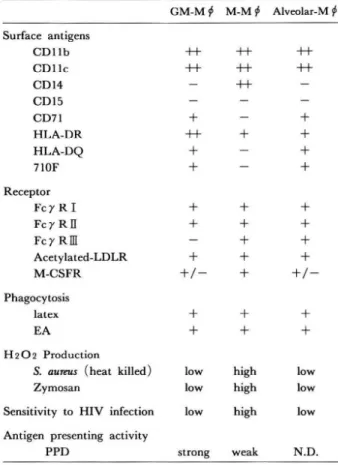
1. は じ め に マ ク ロ フ ァ ージ (Mφ) は, 貧 食 作 用 以 外 に, 殺 菌, 抗 原 呈 示, 腫 瘍 細 胞 傷 害, 分 泌, 骨 の 形 成 と吸 収, 脂 質 の 代 謝 作 用 な ど広 汎 な機 能 を有 し, 感染 防御, 免 疫 応 答, 炎 症作 用 な ど生体 の 防御 機構 や恒 常 性 の維 持 に基 本 的 か つ 重 要 な役 割 を果 た して い る. 一 般 に, Mφは 単 球 に 由 来 す るが, 生 体 の 各 組 織 に存 在 す るMφは, 形 態, 機 能, 細 胞 表 面 抗 原 の 発 現 な どが 異 な り, ク ッパ ー細 胞(肝), 肺 胞Mφ(肺), ミク ロ グ リア(脳), 破 骨細 胞(骨)な ど と異 な る名 前 で 呼 ば れ る. Mφの 分 化, 増 殖, 生 存, 機 能 に直 接 影 響 を及 ぼす 因子 と してmacrophage colony-stimulating factor (M-CSF), granulocyte-macrophage (GM)-CSFそ して interleukin (IL)-3の3つ の コロ ニ ー刺 激 因子(CSF)が あ る. これ らの 因 子 は, 内皮 細 胞, 繊 維 芽 細 胞, 単 球, Mφ, リ ンパ 球 な どか ら産 生 さ れ る. しか し各 種 炎 症, 病 態 の 場 にお い て は, CSF以 外 の 種 々の サ イ トカ イ ン も産 生 さ れ て お り, そ れ らが 局 所 に浸 出 した単 球 や 既 に そ こ に 存 在 す るMφの 増 殖, 分 化, 機 能 に影 響 を与 えて い る と考 え られ る. 本 稿 で は ヒ ト単 球 のMφへ の分 化 と機 能 に対 す る これ らサ イ トカ イ ンの影 響 につ い て述 べ る. 2. ヒ ト単 球 よ りGM-CSF及 びM-CSFに よ り 誘 導 さ れ たMφの 形 態 と表 面 抗 原 に つ い て ヒ ト単 球 をGM-CSFやM-CSFの 存 在 下 に 培 養 す る と, 細 胞 の 大 き さ, プ ラ ス チ ッ ク 面 へ の 粘 着 性 が 増 しMφ へ の 分 化 が 誘 導 さ れ る. 培 地 の み で は 増 殖 もMφへ の 分 化 も 認 め ら れ な い こ と よ り, 骨 髄 前 駆 細 胞 か ら のMφへ の 分 化 同 様, 血 中 単 球 か らMφへ の 分 化 に もCSFが 必 須 で あ る こ と が 知 ら れ た. Fig. 1及 び Table 1に 示 す よ う に, GM-CSFとM-CSF で 誘 導 さ れ たMφ(そ れ ぞ れM型Mφ及 びGM型Mφ と以 下 称 す る)は, 形 態 や 細 胞 表 面 抗 原 の 発 現 が 異 な る. 両Mφと もHLA-DR, CD 11b, CD11cは 発 現 し て い る が, M型Mφは, CD 14を 強 く発 現 す る が, CD71, HLA-DQ, 710Fを 発 現 せ ず, 逆 にGM型Mφは, CD 14は 発 現 せ ず, CD71, 710F, HILA-DQを 発 現 し て い る.ま た GM型Mφで はM-CSFの レ セ プ タ ー で あ るc-fmsの mRNAの 発 現 も低 い. 両Mφと も ス カ ベ ン ジ ャ ー レ セ プ タ ー を 発 現 し, ま た 単 球 同 様Fcγ レ セ プ タ ーI(Fcγ RI), FcγRIIは 発 現 し て い る が, FcγR皿 はM型 Mφの み に 発 現 し て い る. イ ン テ グ リ ン分 子 の 発 現 は 両 者 で 異 な り, M型Mφで は αvβ5が, GM型Mφで は, αvβ3が 発 現 し て い る1). 別 刷 請 求 先: 赤 川 清 子 〒162東 京 都 新 宿 区戸 山1-23-1 国立 感 染 症 研 究 所 免 疫 部
210 真 菌 誌 第38巻 第3号 平 成9年 3. GM-CSFに よ り単 球 よ り誘 導 し たMφと ヒ ト肺 胞 Mφの 関連 先 に我 々 は, M-CSFやGM-CSFが マ ウス 肺 胞Mφの 増 殖 を促 す の み な らず, そ れ ぞ れ性 質 の 異 な る2種 類 の Mφへ と分 化 させ る こ と, Mφ多 様 性 由 来 の 原 因 の 一 つ は少 な く と もMφ形 成 に 関与 す るCSFの 違 い で あ る こ と, 肺 胞 マ ク ロ フ ァ ー ジ の 形 質 はGM-CSFに よ り決 定 され る こ と等 を明 らか に した2). GM-CSFが 肺 胞Mφの 分 化, 機 能 の 発 現 に と り不 可 欠 の もの で あ る こ とは, 1994年 に2つ の グル ー プ に よ り 作 成 され たGM-CSF欠 損 マ ウ ス の結 果 か らも明確 に なっ た. GM-CSF欠 損 マ ウス は, 造 血 系 を は じめ種 々 の 組 織 に著 明 な 変化 は な い が, 肺 は ヒ トの肺 胞 蛋 白症 に似 た 症 状 を示 し, 細 菌 や カ ビの 日和 見 感染 が発 症 す る な ど, 唯 一肺 に異 常 を認 め た3,4). しか しGM -CSF欠 損 マ ウ ス の 肺 にGM-CSFの 遺 伝 子 を導 入 し, GM-CSFを 肺 で 発 現 させ る と肺胞 蛋 白 症 には な らな い5). 即 ち, 肺 胞Mφの サ ー フ ァ ク タ ン ト処 理 能 力 が 導 入 したGM-CSFに よ り 回復 した た め で あ る. Table 1に 示 す よ う に ヒ ト単 球 由 来GM型Mφの 表 面抗 原 やc-fms mRNAの 発 現 パ タ ー ン は, ヒ トの 肺 胞 Mφの そ れ とほ ぼ 同 じで あ り, ま たGM型Mφの 形 態 は ヒ ト肺 胞Mφの 形 態 と似 て い る こ とか ら, ヒ トの 場 合 もマ ウス 同 様GM-CSFが 肺 胞Mφの 形 質分 化 に 重 要 な 役 割 を果 た して い る と推 察 さ れ る. しか し ヒ トの 肺 胞 Mφは マ ウ ス の肺 胞Mφに 比 べ る と増 殖 能 が 極 端 に 弱 く, GM-CSF存 在 下 で も コ ロニ ー形 成 は 認 め られ ず, M-CSFに 対 す る応 答 も ほ と ん ど認 め られ な い6). M-CSF に対 す る応 答 が 弱 い の は, ヒ ト肺 胞Mφで はc-fmsの 発 現 が 非 常 に低 い こ と と一 致 す る. これ らの ヒ ト肺 胞Mφ で認 め られ る性 質 は, ヒ ト単 球 よ り誘 導 したGM型Mφ で も同様 に認 め られ た. 一 方, ヒ ト単 球 由 来M型Mφは, CDI4を 強 く発 現 し, またc-fmsのmRNAを 発 現 す る点 な ど ヒ トの 腹 腔Mφ に似 て い る. 4. CSFで 誘 導 した ヒ ト単 球 由 来Mφの 機 能 に つ い て a) 寅 食 能 M-CSFお よびGM-CSFで 誘 導 した ヒ ト単 球 由 来Mφ はFcγR依 存 性 の 貧 食 能 が 異 な る(Table 1). FcγR には, fgr, hck, lyn, sykな どの チ ロ シ ンキ ナ ー ゼ が 介 合 し, これ に よ るFcγRの リ ン酸 化 が 貧 食 機 能 の 発 現 に 重 要 な こ とが 報 告 され てい る7). そ れ ゆ えGM型Mφの EA(感 作 羊 赤 血 球)の 貧 食 の低 い 理 由 は, チ ロ シ ン キ ナ ー ゼ の 介 合 や そ の活 性 化 にお け る 欠 陥 に 関 連 す る 可 能 性 が 高 い. FcγRIIIはM型Mφに 発 現 して い る が, こ の 発 現 の 違 い と 関 連 して 抗 体 依 存 性 の 細 胞 傷 害 活 性 は M型Mφの 方 が 強 い8). b) 活 性 酸 素 産 生 能 Mφの 殺 菌 作 用 や細 胞 障 害活 性 の エ フ ェ ク ター 分 子 の 一 つ で あ るH2O2の 産生 は, M型Mφの 方 が 高 い. 最 近 我 々 は, H2O2に た い す る 感 受 性 はM型Mφの 方 が GM型Mφに 比 し約10∼100倍 高 い こ と を 見 い だ した.
Table 1 Characteristics of macrophages generated from human monocytes and alveolar macrophages
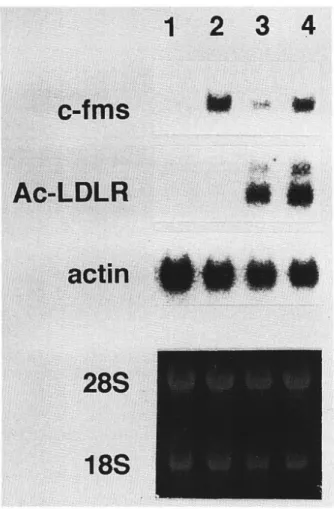
Fig. 1. Morphology of macrophages, dendritic cells and multi-nucleated giant cells derived from human monocytes a . macrophages generated from monocytes by GM-CSF b . macrophages generated from monocytes by M-CSF c. dendritic cells generated from monocytes by GM-CSF
plus IL-4
d. multi-nucleated giant cells generated from monocytes by M-CSF plus IL-4
この事 は, M型Mφは 微 生 物 を貧 食 す る と大 量 の 活 性 酸素 を産 生 し, 殺 菌 に有 利 に働 く と同時 に 自 ら もそ の 活 性 酸 素 で死 ぬ こ と に よ り, 過 剰 の 活性 酸 素 産生 を抑 制 し, 炎 症 の 拡 大 や組 織 の破 壊 を 防 い で い る と考 え られ る. c) HIV感 染 感 受性 ヒ トにAIDZを 起 こすHIVに 対 す る 感 染 感 受 性 も両 Mφは 異 な り, M型Mφで はHIVの 強 い増 殖 が 見 られ るが, GM型Mφで はHIVの 増 殖 が 起 き な い9). HIV の 細胞 へ の 感染 に は レセ プ タ ーで あ るCD4以 外 に, Mφ トロ ピ ックHIVの 場 合 には βケ モ カ イ ン の レ セ プ タ ー で あ るCC-CKR5等 が 補 助 レ セ プ タ ー と して 必 要 で あ る こ とが 最 近 明 らか に な っ た10). 両 マ ク ロ フ ァ ー ジ の CD4の 発 現 に差 は 無 い が, 補 助 レ セ プ タ ーの 発 現 に違 い が あ るか 否 か は今 の所 明 ら か で は な い. しか しHIVの プ ロ ウ イル スDNAはGM型Mφに も存 在 した こ と よ り, GM型MφのHIV感 染 抵 抗 性 は, 補 助 レ セ プ タ ー の 発 現 の 違 い に よる とい うよ り, こ のMφで は何 らか の 理 由 に よ りウイ ル ス遺 伝 子 の活 性 化 が 抑 制 さ れ, い わ ゆ る潜 伏 感 染 の状 態 を と っ て い る た め と考 え ら れ る. 両 Mφに お け るHIV感 染 感 受性 の 違 い は, HIVの 増 殖 及 び 増 殖抑 制機構 の解 明 に有 用 で あ ろ う. d) 抗 原呈 示 活 性 一 方, BCG菌 や そ の 可 溶 性 抗 原 で あ るPPDをT細 胞 に提 示 す る抗 原呈 示 活性 は, GM型MφがM型Mφに 比 べ は るか に強 い. 抗 原 呈 示 に必 要 なHLA■DR抗 原 の 発 現 は 両 者 で あ ま り変 わ ら ない こ と よ り, この 違 い は そ れ 以 外 の 理 由 に よ る と思 わ れ る. こ の 点 に 関 し, 最 近 我 々 は, BCG菌 やPPDを パ ル ス したM型MφはTh 1 細 胞 の 活 性 化 を抑 制 す るIL山0を 大 量 に 産 生 す る が, GM型 マ ク ロ フ ァー ジ は ほ とん ど産 生 しな い こ と, ま た M型Mφで は, T細 胞 活 性 化 の共 刺 激 分 子 で あ るCD 54 (ICAM-1), CD80, CD86な どの 発 現 が ほ とん ど認 め ら れ ない こ と等 を見 つ けた. お そ ら くこ う した 抑 制 因 子 の 産 生 や 共 刺 激 分 子 の 発 現 の 低 さ がM型 マ ク ロ フ ァ ー ジ の抗 原 呈 示 活 性 の 低 さ と関 係 す る と考 え られ る. e) CD14依 存 性 機 能 CD 14は, グ ラ ム陰 性 菌 外 膜 の 主要 構成 成分 で あ るLPS と血 清 中 のLPS結 合 蛋 白LBPと の 複 合 体 を 認 識 す る レ セ プ ター で あ りMφのLPS応 答 に と り重 要 な分 子 で あ る11). ま たCD14は 結 核 菌 の細 胞 壁 成 分 の リポ ア ラ ビ ノ マ ンナ ンを結 合 しTNF-α, ILI-8, NOな どの 産 生 を誘 導 し, ま た ミク ロ グ リアが 結 核 菌 を貧 食 す る と きの レ セ プ ター で もあ り, 単 球 がIL-1に よ り活 性 化 され た 血 管 内 皮 細 胞 に接 着 す る と きの接 着 分 子 で もあ る12).こ のCD 14の 発 現 は 両Mφで 大 き く違 う事 よ り, CD 14が 関 連 し た機 能 も両 者 で大 き く異 な る で あ ろ う. 5. ヒ ト単 球 由来 樹 状 細 胞(DC)の 形 質 DCは, 抗 原提 示 の プ ロ フェ ッシ ョナ ル細 胞 と して 免 疫 応 答 に重 要 な細 胞 で あ る. 特 に1次 免 疫 応 答 に お け る T細 胞 活 性 化 は, DCの み で誘 導 されMφやB細 胞 で は 誘導 され な い. 1992年Couxら に よ りGM-CSFとTNF-αの 共 刺 激 に よ り, CD34陽 性 前 駆 細 胞 か らCD 1陽 性 の皮 膚 の ラ ン ゲ ル ハ ンス細 胞 様DCの 分 化 が 誘 導 され る こ とが報 告 さ れ た13). 我 々 は, ヒ ト単 球 をGM-CSFとIL-4共 存 化 に 培 養 す る と, GM-CSF単 独 培 養 の 場 合 と異 な りMφ へ の分 化 が 抑 制 され, 貧 食 活 性(ラ テ ック ス 粒 子, EA), ス カベ ン ジ ャー レセ プ ター, 非 特 異 的エ ス テ ラ ー ゼ 活 性 な どのMφマ ー カ ー 陰 性 の 非 粘 着 性 のDCの 分 化 が 誘 導 され る こ と を見 い だ した14)(Fig. l). 単 球 由来DCは, 既 に述 べ た 皮 膚 のDCで あ る ラ ンゲ ルハ ンス 細 胞 に特徴 的 と さ れ るCDla抗 原, MHCク ラ スII抗 原 を強 く発 現 し, またDCの 抗 原 呈 示 細 胞 と し て の機 能 に必 要 なCD80, CD86, CD54, CD40な ど の 共 刺 激 分 子 を 強 く発 現 す るが, Mφマ ー カ ー で あ るCD 14, CD71, 710Fを 発 現 しな い. 単 球 由 来DCが 真 のDCで あ る こ とは, こ の細 胞 が, DCの 分 化 に 必 須 で あ る 転 写 因子NF-kBフ ァ ミリ ーの サ ブユ ニ ッ トとの 一 つ で あ る Re1B15,16)を 発 現 して い る こ と, またDCの 機 能 的 特 徴 で あ る 自 己及 び異 系 リ ンパ 球 混 合 培 養 にお け る 静 止期T 細 胞 刺 激 活 性 を有 してい る こ とか ら も明 らか で あ る. こ の よ うにDCはCD34陽 性 前 駆 細 胞 の み な らず 単 球 か ら も分 化 誘 導 可 能 な こ とが 知 られ た. しか しそ の 分 化 誘 導 に必 要 なサ イ トカ イ ンはそ れ ぞ れ 異 な る こ とが知 られた. 6. ヒ ト単球 由 来DCのMφ変 換 能 GM-CSFとIL-4に よ り ヒ ト単 球 か ら分 化 して きたDC は, 単 球 とは 異 な りGM-CSFの 刺 激 に よ り も は やMφ に変 換 しな いが, M-CSFに 応 答 し て プ ラ ス チ ッ ク粘 着 性 及 び ラ テ ック ス粒 子 貧 食 能 を 有 す るMφに 分 化 可 能 で あ る こ と, しか し このDCをTNF-αで 処 理 す る とM-CSFで 刺 激 して もMφへ の 分 化 は 誘 導 さ れ ず 培 地 の み の 場 合 と同 じよ う に死 ん で し ま う14). こ れ は, TNF-α 処 理 に よ りM-CSFの レセ プ ター で あ るc-fmsの 発 現 抑 制 が 起 こ った た め で あ る(Fig. 2)・TNF-α処 理 に よ り DCへ の最 終 分 化 が 誘 導 され る と き, 抗 原 呈 示 能 も増 加 しMLR刺 激 活性 の増 強 が起 こる. こ の増 強 は ク ラスII 抗 原, CD80, CD86な どの共 刺 激 分 子 の発 現 増 強 と一 致 して い た. 7. ヒ ト単 球 よ りのTRAP陽 性 多 核 巨 細 胞 の 分化 我 々 は, ヒ ト単球 をM-CSFとIL4共 存下 に培 養す る と, 2∼3個 か ら10数 個 の核 を有 す る 多 核 巨 細 胞(MGC) が形 成 され る こ と を 明 らか に した14)(Fig. 1). MGC は, 種 々 の感 染, 異 物 反 応, 癌, サ ル コ イ ドー シ ス そ し て リウマ チ な どの 場 にお い て み られ る. また す で に 述 べ た よ うに骨 の吸 収 と形 成 に重 要 な役 割 を果 た して い る破
212 真 菌 誌 第38巻 第3号 平 成9年 骨 細 胞 も, MGCの 形 態 を し て い る. op/opマ ウ ス の 解 析 か ら破 骨 細 胞 形 成 に はM-CSFがMGC形 成 因 子 と し 必 須 な こ と が 知 ら れ て い る が17), IFN-γ, IL-3, GM-CSF, IL-4な ど の サ イ トカ イ ン も ヒ ト単 球 や, マ ウ ス 骨 髄 細 胞, ラ ッ ト ミ ク ロ グ リ ア か ら のMGC形 成 刺 激 活 性 を 有 す る こ とが 報 告 さ れ て い る. し か しGM-CSF, IFN-γ, IL-1α, IL-6, TNF-α は, い ず れ も 単 独 及 びM-CSF 存 在 下 で も ヒ ト単 球 よ りMGC形 成 を 誘 導 せ ず, IFN-γ, GM-CSF, TNF-α は む し ろM-CSFとIL4に よ るMGC 形 成 を 抑 制 し た, ま たM-CSFとIL4に よ るMGC形 成 は, 内 因 性 のIL-1, IL-6が 関 与 す る こ と が 知 ら れ た.し か しM-CSFとIL-1α あ る い はIL-6ま た は 両 者 を 共 存 さ せ て もMGCの 形 成 は 認 め ら れ な い こ と よ り, こ れ ら の サ イ ト カ イ ン は 必 要 だ が 十 分 で な い こ とが 示 唆 さ れ た. 破 骨 細 胞 や 肉 芽 腫 のMGCは, 単 核 の 単 球, Mφが 融 合 して 形 成 さ れ る と 考 え ら れ て い る が, M-CSFとIL-4 に よ る ヒ ト単 球 よ り のMGC形 成 に は, CD 14と そ の 未 知 の リガ ン ド, CD4とHLIA-DR, LFA-1とICAM-1と の接 着 を介 した結 合 が 必 要 で あ る こ とが 知 られ た.こ の こ と は, 単 球 ・Mφ細 胞 間 の 融 合 に よ るMGC形 成 は, 単 球 の 血 管 内 皮 へ の接 着 及 び 抗 原 呈 示 細 胞 とT細 胞 間 の 接 着 の メ カ ニ ズ ム に 良 く似 た もの で あ る こ と を示 す. M-CSFとIL-4に よ りヒ ト単 球 よ り形 成 され たMGC は Table 2に 示 す よ うに, Mφと 異 な りC3レ セ プ ター, Fcレ セ プ ター, 非 特 異 的 エ ス テ ラ ー ゼ 活 性 が 弱 い か ほ とん ど発 現 して い な い が, 破 骨細 胞 の マ ー カー で あ る 酒 石 酸 抵 抗 性 の 酸 フ ォス フ ァ ター ゼ(TRAP)活 性 が 陽 性 で あ り, また こ のMGCはBCG菌 を 良 く貧 食 した. 肉 芽腫 形 成MGCの 機 能 や 形 成機 構 及 び破 骨 細 胞 との 関 連 な ど, 現 在 不 明 な 点 が 多 い が, こ の in vitro にお け る ヒ ト単 球 よ りのMGC形 成 系 は, そ れ らの解 明 に と り有 効 な手 段 とな る で あ ろ う. 8. お わ り に CSFに よ る単 球 か らMφへ の 基 本 的 分 化 は, 種 々 の サ イ トカ イ ン に よ り修 飾 さ れ る. 特 にTh2由 来 のIL-4 が 単 球 か らDCやMGCの 分 化 を誘 導 した こ と は 興 味 深 い. 最 近 我 々 は, Th2由 来 の も う一 つ の サ イ トカ イ ン で あ るIL-10が, これ らM-CSFとGM-CSFに よ る単 球 か らMφへ の分 化 に 対 し, そ れ ぞ れ 分 化 の 増 強 と ア ポ トー シ ス とい う全 く正 反対 の影 響 を 与 え る こ と を見 い だ した18). GM-CSFで 誘 導 したMφは, 抗 原 呈 示 活 性 が 強 い こ と, ま たM-CSFで 誘 導 したMφは, 活 性 酸 素 産 生 能, 貧 食 能, ADCC機 能 な どが 強 い こ と よ り, IL-10は 抗 原 呈 示 機 能 を 有 す るMφの 分化 を抑 制 し, エ フ ェ ク ター 機 能 を有 す るMφの 分 化 を増 強 す る こ とが 示 唆 さ れ た. 多 くの 疾 患 にお い て, そ こ に集 ま りま たそ こで 増 殖, 分 化 したMφが 病 態 の 発 現 に 重 要 な役 割 を 果 た す こ と が 示 Fig. 2. Expression of c-fms mRNA in dendritic cells generated
from monocytes by GM-CSF+IL-4.
Total RNAs (10/1 g/lane) extracted from TNF- a -treated DC (lane 1), TNF- a -non-treated DC (lane 2),
induced macrophages (lane 3) or M-CSF-induced phages (lane 4) were subjected to agarose gel electrophoresis and then transferred to nylon membrane and hybridized with a labeled cDNA probe of c-fms. Rehybridization was
carried out wyth probes specific for Ac-LDLR or for actin.
Table 2 Characteristics of multi-nucleated giant cells from human monocytes by M-CSF+IL-4
唆 され て い る が, そ こ に 関 与 す る サ イ トカ イ ンの 種 類, また そ れ らMφ の機 能 が どの よ う な もの か は必 ず し も 明確 で 無 い. こ こに述 べ た ヒ ト単 球 のサ イ トカ イ ン に よ る分化 の系 は あ く まで も基 本 的 な もので, 生 体 で は よ り 複雑 な サ イ トカ イ ンの種 類 と量 の 組 み 合 わ せ が 起 こ り, Mφ の 分化 及 び そ の機 能 の発 現 に関 与 す る と考 え られ る. 実 際 の病 気 の場 で のMφ の分 化 と機 能 が どの ように なっ て お り, そ れ が病 態 の発 現 に どの よ う に 関 与 す るか, 今 後 よ り明 らか に な っ て くる こ と と思 わ れ る. 文 献
1) De Nichilo MO, Burns GF: Granulocyte-macrophage and macrophage colony-stimulating factors differentially
regulate a v integrin expression on cultured human macrophages. Proc Natl Acad Sci USA 90: 2517-2521,
1993.
2) Akagawa KS, Kamoshita K, Tokunaga T: Effects of granulocyte-macrophage colony-stimulating factor and colony-stimulating factor-1 on the proliferation and differentiation of murine alveolar macrophages. J
Immunol 141: 3383-3390, 1988.
3) Dranoff G, Crawford AD, Sadelain M, et al: ment of granulocyte-macrophage colony-stimulating factor in pulmonary homeostasis. Science 264: 713-716,
1994.
4) Stanley ER, Lieschke GJ, Grail D, et al: Granulocyte/ macrophage colony-stimulating factor-deficient mice
show no major perturbation of hematopoiesis but develop a characteristic pulmonary pathology. Proc Natl Acad Sci USA 91: 5592-5596, 1994.
5) Huffman JA, Hull WN, Dranoff R, et al: Pulmonary epithelial cell expression of GM-CSF corrects the alveolar proteinosis in GM-CSF-deficient mice. J Clin
Invest 97: 649-655, 1996.
6) Nakata K, Akagawa KS, Fukayama M, Hayashi Y, Kadokura M, Tokunaga T: Granulocyte-macrophage
colony-stimulating factor promotes the proliferation of human alveolar macrophages in vitro. J Immunol 147:
1266-1272, 1991.
7) Wang AVT, Scholl PR, Geha RS: Phusical and tional association of the high affinity immunoglobulin G receptor (Fc y R I ) with the kinases Hck and Lyn. J Exp Med 180: 1165-1170, 1994.
8) Yaung DA, Lowe LD, Clark SC: Comparison of the effects of IL-3, granulocyte-macrophage
ing factor, and macrophage colony-stimulating factor in supporting monocyte differentiation in culture. Analysis of macrophage antibody-dependent cellular cytotoxicity. J Immunol 145: 607-615, 1990.
9) Matsuda S, Akagawa K, Honda M, Yokota Y, Takebe Y, Takemori T: Suppression of HIV replication in human monocyte-derived macrophages induced by granulocyte/macrophage colony-stimulating factor. AIDS Research and Human Retroviruses 11: 1131-113 8,
1995.
10) Doranz BJ, Rucker J, Yi Y, et al: A dual-tropic primary HIV-1 isolate that uses fusin and the beta-chemokine receptors CKR-5, CKR-3, and CKR-2b as fusion cofactors. Cell 85: 1149-1158, 1996.
11) Pugin J, Heumann D, Tomasz A, et al: CD 14 is a pattern recognition receptor. Immunity 1: 509-516, 1994.
12) Beekhuizen H, Blokland I, Corsel-van Tilburg AJ, Koning F, van Forth R: CD 14 contributes to the adherence of human monocytes to cytokine-stimulated endothelial cells. J Immunol 147: 3761-3767, 1991. 13) Caux C, Dezutter-Dambuyant C, Schmitt D, et al:
GM-CSF and TNF-a cooperate in the generation of dendritic Langerhans cells. Nature 360: 258-261, 1992. 14) Akagawa KS, Takasuka N, Nozaki Y, Komuro I, Miyuki A, Ueda M, Naito M, Takahashi K: Generation
of CD 1 + Re 1 B + Dendritic Cells and TRAP-positive Osteoclast-like Multi-nucleated Giant Cells from Human Monocytes. BLOOD 88: 4029-4039, 1996.
15) Burkly L, Hession C, Ogata L, Reilly C, Marconi LA, Olson D, Tizaard R, Cate R, Lo D: Expression of re1B
is required for the development of thymic medulla and dendritic cells. Nature 373: 531-536, 1995.
16) Weih F, Carrasco D, Durham SK, Barton DS, Rizzo SA, Ryseck P-P, Lira SA, Bravo R: Multiorgan
inflammation and hematopoietic abnormalities in mice with a targeted disruption of relB, a member of the NF-kB/rel family. Cell 80: 331-340, 1995.
17) Yoshida H, Hayashi S, Kunisada T, et al: The murine mutation osteoetrosis is in the coding region of the macrophage colony stimulating factor gene. Nature
345: 442-443, 1990.
18) Hashimoto S, Yamada M, Motoyosi K, Akagawa KS: Enhancement of macrophage-colony-stimulating induced growth and differentiation of human monocytes by interleukin-10. BLOOD 89: 315-321, 1997.
214 真 菌 誌 第38巻 第3号 平 成9年
Generation
of Two Phenotypically
Distinct
Types of Macrophages,
CD 1+Dendritic
Cells and TRAP+Osteoclast-like
Multi-nucleated
Giant Cells from Human
Monocytes
Kiyoko S. Akagawa
Department of Immunology, National Institute of Infectious Diseases,
1-23-1, Toyama, Shinjuku-ku, Tokyo 162, Japan
We demonstrated that granulocyte-macrophage
colony-stimulating factor (GM-CSF) and
macrophage-CSF (M-csF)
stimulate the differentiation of CD14+human
monocytes into two phenotypically distinct
types of macrophages, based on morphorogy, cell surface antigen expression and functions including
phagocytosis, production of reactive oxygen intermediate,
antigen presenting activity and sensitivity to
HIV infection. In vivo, however, not only CSF but also many other cytokines are produced under
various conditions, and those cytokines may modulate the differentiation of monocytes by CSFs. We
demonstrated that CD 14+human
monocytes can differentiate into CD1+re1B+dendritic
cells (DC)
by combination of GM-CSF plus interleukin-4 (IL-4), and that they differentiate into tartrate-resistant
acid phosphatase (TRAP)-positive
osteoclast-like multinucleated
giant cells (MGC)
by combination of
M-CSF plus IL-4. However, the monocyte-derived DC were not terminally differentiated cells; they
could still convert to macrophages in response to M-CSF. Tumor necrosis factor- a (TNF-a)
stimulated
the terminal differentiation of the DC by down-regulating the expression of the M-CSF receptor,
c-fms mRNA and aborting the potential to convert to macrophages.
Taken together, these results
provide a new aspect to our knowledge of monocyte differentiation, providing evidence that human
monocytes are flexible in their differentiation potential, and that they are precursors not only of
macrophages but also of CD1+re1B+DC
and TRAP-positive MGC.
Such a diverse pathway of
monocyte differentiation may constitute one of the basic mechanisms of immune regulation.
この 論 文 は, 第40回 日本 医真 菌 学 会 総 会 の “ワ ー ク シ ョッ プ: 真 菌 感 染 にお け る マ クロ フ ァー ジ の役 割 ”に お い て 発 表 され た もの で す.