七飯におけるエンレイソウ属植物の交雑 (英文)
9
0
0
全文
(2) Journal of Hokkaido Gagugei University. Vol. 10, No. 2. Dec. 1959. Hybridizadon of Trillium in a Habitat at Nanae Satoshi MATSUZAKA and Masataka KURABAYASHI^* Biological Laboratory, Hakodate Branch, Hokkaido Gakugei University. ^. r^jEi^: -baStc^U-^^y^/f yM^t?®^^. Nanae is a village about 10km. north of the city of Hakodate. JVIarine terraces of Quarternary origin run along the margin of the Ohno Plain of alluvial soil. From the foot of Mt. Yokotsu, a non-active volcano of recent origin, spread gentle slopes terminating a little west of the margin, on which the village is situated. It is patched with secondary woodland of Larix, Crypiomeria, Abies, Chamaecyparis, etc. having an area 1500-5000 m . Trillium form the undergrowth. The mode of habitation of the plants varies with the age, composition and topography of the woods. We can find there many examples of Trillium colonies with various characteristics. Four of the colonies named A, B, C and D, were chosen for the present examination (Fig. 1).. Fig. 1. A map of Nanae Village showing the four colonies, A, B, C, D. The composition of the wood is represented by the following marks. X : Larix. G; Cryptomeria. P : Abies, Picea and Pinus. H : Chamaecyparis. D : deciduous trees.. Results of the Examination Colony A. This colony occupies a flat area of 30 x 80m2 covered with Larix woods 50 years old. A small number of Cormis and chestnut trees are found mixed with them. Young trees of Abies of about 1 m. height were planted recently in the wood. T. kamtschaticum. is the only Trillium found in this colony. A rectangle of 5 x 10 m2 was fixed there (Fig. 2). T. kamtschaticum grows evenly there. Colony B. This is found in Larix woods of 20 years old planted in an area of about 25 x 60 m2. A small number of Aescnlus, walnut and chestnut trees are mixed with Larix. The bed on the northern edge of the wood is inclined northward-down toward a swamp, being depressed about 1m. beneath the bed. Two species of Trillium, T. kamtschaticum and T. ischonoskii are found there. The former grows evenly in the wood, while the distribution of the latter is restricted to the inclined area. There the two species grow together and The present investigation was partly supported by a grant in aid of fundamental scientific researches from the Ministry of Education in Japan. Dedicated to Prof. Hajime Matsuwa and Prof. Yukio Yamada celebrating their Sexagenary birthdays. — 181 —.
(3) Satoshi IVlatsuzaka and Masataka Kurabayashi •. •. ••^B'. • ^•o. •. $•'. ••. ». 9. •a •• •9 • •. •. ••/. 5. «IO m. x. a. • •. ^ "•. %. •. 3. •. ». •X:-. •^-. •. ^(. ••. .1.. •. o. 0. • s • •. x. ^. •:•. (3%<f <?. •. •. •. •. 0. •. ^. •••. •. <30. •. •I. & • •. 8«'. x. 0°. •. •. •. «p. 8 I. a. a. 0. •. —. e. •. •. 0 •. 0. s^ A' <f ». 9. e. •. •. • ' • •. I). a. a •. ••». •. -^ S X 17. x. •••• a. •. d*. -^-,. 9. •«*•. •. a. •. • a. »a-. •. •r •. f. •. X. •:•/.. ••. 9 •®. •. ••. •• • x. •••••' !•. •. ^. 9. ® 99 0. •. % ". •. •. •. ». •°. •. •»• t, « • OB. •. ~f». ••*• '•••. • • 9 0. So. ••-•. :.x.. 3. • 9» • •. 9. •• • •_• ••. •>••. •• x. •. •. • •. •••' '•»•*. 4. S < 50. CD cpo. e. x. x. r,° ^. x. ••:•. x. o 99. a. x. X.. \ .<.. •X. x. X. — • 00^0 •. • • •. x. •. ••. •*L. OD. 0. x. • x x. x. x. x 0. x * •• •a • x •• •••. • x. •• x. •. n ••. ^ 0. x. • x • x •• 09. •. ••-. X. e». x. 0. ••. x •. x. •X.. 0. x. •. x. .*or°.. 0. x. x. x. x. .X. •°So°8(3. ^•.'. x •. x. .<.. QCS> 0 Ox 0. x. ••x 9. x. x. co 0. x. x. x. A. )°)? ^ Q cP%<;)0o 0.. X. ?.?.. •*s*. x. 9. •'. J.
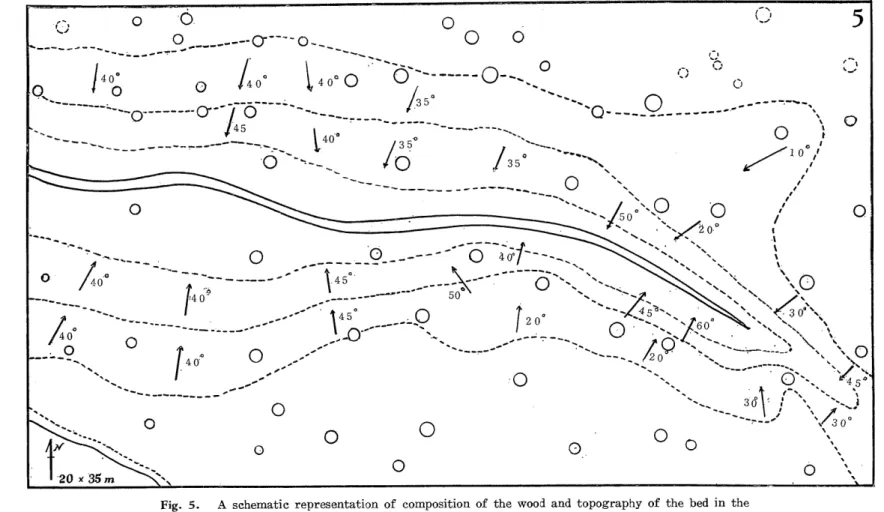
(4) 0. ^—-—cr"'~o^,. 0. 0 0. o o----o-^. 0. 0 Q-—.-^-———-. 0. 0 ). 0. T^^i^^^^. 0. 0 40a7"~~'-^. l41-----^-'"""o~":;:~-'<. 40'. Ho r40'. 0. Q. ^"'/5»:~"° ..°. .(. 40-. ^<:>^_:b\\o. ~"'--:"o-./^> ,' 0'---. :^. 0 0. ^^'/60° ^<'<s\ \. ,1-0-""" 'Y. 0. \©. ~~'^~'~.^<'.N<^\. I450 __-—0. 0. v. ^ '^,. 0. 0 0. "~--Ar"\>:J 0. Fig. 5. A schematic representation of composition of the wood and topography of the bed in the rectangle fixed in colony D. Arrows indicate directions of inclination towards the ditch. The inclination in degrees is written beside each arrow. Entire circle: Cryptomeria. Broken circle : deciduous trees.. '30°. 0 0 0. ffi. t< 0-. 10~/.
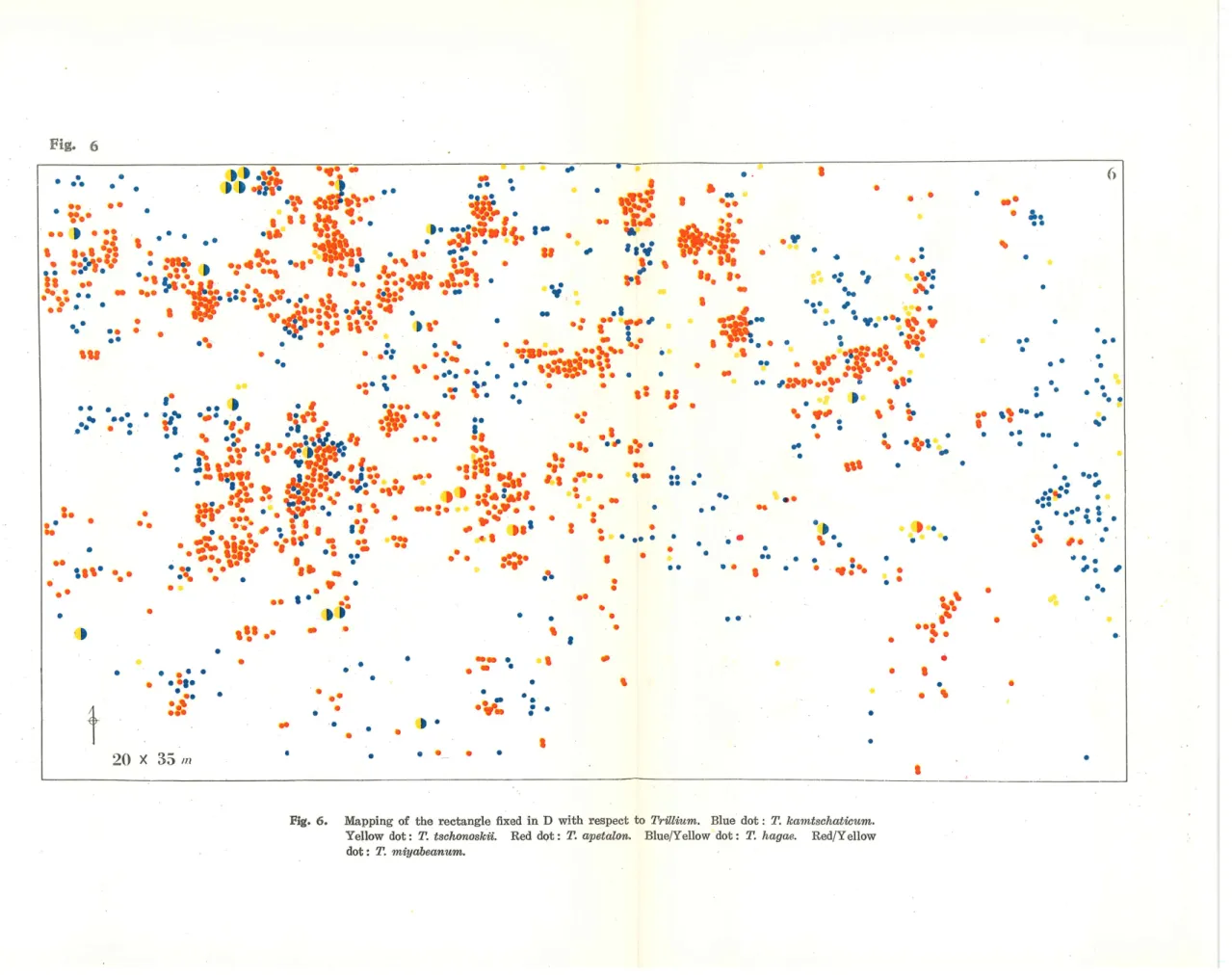
(5) Satoshi Matsuzaka and Masataka Kurabayashi. form a sterile hybrid, T. hagae (3 X). A belt of 5 x 17 m2 was drawn from middle to the northern end of wood. One plant of T. hagae was occasionally found to be included in the northern part of the belt where mixed habitation of the two species was seen (Fig. 3). Colony C. A pathway runs a little north to the railway station from east to weat (Fig. 1). A young wood of Larix (6 x 60m2) planted four years ago grows along the southern side of the pathway. There is a shallow ditch 1 m. wide and 50 cm. deep running for 46 m. in the wood. Big chestnut trees with diameter of 30-40 cm. are planted in a row on the southern side of the wood at intervals of about 5 m. A belt of 5 x 50 m2. was fixed in the wood. Two kinds of Trillizim, T. kamtschaticum and T. tschonoskii were found there. They do not grow mixed together more or less separated from each other forming, species by species, aggregates in the wood (Fig. 4). A single plant of T. apetalon was found in the middle of the belt. No hybrid plants were detected. Colony D. There are woods of mixed composition on the northern side of the pathway mentioned above. They occupy an area of 45 x 55m2. A deep branching ditch, cuts the western three quarters of the forest bed. Cryptomeria trees with a diameter of about 30 cm. grow in the central part of the forest. Its peripheral zone is composed of Larix, Cormis, Morus, etc. The bed, being variously inclined (from 10° to 60°) toward the ditch, is occupied densely by Trilliwn. A rectangle of 20 x 35 m2 was fixed in the central part (Fig. 5). Three species and two interspecific hybrids of Trillitim grow there (Fig. 6). A total of 501 plants of T. kamtschaticnm were found over all the area examined. Their distribution is more dense in the peripheral zone of the rectangle than in the central part of it. The frequency of T. tschonoskii is much smaller than that of the former. Only 93 plants were found scattered at random in the rectangle. The above two species occa-. sionally produce a hybrid, T. hagae (3X). A total of 17 plants of this hybrid were detected. T. apetalon forms dense aggregates in shady dark niches along the ditch. This species forms a hybrid, T. miyabeanum, with T. tschonoskii. Four plants of the hybrid were found on the edges of the ditch. Each of the two kinds of hybrids is intermediate in character between the parental species and shows vigorous vegetative growth. They are both complately sterile (Haga 37', '56, Kurabayashi and Saho '57). Discussion It is evident from the examination of the four colonies that the mode of habitation of Trillitim differs markedly from colony to colony. Woods of simple composition in a flat area are occupied a single species of Trillium. Sparse woods of Larix are suitable for habitation of T. kamtschaticum which likes semi-shade. The Abies trees planted in the Larix wood in colony A are too small to effect the light in such a way as to prevent T. kamischaticnm there. The flat area of the Larix wood in colony B is occupied only by this plant. A topographical change in the bed of the latter wood affords a niche for T. tschonoskii which grows an inclined area, while T. kamtschaticum prefers a flat one. Mixed habitation of these species in the inclined zone in B has promoted hybridization between them. The two species coexist also in colony C. However, they live there more or less separated each forming independent aggregates. Such discontinuous distribution may have been brought about by uneveness in shade given by the chestnut trees which are planted at intervals of 5 m. on the southern side of this habitat. Topographical variation induced by the shallow —184 -.
(6) ••. fc. •••--'•. -. •. •• l-•••••••• t?»w-'r. • • ' - ' ,:v.»^'*\ •;• % N ^ •• • • _• _ •• fS^9***y ~s tt» , • ~* ' •7 %'* •• ' ' tS •: -—-. ^ ^- ^ -, ^. -••9. *_t. »f. ••. •. ~. ~^. •. •. y. I. •. '. *. ,^. '•. ^>':'«c.':::':. % ^'^ ,. .; , " ^'r, »l? ;.*»:- ... ^ ^J^'^l^ '" ..& A"- ^^' "-'' L \'frt; . ' ': " • • •sit^« ~ T%S.^»?- - •tSS\. . 99:^ - • nt • • •. ^iw^ jsSSy9\^ :^9 'stitl&»* /t<—* »~ :: •• lw a ' *••••.. L-. .-. »;l?-.€?'?:'A'-:-'>"^^'.r.:-'';'"'•••--••..' . &. \\: " -M^^ ? ;? /^'l \":: -. ••. ^ •1>- ^ "^'. [w... ^^ 'v^r'" •'' ^ ^ \ • ^ '•" :' ••/^ •/ "';:'®. I''-'. ^. •'. .*.. ^^. ...:";':e'. tV-. •. •. ». .A. •.."••! ,. ^. ". ^. •. ... "...^'.. ••^. • —— •. .'V: '.:. •.. •. .. *'. Fig. 6. Mapping of the rectangle fixed in D with respect to TrUlium. Blue dot : T. kamtschatwum. Yellow dot: T. tschonoskn. Red dot: T. apetalon. Blue/Yellow dot: T. hagae. Red/Yellow dot: T. miyabeanum.. •".
(7) Hybridization of Trillizim in a Habitat at Nanae ditch may also be responsible for the uneven habitation. This area, therefore, can be devided into zones varying discontinuously from each other in respect of the daily supply of sunlight, inclination of the bed and humidity. The Larix wood was planted only four years ago. Then afterwards the seeds of each species of Trillitim may have germinated in spots suitable for the germination. Difference in preferance for the above mentioned conditions among the species (cf. Kurabayashi and Saho 57', Shibukawa and Kurabayashi '59) may result in the discontinuous aggregation of different species in different spots. No hybrid plant was found in this colony, because the parental species have lived there only for four years or less in separate groups. The woods in which colony D lives are the ones most complex in composition and in topography of the bed among the woods distributed in Nanae. So also is the mode of habitation of Trillizim found there. The density of T. kamtschatictim is larger in the peripheral zone which is less shady and more flat than the central part where Crypiomeria grows. T. tschonoskii occupies shady and inclined areas which are less humid than depressions in the ditch where T. apeialon aggregates densely. The composition of the wood and the topogra-. phy of the bed is so complex that the bed is divided into minute niches heterogeneous in respect to shade, humidity and inclination. Such situations allow the three species to live more or less mixed together in spite of the difference in ecological preference among them. The seemingly random distribution of T. kamtschaiictim and T. ischonoskii in this colony may not, therefore, be the result of a lack in ecological preference but be based on the mosaic character of this habitat. Non-random distribution of T. apetalon is due to its demand for humid situations, forming dense aggregates in depressions of the ditch. Analogus mode of habitation can often be met with in habitats where the three species coexist (Kurabayashi and Saho '57), Colony D is supposed to be one of the best examples revealing a model of their coexistence in condensed form. The mixed habitation allows hybridizafcion among the three species. The frequency of hybrid plants was the highest ever examined by us. The high density of the three species and their intimate mixture may. facilitate hybridization among them. The complexities in the habitat may provide niches Suitable for the hybrids, which demand ecological conditions intermediate between the parental species. The ecological isolation among the three species has been, as described above, broken down in this colony. Seasonal isolation has been effective in preventing hybridization between T. kamtschaticum and T. apetalon. The former is the latest and the latter is the earliest in flowering among the three. The frequency of hybridization is much higher between T. kamtschaticum and T, tschonoskii than between the latter and T. apetalon*. This difference is supposed to be due to difference in frequency of cross-pollinations between the two pairs of parents. T. apetalon, being an apetal species, is seldom visited by pollinators. They are land snails and small insects which generally cover small areas. It seems, therefore, that the mixed habitation of parental species may be one of the necessary conditions for produc-. tion of hybrids (cf. Shibukawa and Kurabayashi '59).. * The ratio of the ability for production of the hybrids between the two pairs of parents (T. kamtschatikzimXT. tschonoskii and T. tschonoski-ixT. apetalon) will be: 17/(501X93): 4/(1116><93), 9: 1, provided that the three species were equally mixed. 185—.
(8) Satoshi IVIatsuzaka and Masataka Kurabayashi. A Preliminary Note on the Mechanism of Isolation among Japanese Species of Trillium. Summing up the findings hitherto obtained concerning the mechanism of isolation among Japanese species of Trillium (cf. Kurabayashi '58, Kurabayashi and Saho '57, Shibukawa and. Kurabayashi '59) following formulas will be given (cf. Stebbins '50 pp. 189-250), 1. Difference in ecological preference. a) Demand for light: kamtschatiGum'>hagae (6X)> tschonoskii ==8mallii'> apetalon. b) Demand for humidity : apetalon>ka,mt8chatieum~> hagae (6 ~X.)=:tschonoskii=smallii. External Barries. c) Ability to grow on steep slopes: tsconosfciC> apetalon^>smallii==hagae (6 'X)^>kamtschatiGzi,m. 2. Difference in flowering time apetalon kam-ts chatiGum. tschonoskii sin allii ha g ae (6 X) early. —». late. Hybrid sterility (chromosomal) Internal Barries. a) Univalent and multivalent formation b) Non-dis junction due to in version bridges c} Random assortment of different genomes in gametes. We are much obliged to Professor Hideji Yamaguchi for giving us kind encouragement and to the staffs of our laboratory for their co-operation in examining the habitat. Thanks are also due to Professor Makoto Numata, Chiba University, for his valuable criticism and advice.. Summary Four of the colonies of Trilliwn found in Nanae, a village about 10 km. north of the city of Hakodate, were chosen for examination of their mode of their habitation. Each of the colonies grows in woods of various composition and topography. Thin Larix woods planted in a flat area are occupied by a colony composed of T. kamischaticum only. Larix woods with ups and downs in their bed allow mixed habitation of T. kamfschatictim and T. ischonoskii. The latter is found to be restricted to inclined areas in the bed. The two species occasionally produce hybrid plants when the two are intimately mixed. Mixed habitation of three species T. kamtschaticum, T. tschonoskii and T. apeialon was found in a forest composed of Cvyptomeria, Larix, and several kinds of deciduous trees.. Three quarters of the bed are cut by a branching ditch. Thus the bed is divided into a variety of small niches with differences in shade, humidity and topography. Seemingly random mixture of the three species in this forest is due to the complexities in ecological conditions in the bed. Two kinds of hybrids are produced there among the three species.. Summing up the findings hitherto obtained, a preliminary note on the mechanisms of isolation among Japanese speciec of Trillium was given.. —186-.
(9) Hybridization of Tr'illium in a Habitat at Nanae. Literature. Haga, T,, 1937 : Genome and polyploidy in the genus Trillmm. I. Chromosome affinity between the genomes. Jap. J. Gen. 13 :135-145. ", 1956 : do VI. Hybridization and speciation by chromosome doubling in nature. Heredity, 10 : 85-98. Kurabayashi, M., 1958 : Evolution and variation in Japanese species of Trillium. Evolution, 12 : 286310. Kurabayashi, M. and T. Saho, 1957a : Evolution and variation in Trillium. IX. Chromosome comple-. meat in two interspecific hybrids newly found. Cytologia, 22: 263-272. and —, 1957b : do XI. Mechanism of chromosomal isolation in Japanese Trillizwz.. Cytologia, 22: 273-286. Shibukawa, S. and M. Kurabayashi, 1959: Ecological analysis of a Trillium habitat found in Mt. Hakodate. Jour. Hokkaido Gakugei Univ., Sec. B. 10: 176-180. Stebbins, G. L., 1950: Variation and evolution in plants. Colombia Univ. Press.. —187—.
(10)
図

関連したドキュメント
Instead an elementary random occurrence will be denoted by the variable (though unpredictable) element x of the (now Cartesian) sample space, and a general random variable will
A wave bifurcation is a supercritical Hopf bifurcation from a stable steady constant solution to a stable periodic and nonconstant solution.. The bifurcating solution in the case
In this work we give definitions of the notions of superior limit and inferior limit of a real distribution of n variables at a point of its domain and study some properties of
Zhang; Blow-up of solutions to the periodic modified Camassa-Holm equation with varying linear dispersion, Discrete Contin. Wang; Blow-up of solutions to the periodic
After proving the existence of non-negative solutions for the system with Dirichlet and Neumann boundary conditions, we demonstrate the possible extinction in finite time and the
For computing Pad´ e approximants, we present presumably stable recursive algorithms that follow two adjacent rows of the Pad´ e table and generalize the well-known classical
This problem becomes more interesting in the case of a fractional differential equation where it closely resembles a boundary value problem, in the sense that the initial value
It is thus often the case that the splitting surface of a strongly irreducible Heegaard splitting of a graph manifold can’t be isotoped to be horizontal or pseudohorizontal in
