Regulation of Cell Motility by La-Related
Protein 4 in Ovarian Cancer Cell
著者
MAHY NABIL MAHMOUD EGIZ
学位授与機関
Tohoku University
学位授与番号
11301甲第19138号
Doctoral dissertation
Regulation of Cell Motility by La-Related
Protein 4 in Ovarian Cancer Cell
(卵巣癌細胞の運動性の抑制因子としての La
関連タンパク質 4)
Tohoku University, Graduate School of Medicine
Doctorate course
Reproductive and Developmental Medicine
Gynaecology
Mahy Nabil Mahmoud Egiz, MD
2
INDEX
(I)
Acknowledgment ……… (4)
(II)
List of abbreviations………… (6)
(III) Abstract
……… (7)
(IV) Background of research…… (10)
(V) The purpose of research ……… (21)
(VI) Research methods ……… (23)
(VII) Results of research ………… (30)
(VIII)
Discussion ….……… (35)
(IX)
Summary & Conclusion … (42)
(X)
References ……… (45)
3
索引
(I) 謝辞………(4)
(II) 略語のリスト………(6)
(III)要約……… (7)
(IV)はじめに………(10)
(V)研究の目的…………(21)
(VI)試料と方法…………(23)
(VII)結果………(30)
(VIII)討論………(35)
(IX)まとめと結論………(42)
(X)参考文献………(45)
(XI) フィギュア………(50)
4
ACKNOWLEDGMENT
I would like to express my deepest gratitude and appreciation to Prof. Dr. Nobuo Yaegashi, Professor, and chairman of Department of Obstetrics and Gynecology, Tohoku University Graduate School of Medicine, for his guidance, supervision, and kind direction of the work of this thesis and for my whole stay in Japan.
I am greatly indebted to Dr. Kazuyuki Kitatani and Dr. Masafumi Toyoshima Department of Obstetrics and Gynecology, Tohoku University Graduate School of Medicine, who have devoted much of their time and effort throughout the creation of this work and for reviewing the whole work with me. It is an honor for me to be educated by them.
I wish to conduct my special thanks to all staff members of the Department of Obstetrics and Gynecology, Tohoku University Graduate School of Medicine for support, encouragement, and friendly behavior.
Finally, my limitless love and appreciation to the persons who had been suffering with me during the whole period of creation of the current work, my little family.
My husband who spared no effort supporting me and our family, besides being a good doctorate student at the Neurosurgery department, Tohoku University.
5
I owe apology to my two sons, Mazen and Malek as this work superimposed on their rights of refreshment and recreation with Mum.
To the memory of my dear lovely father, my mother, sister, brother, and all my big family who stand behind me push me always forward and being not satisfied except by excellence and perfection of the work despite long space in-between us.
6
LIST OF ABBREVIATIONS
LARPs The La-related proteins
RBPs RNA binding proteins
mRNA Messenger ribonucleic acid
SSB Sjögren syndrome type B antigen
LAM La motif
RRM RNA recognition motif
EOC epithelial ovarian cancer
IRES internal ribosome entry site
RPMI Roswell Park Memorial Institute
PI3K phosphatidylinositol-3-kinase
DMEM Dulbecco’s Modified Eagle’s Medium
SDS-PAGE sodium dodecyl sulfate–polyacrylamide gel electrophoresis
8
I.
ABSTRACT
The La-related proteins (LARPs) are a family of RNA binding proteins that
modulate the degradation and stabilization of RNAs. Multiple studies implicated the dysregulation of LARPs in cancer progression. Cell motility is known to potentiate the metastatic potential of ovarian cancer cells. However, the roles of LARPs in cell motility remain unknown. In the present study, we investigated the roles of LARPs in the progression of ovarian cancer using SKOV3 human ovarian cancer cells and a public database that integrates microarray-based gene expression data and clinical data. To explore the involvement of LARPs in the cell motility, we performed RNA interference screening for LARPs in SKOV3 cells. The screening identified LARP4 as a potential suppressor of the formation of lamellipodia. Conversely, enforced expression of LARP4 suppressed the formation of lamellipodia. Moreover, cell migration was significantly increased in LARP4-knocked-down SKOV3 cells. Mechanistically, LARP4 knock-down was associated with the increase in RhoA protein expression. These results suggest that LARP4 may limit RhoA-dependent cell motility. In a mouse xenograft model with SKOV3 cells, the peritoneal metastasis was increased with LARP4 knock-down increased. Upon analysis of a public database of patients with ovarian cancer, the LARP4 mRNA-high expression group (n = 166) showed longer overall survival compared with the LARP4 mRNA-low expression group (n = 489), implying a positive correlation of LARP4 mRNA levels in ovarian cancer tissues with patient prognosis. In conclusion, LARP4 is proposed to suppress the motility and metastatic potential of ovarian cancer cells.
9
11
II. BACKGROUND OF RESEARCH
1. Epithelial ovarian cancer (EOC)
EOC is the most lethal gynecologic malignancy and the fifth most common cause of cancer-related death in women 1. Each year, EOC is responsible for
over 140,000 deaths worldwide. It is the sixth most common cause of cancer mortality amongst women in developed countries, and the most lethal gynecological malignancy 1. The morbidity of ovarian cancer in Japan was
9,384 women in 2012. The mortality of ovarian cancer was 3.2 per 100,000 in 2012 2. In 2014, 43.2% of ovarian cancer patients were classified as stage I,
9.1% as stage II, 27.6% as stage III, and 7.2% as stage IV 3.
The lethality of ovarian carcinoma primarily stems from the late diagnosis of the patients at the advanced stages with widely metastatic disease within the peritoneal cavity. Ovarian cancers detach from the primary tumor and then exfoliated cancer cells are transported throughout the peritoneum by peritoneal fluids and disseminate within the peritoneal cavity. 4. Thus, it is important to
understand the mechanisms by which ovarian cancer cells acquire metastatic properties to identify new potential molecular targets.
2. Mechanism of metastasis (1) Cell motility
In cancer progression, upregulated cell motility is a prerequisite for metastasis; cell migration involves reorganization of the F-actin cytoskeleton leading to the formation of membrane protrusions at the cell front known as lamellipodia. They are typically elongated at the leading edge of moving cells 5.
12 Lamellipodia are transient structures, which protrude and retract. If the lamellipodium persists long enough, cells can establish new integrin-mediated adhesions with the underlying substratum, contract and then detach adhesions at the rear; this series of events forms the basis of protrusion-based migration 6.
Lamellipodia are critical for directional cell migration and are responsible for cell motility, which contributes to metastatic potential in cancer cells 7, 8. Thus, in the
present study, we utilized ovarian cancer SKOV3 cells as they are characterized by their high ability to form lamellipodia and migrate.
(2) Epithelial mesenchymal transition (EMT)
The initial cellular events required for metastasis are triggered by a switch from an epithelial cell type to a less differentiated mesenchymal one, which is known as EMT. Cells undergoing EMT reorganize their cytoskeletons and extend protrusions such as lamellipodia, allowing for increased migratory capacity. Those events could lead to cancer invasion and metastasis 9. Therefore,
uncovering the regulatory mechanisms of lamellipodia formation is of critical importance to understanding cancer metastasis and discovering novel therapeutics.
(3) Signaling pathways for ovarian cancer metastasis
Previous reports demonstrated the pivotal role of multiple molecular signaling pathways in tumorigenesis and cancer progression. Accumulating evidence is emerging that the phosphatidylinositol 3-kinase (PI3K)–Akt pathway plays important roles in cancer cell growth and motility 10. This pathway is frequently
13 cancer cells, PI3K–Akt pathway has been shown to drive the formation of lamellipodia 12,13.
RhoA, a member of the Ras superfamily, also modulates a diverse set of biological responses of cancer cells, including cell motility, proliferation, apoptosis inhibition, cell cycle progression, invasion, and metastasis in cancer cells 14. Accumulating evidence has shown that RhoA activity is upregulated in
most human carcinogenesis 15. Some studies proposed possible involvement of
RhoA in the progression of ovarian cancer. RNA binding proteins (RBPs) and cancer
Expression profiling has demonstrated that there are substantial differences of mRNA expression between ovarian malignancies and non-cancer tissue, and even between different EOC subtypes 16. There is a growing appreciation that
such differences in the expression are not only determined at the genetic or epigenetic level, but can be the result of post-transcriptional regulation 17.
RNA binding proteins (RBPs) bind transiently to RNAs and resolve trapped, misfolded conformers of RNAs 18, thereby regulating stability and translation of
mRNAs 19-21. These proteins have been considered as RNA chaperone 22.23.
RBPs are increasingly being recognized as key drivers of cancer progression and chemo-resistance in several cancers 24, including ovarian malignancies 25.
mRNA Binding Proteins and cancer 3. La-related proteins
The La-related proteins (LARPs) are an ancient family of a highly conserved RBPs with an RNA chaperon activity that regulates mRNA stability. The first
14 family member to be identified was SSB/La (Sjögren syndrome type B antigen (SSB)/genuine La protein), now known as LARP3. Six LARP subfamilies have been identified in human: LARP1, LARP3/SSB/La, LARP4, LARP4b, LARP6 and LARP7. All family members share a unique N-terminal or central RNA-binding domain, named the La motif (LAM), separated from an RNA recognition motif (RRM) by a short linker. Human LARP3 and LARP7 also possess an additional RRM. The La module recognizes terminal UUU-3’OHs on small, nascent RNA transcripts 23.
Previous studies demonstrated the implication of LARPs in different cancers. For example, Sommer et al. 26 revealed that over-expression of La drives cell
migration and invasion in squamous cancers of the head and neck.In hepatocellular cancer cells, La has been shown to drive epithelial to mesenchymal transition (EMT) through expression of the internal ribosome entry site (IRES)-mediated target Laminin B1 27. LARP1 has been also implicated in
the tumorigenesis of cancer cervix and ovarian cancer 28. Moreover, LARP1
expression in tumor tissues was correlated to poor prognosis in patients with colorectal cancer 29, In contrast, LARP7 is a potential tumor suppressor protein in
gastric cancer 30. For LARP4, little is known about its role in ovarian cancer but it
is documented as one of the novel regulators for the cytoskeletal organization 31
and also as a driver for prostatic cancer cell migration 32.
(1) LARP3/SSB and cancer
Originally identified as a serological marker of autoimmune disease, particularly Sjögren’s syndrome 33, LARP3/SSB has been studied in much
15
greater depth than other members of the LARP family. Accumulating evidence supports a role for LARP3 in cancer development and progression. The gene expression is upregulated in cervical cancer tissues and oral squamous cell carcinomas compared to normal counterpart tissues 26, 34, as well as in multiple
cancer cell lines 35.
LARP3 was found to preferentially regulate the translation of over 200 transcripts, including key cancer-related genes such as VEGF, PDGFA, and BCL2L11 36. Knockdown of LARP3 is associated with reduced migration and
invasion 26. Therefore, it appears that LARP3 acts as a key oncogenic protein 37.
Potential inhibitors of the LARP3-RNA interaction have recently been identified, and found to have biological activity in the inhibition of hepatitis B viral antigen production 38. This suggests that it may be possible to generate anti-cancer
therapies based around the disruption of the function of LARP3 in neoplastic cells.
(2) LARP1 and cancer
In humans, there are two LARP1 proteins, LARP1 and LARP1b, whilst Arabidopsis and other plants have a third gene, LARP1c 39. LARP1 was highly
expressed in hepatocellular cancer (HCC)-derived cell lines compared to a benign cell line 40. In addition, higher LARP1 protein levels were seen in HCCs
compared to non-malignant adjacent liver tissues. LARP1 was identified as an independent predictor of reduced overall survival in multivariate models: patients with high LARP1 expression had a 25% increased risk of death at any time. This survival association was greater than that seen for tumor size or number,
16
indicating a highly clinically significant trend. The ability of LARP1 protein levels to predict outcome outperformed the current gold standard circulating biomarker, alpha-feto protein (AFP) 40.
At the level of cell biology, LARP1 was proposed to play a role in promoting cell motility by interacting with cytoskeletal components. LARP1 protein was concentrated at the leading edge of migrating cells41. LARP1 knockdown
inhibited cell proliferation and induced apoptosis in some types of cancer cell lines studied so far 41, 42.
LARP1 has been identified as a downstream target of the oncogenic PI3K signaling cascade43. Significantly, LARP1 has also been independently identified
in two separate studies as a downstream phospho target of mTORC1 signaling 44, 45, a pathway frequently activated in cancer, and capable of promoting cancer cell
invasion and metastasis 46.
(3) LARP6 and cancer
The first paper describing LARP6 was published in 2007, where it was identified in a screen of genes expressed in intersegmental muscles (ISM) of the moth Manduca sexta during programmed cell death at the end of metamorphosis
47. LARP6 appears to play a role in myogenesis 48, 49.Overexpression of LARP6
in highly metastatic MDA-MB-231 breast cancer cells enhanced proliferation and invasion, associated with increased expression of matrix metalloproteinase-9 (MMP-9) and vascular endothelial growth factor (VEGF). LARP6-overexpressing cells generated significantly larger tumors in vivo, compared to controls, with a
17 significant increase in VEGF expression and tumor angiogenesis 50. Therefore,
LARP6 was proposed to serve as an oncogenic protein.
(4) LARP7 and cancer
LARP7 may play a key role in suppressing malignant transformation. Analysis of LARP7 expression in breast cancer tissues demonstrated that low LARP7 expression was associated with poor overall survival of patients 51. Moreover,
LARP7 knockdown in breast cancer cell lines promoted cell motility and enhanced invasion and cell metastasis in vivo.
LARP7 expression was also shown to be significantly reduced in the tissues of gastric cancers, compared to normal tissue. In non-malignant gastric cells, LARP7 knockdown led to increased cell proliferation and enhanced migration, associated with decreased abundance of 7SK RNA 30. LARP7 expression was
also downregulated in tumors from patients with lymph node-metastatic cervical cancer, compared to those with localized disease 52, suggesting a potential role
in limiting cancer metastasis.
(5) LARP4, 4b and cancer
The LARP4 subfamily is the least studied to date. Two LARP4 genes are present in the human genome, LARP4 and LARP4b (previously LARP5) 39, which
share 37% amino acid identity and 53% sequence similarity 53. LARP4 and 4b
show less La Motif conservation with LARP3 than LARP7, particularly in the side chains needed for 3’UUU-OH binding 23, 53, suggesting they may interact with
18 different RNA targets. In contrast to LARP3 and 7, LARP4 and LARP4b are predominantly cytoplasmic proteins 54, 55.
Both LARP4 and LARP4b possess a PAM2-like sequence in the N-terminal region 55. PAM2 motifs are conserved 15 amino acid sequences involved in
binding to polyA binding proteins (PABPs) at the PABPC domain, and are found in other PABP-interacting proteins such as eukaryotic translation termination factor 3 (eRF3) which play a critical role in mRNA translation and modulating mRNA stability 56, 57, 58. LARP4 and LARP4b were also shown to bind RACK1 54, 55, a scaffold protein that interacts with the 40S ribosomal subunit 59. Supporting
a role in translation, LARP4 and LARP4b were co-sedimented with polyribosomes, and the knockdown resulted in a 20-40% decrease in overall protein synthesis 54, 55. LARP4 has also been suggested to promote mRNA
stability 55.
Only few studies have reported possible involvement of the LARP4 in cancer biology and progression. Knockdown of LARP4 in prostate cancer-derived PC3 cells produced elongated, bipolar cells with increased cell motility 32. The patterns
of expression and significance of both proteins in cancer cells remain to be determined.
4. Study hypothesis
Over the last two decades, significant progress has been made in determining the origins of ovarian cancer and in understanding the molecular
19 basis of its development. Unfortunately, we now know ovarian cancer to be a complex and genetically highly heterogeneous disease.
Although our increased knowledge of the dysregulated pathways in EOC has led to trials of targeted agents, these have so far met with limited success 60.The
central role of post-transcriptional regulation in the development of malignancy is an area of growing research interest. It has been suggested that the majority of the regulation of gene expression is determined post-transcriptionally 17, which
has important implications for our understanding of cancer development and neoplastic cell plasticity. The journey from transcription of nascent mRNAs to their translation in the cytoplasm is a highly complex and tightly regulated process, some aspects of which we are only just beginning to clarify. At the heart of determining RNA fate are a large and diverse family of RBPs. More highly expressed that other regulatory proteins 24, there may be over 1,500 genes
encoding proteins with RNA-binding properties. To date, the functional role of relatively few RBPs has been studied in significant depth LARP3/SSB was one of the first RBPs to be characterized. Over forty years of research has revealed it to participate in an extensive array of functions, from promoting mRNA translation to ensuring correct mRNA folding. We now recognize LARP3 as a member of a family of highly conserved proteins that share similar RNA-binding motifs. LARPs play diverse roles within normal cells, but are increasingly being recognized as significant to cancer development and progression.In particular, LARP4 appears to regulate both mRNA stability and translation, and, at the level of cell biology,
20 promotes cell motility and regulate cell morphology. This suggests that LARP4 expression may be beneficial to the limitation of cancer cell metastasis.
I hypothesize that the LARP4 may play a key role in malignant suppression, and may therefore function as a tumor suppressor in several cancer types, including ovarian cancer. Ovarian cancer is disease in which the silent metastasis leads to late discovery, worse patient prognosis and high mortality rate. LARP4 may potentially act as a metastasis suppressor biomarker, controlling cell behaviors such as cell motility and cell morphology. If this is the case, it may have potential as a survival biomarker and potential therapeutic target.
22
III.
THE PUROPOSE OF RESEARCH
To date, little studies have investigated LARP4 in cancer, and no study explored in-depth the mechanism by which LARP4 may limit cancer progression. The aim of this PhD project were therefore to investigate the role of LARP4 in ovarian cancer metastasis being a silent killer. This will provide a novel biomarker for ovarian cancer metastasis improving patient prognosis, overall survival and quality of life.
24
IV.
RESEARCH METHODS
Antibodies and reagentsAntibodies specific for LARP4 (ab156687), phosphatidylinositol-3-kinase (PI3K) C2β (ab55589), and PTEN (ab32199) were from Abcam (Cambridge, MA, USA). Antibodies specific for JNK (#99252S), JNK (#4668S), phospho-mTOR (#2971S), p110α (#4249), E-cadherin (#3195P), vimentin (#5741P), slug (#9585S), Rac 1/2/3 (#2465T), CDC42 (#2466T), RhoA (#2117S), Akt (#9272S), and phospho-Akt (#9271S) were from Cell Signaling Technology (Boston, MA, USA). Horseradish-peroxidase-conjugated antibodies for rabbit IgG (sc2004) and mouse IgG (sc2005) were from Santa Cruz Biotechnology (Dallas, TX, USA). RNAiMax, Lipofectamine 2000, control siRNAs, LARP4 siRNAs (LARP4 si1, CCAUGGUACAAGUUGAUGAtt; LARP4 si2, CACCAAAGUUUG ACUUAUUtt; LARP4 si3, GGAUGGUCUCAAUCAGACAtt), and other siRNAs for the LARP family members were from Life Technologies (Carlsbad, CA, USA). Tetramethylrhodamine isothiocyanate (TRITC)-conjugated phalloidin and β-actin antibodies (A5441) were from Sigma (St. Louis, MO, USA). Hoechst 33342 was from Dojindo (Kumamoto, Japan). SuperSignal West Dura Extended Duration Substrate and Halt Phosphatase Inhibitor Cocktail were from Thermo Fisher Scientific (Rockford, IL, USA).
Cell culture
Human SKOV3 ovarian cancer cells were grown in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% fetal bovine serum (FBS). These SKOV3 cells are ovarian cancer cells with high ability to form lamellipodia
25
and migrate, therefore they are good candidate cells for our study. Human PC3 prostate cancer cells were cultured in Roswell Park Memorial Institute medium (RPMI) supplemented with 10% fetal bovine serum. Cells were maintained at < 80% confluence under standard incubator conditions (humidified atmosphere, 95% air, 5% CO2, 37°C). No mycoplasma contamination was observed. Basal LARP4 expression in SKOV3 cells compared to other cell lines should be performed in future studies.
Preparation of LARP4 expression vector
Human LARP4 cDNA segments in a cDNA library of SKOV3 cells were amplified by PCR. The PCR products and pcDNA3.1/ V5-His empty vector (ThermoFisher Scientific) were digested with BamH1/XhoI and each fragment was ligated, forming pcDNA3.1/ V5-His LARP4 vector.
Transfection with siRNAs or LARP4 vector
For siRNA transfection, cells (2 × 104) were grown on 35-mm glass-bottomed
dishes and then transfected with 5 nM siRNAs using RNAiMax transfection reagent (Life Technologies) for 48 h. For transfection with plasmid vectors, SKOV3 cells (5 × 104) grown on 35-mm glass-bottomed dishes were transfected
with 2 µg of pcDNA3.1/V5-His empty vector or pcDNA3.1/V5-His LARP4 vector for 24 h using Lipofectamine 2000.
Analysis of lamellipodium formation
SKOV3 cells were grown in 35-mm glass-bottomed dishes at a density of 2 × 104 cells/dish. After transfection, cells were washed with Phosphate-buffered saline (PBS) twice, fixed with 4% formaldehyde for 10 min, and treated with 0.1%
26
TritonX-100 for 10 min, followed by staining with Hoechst 33342 and TRITC-conjugated phalloidin for 5 min. For the analysis of lamellipodia, samples were examined using confocal microscopy. Lamellipodia are thin and veil-like extensions at the edge of cells that contain a dynamic array of actin filaments and are biologically characterized by the enrichment of F-actin. Cells were counted as having formed lamellipodia if there was an increase in visualized F-actin in the lamellipodia. The percentage of cells forming lamellipodia was determined by counting more than 200 cells.
Immunoblotting analysis
Cells were washed three times with PBS supplemented with Halt Phosphatase Inhibitor Cocktail and then lysed using Laemmli sample buffer (Wako, Tokyo, Japan). After brief centrifugation, the protein content was determined using the BCA protein assay reagent. Then, equal amounts of proteins (5 or 10 µg) were loaded for sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) (4%-20% gradient gels). Proteins were electrophoretically transferred to nitrocellulose membranes, blocked with PBS/0.1% Tween 20 (PBS-T) containing 5% nonfat dried milk for 30 min, washed with PBS-T, and incubated with primary antibodies for LARP4 (1 in 1,000 dilution), p110α (1 in 1,000 dilution), PI3KC2β (1 in 1,000 dilution), Akt (1 in 1,000 dilution), phospho-Akt (1 in 1,000 dilution), phospho-mTOR (1 in 1,000 dilution), PTEN (1 in 1,000 dilution), ERK1/2 (1 in 1,000 dilution), phospho-ERK1/2 (1 in 1,000 dilution), p38δ (1 in 1,000 dilution), JNK1/2 (1 in 1,000 dilution), phospho-JNK1/2 (1 in 1,000 dilution), E-cadherin (1 in 1,000 dilution),
27
vimentin (1 in 1,000 dilution), Slug (1 in 1,000 dilution), Rac (1 in 1,000 dilution), CDC42 (1 in 1,000 dilution), RhoA (1 in 1,000 dilution), or β-actin (1 in 50,000 dilution). The membranes were washed three times with PBS-T and then incubated with a secondary antibody conjugated with horseradish peroxidase (1:5,000 dilution) in PBS-T containing 5% nonfat dried milk for 1 h. Proteins were visualized using Supersignal West Dura Extended Duration Substrate (Thermo Fisher Scientific). Quantification of the chemiluminescent signals was performed with a digital imaging system (ChemiDoc, Bio-Rad, Hercules, CA, USA).
Quantitative real-time PCR
RNA was extracted using the RNeasy Mini Kit (Qiagen, Hilden, Germany), in accordance with the manufacturer’s manual. After reverse transcription of RNA with SuperScript III (Thermo Fisher Scientific). quantitative real-time PCR was performed using the StepOne Plus Real-Time PCR System. TaqMan probes specific for LARP1 (Hs00391726_m1), LARP3 (Hs04187362_g1), LARP4b (Hs00299621_m1), LARP6 (Hs00217969_m1), LARP7 (Hs00277883_m1), and β-actin (4333762F) were employed. β-Actin was used as an internal control.
Cell viability assay
Cell viability was assessed with the CellTiter-Glo Luminescent Cell Viability Assay (Promega). Cells were seeded on 96-well plates at a density of 1 × 103
cells in 100µl /well. Preparation of control wells containing medium without cells to obtain a value for background luminescence. After incubation for the indicated periods, equilibration of the plate and its contents at room temperature for approximately 30 minutes. Then addition of a volume of CellTiter-Glo® Reagent
28
equal to the volume of cell culture medium present in each well. Mixing contents for 2 minutes on an orbital shaker to induce cell lysis. Incubation at room temperature for 10 minutes to stabilize luminescent signal. Finally recording luminescence.
Cell migration assay
For SKOV3 cells, cell migration was determined by a Transwell migration assay, as described by Zhang et al. (2018), while for PC3 prostate cancer cells the incubation time was extended up to 24 h, as described by Seetharaman et al. (2016).
Invasion assay
Cells (5 × 104 cells/well) in serum-free medium were seeded onto the upper
chamber of the Transwell with 8-mm pores that were coated with 5% Matrigel. The lower chamber was filled with 10% FBS-containing DMEM. After 18 h of incubation, non-invading cells at the top of the membrane filter were removed using a cotton swab. Invading cells attached to the bottom of the membrane filter were fixed in 100% methanol and then stained with 1% toluidine blue for 5 min. The cell invasiveness was determined by counting invading cells in optical microscopy images (at least ten fields for each determination).
Xenograft model with human ovarian cancer cells
SKOV3 cells (5 × 106 cells/mouse) were transfected with 5 nM of control si2
or LARP4 si2 for 24 h and then inoculated into the peritoneal cavity of female nude mice (BALB/c; Charles River Japan, Yokohama, Japan) to determine peritoneal metastatic potential. The mice were sacrificed at 4 weeks after
29
inoculation, and the number of overt metastases (> 1 mm) was quantified. All animal studies were approved by the Institutional Animal Care and Use Committee of Tohoku University, Japan.
Kaplan-Meier Plotter analysis
The prognostic value of LARP mRNA expression in ovarian cancer was analyzed using an online database, Kaplan-Meier Plotter (http://www.kmplot.com). Online analysis of the database that integrates microarray-based gene expression data and clinical data was performed, leading to the creation of Kaplan-Meier survival plots. Patients were split into two groups with low (black line) and high (red line) gene expression by an automatically selected optimal cut-off. All possible cutoff values between the lower and upper quartiles were computed and the best performing threshold was used.
Statistical analysis
Data in bar graphs are presented as the mean ± standard error (SE) of at least three independent experiments. Statistical analyses were performed using GraphPad Prism 5.03 (California corporation, USA). All P values less than 0.05 were considered significant.
31
V.
RESULTS OF RESEARCH
Identification of LARPs that regulate cell motilityTo identify LARPs responsible for modulating the formation of lamellipodia, we used siRNAs specific for six LARPs (LARP1 si, LARP3 si, LARP4 si2, LARP4b si, LARP6 si, and LARP7 si). The effectiveness of siRNAs for individual LARPs was confirmed in SKOV3 ovarian cancer cells via knockdown of mRNA expression (Fig. 1A). As shown in Fig. 1B and C, knockdown of LARP4 or LARP4b had stimulatory effects on the formation of lamellipodia. LARP4 appeared to have particular potential to suppress the formation of lamellipodia. In contrast, LARP3 knockdown had inhibitory effects on their formation, while knockdown of other LARPs (LARP1, LARP6, and LARP7) had no significant effects. To further evaluate the potential of LARP4 as a suppressor of cell motility, we used an additional siRNA sequence (LARP4 si1) and also constructed a human LARP4-overexpressing vector (pcDNA3.1/V5-His LARP4). Transfection of SKOV3 cells with either LARP4 si1 or LARP4 si2 was confirmed to suppress LARP4 proteins (Fig. 2A) and significantly increased the formation of lamellipodia (about 1.5- to 2-fold compared with that in the control) as shown in Fig. 2B. Reciprocally, LARP4 overexpression by transfection with pcDNA3.1/V5-His LARP4 vector suppressed the formation of lamellipodia to 0.4-fold compared with that of pcDNA3.1/V5-His empty vector (Fig. 3A-C). Knockdown of RhoA partly attenuated the formation of lamellipodia in SKOV3 cells (Fig. 4). These results suggest that LARP4 suppresses the RhoA-dependent formation of lamellipodia. In addition, the effects of LARP4 knockdown on the Transwell
32
migration of SKOV3 cells were assessed. Similar to the results on the formation of lamellipodia, LARP4 knockdown with individual sequences (LARP4 si1 and LARP4 si2) significantly increased cell migration by 1.35- and 1.85-fold compared with that upon the control knockdown, respectively (Fig. 5A and B). Thus, LARP4 may serve as a suppressor of the motility of ovarian cancer cells. On the other hand, LARP4 knockdown had no significant effects on the invasiveness of SKOV3 cells (Fig. 5C and D). We next tested the effects of LARP4 siRNA on the migratory and invasive activities of PC3 prostate cancer cells. LARP4 knockdown promoted both cell migratory and invasive activities (Fig. 6A-D), consistent with a previous report 32. These results suggest a
difference in the suppressive effect of LARP4 on the invasiveness between ovarian cancer and prostate cancer. In addition, identification of basal LARP4 expression in SKOV3 cells, compared to another cell lines, should be described in future studies.
LARP4 suppresses RhoA expression
Class II PI3K β isoform (PI3K C2β) has been implicated in the formation of lamellipodia in SKOV3 cells 61, 62. To determine the molecular mechanisms by
which LARP4 suppresses cell motility, we evaluated the effects of LARP4 knockdown on PI3K C2β and its associated signaling pathways in SKOV3 cells (Fig. 7). The PI3K-Akt-mTOR pathway was unlikely to be involved in the regulatory mechanisms of LARP4-controlled cell motility. Moreover, the involvement of MAPK and epithelial- mesenchymal transition (EMT) pathways 63, 64 was assessed. No changes in the expression of those pathway- associated
33
proteins were observed in LARP4-depleted SKOV3 cells (Fig. 7). Tumor suppressor genes such as P53 and PTEN are associated with ovarian cancer progression 65. We thus tested the possible involvement of LARP4 in regulating
the expression of these tumor suppressor proteins. As shown in Fig. 7, LARP4 knockdown had no noticeable effects. Rho GTPases are intracellular signal transducers implicated in the control of actin cytoskeleton organization, cell migration, and invasion 66, 67. We investigated the expression of Rho GTPases in
LARP4-knocked-down SKOV3 cells. LARP4 knockdown resulted in a significant increase in RhoA protein expression (Fig. 8A and C), while the expression of Rac and CDC42 did not change (Fig. 8A, E and G). These results suggest that LARP4 suppresses RhoA expression. The sufficient LARP4 knockdown was calculated and plotted in fig. 8A, B, D and F.
Anti-metastatic potential of LARP4 in a mouse xenograft model of ovarian cancer
To assess the involvement of LARP4 in the metastasis, we employed a xenograft model of SKOV3 ovarian cancer cells 62 and evaluated the effects of
LARP4 knockdown on metastasis. LARP4 siRNA was confirmed to persistently achieve knockdown for up to 96 h (Fig. 9A). LARP4 knockdown caused a significant increase in the number of peritoneal metastatic nodules compared with that upon control knockdown (Fig. 9B). LARP4 was, therefore, suggested to suppress the metastatic potential of ovarian cancer cells.
Correlation of LARPs mRNA expressions with the overall survival probability in ovarian cancer patients.
34
To assess the relevance of LARP expression to the prognosis of patients with
ovarian cancer, we performed an online analysis using ovarian cancer microarray datasets 68. Overall survival graphs of ovarian cancer based on LARPs
expression were performed using publicly available database (KM plotter; www.kmplot.com) with the following options: auto select best cutoff, overall survival (OS), 120 month, the plot include addy-id and sample number. cut off values were as follow: LARP1 (19), LARP4 (253), LARP6 (300), LARP3 (1738), LARP4B (25), LARP7 (813). Patients were divided into two groups, with high and low LARP expression. Overall survival periods were plotted to generate Kaplan-Meier curves. As shown in Fig. 10A-F, the LARP4-high patient group showed longer overall survival than the LARP4-low group, suggesting a positive correlation of LARP4 mRNA in ovarian cancer tissues with patient prognosis. Conversely, LARP3 mRNA expression was inversely correlated with overall survival (P < 0.007). No significant correlation of overall survival with the mRNA expression of other LARPs was observed (Fig. 10).
36
VI. DISCUSSION
In this study, we provide the first report that shows the regulatory role of RNA chaperon for lamellipodia formation, we characterized for the first time that LARP4 has a suppressive potential in ovarian cancer metastasis by reducing the migratory activity of ovarian cancer cells through limiting lamellipodia formation. However, no effect was observed on cell growth nor invasion in LARP4 depleted cells. Our data, for the first time, uncovered Rho A protein as a target for LARP4. We demonstrated the anti-metastatic role of LARP4 in the xenograft model, equally as important we demonstrated the positively correlated LARP4 mRNA expression to higher patient overall survival. Together, these results support our hypothesis that LARP4 is a tumor metastasis suppressor protein and provide an evidence for the possible future role of RNA chaperon as a tumor predictor biomarker.
In a former report, LARP4 was identified as one of several novel regulators of prostate cancer cell morphology 31. Furthermore, Seetharaman et al
demonstrated that LARP4 depletion decreases cell circularity and increases cell perimeter in PC3 cells 32. Those data suggested that LARP4 influences
lamellipodia formation in prostatic cancer but for ovarian cancer, no obvious data show the role of LARP4 in lamellipodia formation and cell migration, thus our concern is to illustrate this role and provide a clear evidence for the tumor regulatory potential of LARP4 in ovarian cancer.
Our results demonstrated that LARP4 knockdown significantly up-regulated lamellipodia formation while LARP4 overexpression significantly downregulated
37
lamellipodia formation (figure 3). We demonstrated that LARP4 significantly suppresses cell motility, which is consistent with the limitation of lamellipodia formation. Seetharaman et al. demonstrated that LARP4 depletion increased cell motility and invasiveness in PC3 prostate and MDA-MB-231 breast cancer cells
32 and RhoA was reported to promote cell motility and invasiveness in those cell
lines 69-71. In our study, LARP4 is suggested to target RhoA-dependent cell
motility but not invasiveness in ovarian cancer cells. Though this discrepancy remains unknown, the regulatory mechanism of invasiveness in SKOV3 cells might be different from those of PC3 and MDA-MB-231 cells. Collectively, our data indicated that LARP4 has a selective suppressive activity of cell motility in SKOV3 ovarian cancer cells.
The molecular mechanism by which LARP4 can regulate lamellipodia formation and motility in ovarian cancer is still uncovered. Therefore, I hypothesized that LARP4 acts via dysregulation of cell motility related proteins which in turn will regulate lamellipodia formation, motility and finally ovarian cancer metastasis. Cell motility regulators in ovarian cancer includes many signaling pathways with a prominent role for PI3K-AKT-Mtor, MAPK signaling pathways 62, 72. Accumulating evidence is emerging that the phosphatidylinositol
3-kinase (PI3K)–Akt pathway plays important roles in cancer cell growth and motility, and this pathway is frequently active in ovarian cancer and is proposed to be a therapeutic target 10. Therefore, I tested PI3K-AKT-mTOR signaling
pathway as a target for LARP4 however there was no effect of LARP4 depletion on the expression of these proteins (fig.7). MAPK proteins’ expressions were not
38
changed by LARP4 depletion in SKOV3 cells. Thus I investigated the downstream effector of these pathways which are the Rho GTPase signaling pathway proteins.
In the present study, LARP4 is thought to regulate the RhoA expression, transcriptionally, translationally and/or post-translationally, while the molecular mechanisms by which LARP4 down-regulates RhoA protein expression remain unknown. LARP4 has an RNA chaperon activity by which LARP4 stabilize or destabilize the target mRNA regulating its stability and activity 23, therefore the
effect of LARP4 depletion in SKOV3 cells on RhoA mRNA expression was tested however there was a change in the mRNA expression of the housekeeping genes resulting in difficult normalization and inaccurate interpretation of the results. Therefore, using other housekeeping genes as ribosomal mRNA in future studies might be beneficial to evaluate LARP4 effect on RhoA mRNA levels in ovarian cancer cells. Rho GTPase signaling is controlled by multiple regulators, including Rho GTPase-specific guanine nucleotide exchange factors, GTPase activating enzymes and Rho guanine nucleotide dissociation inhibitors 66, 67.
Those proteins are known to control RhoA activity, however we did not determine it in the present study. Further investigations need to be performed for uncovering the molecular mechanisms.
RhoA is widely implicated in the progression of numerous human cancers including breast, prostatic, testicular and esophageal cancers 70, 73-77. Similarly,
RhoA has been emerging as a key player in the progression of ovarian cancer associated with metastasis 78-81. Considering those studies, the regulatory
39
system of RhoA by LARP4 is suggested to play an important role in ovarian cancer progression.
Of the LARPs, LARP3 was shown to promote cell proliferation in cervical, prostatic and hypopharyngeal cancers 26, 34. LARP3 was also shown to promote
cell migration in hypopharyngeal squamous adenocarcinoma cells, although pathobiological roles for LARP3 in ovarian cancer remained fully unknown. We provide an evidence that LARP3 promotes cell motility in ovarian cancer cells (Fig. 1B and C). In addition, we explored the clinical importance of LARP3 mRNA expression in ovarian cancer progression (Fig. 10B). Altogether, LARP3 might serve as an oncogenic RBP in ovarian cancer.
We further investigated the possible molecular targets of LARP4 in motility regulation. First, we investigated the major molecular pathways involved in regulating cell migration as the PI3K-AKT-mTOR, MAPK and EMT pathways, our results denoted that LARP4 targets other pathways in regulating cellular migratory activity. Next, we hypothesized that LARP4 regulates cytoskeleton proteins such as Rho GTPases, which are intracellular signal transducers that act as the downstream for the previously mentioned major pathways and is strongly implicated in controlling actin cytoskeleton organization and cell migration in mammalian cells 66.In their study Akiko Horiuchi et al showed that
Rho A has a high mRNA expression in ovarian cancer cells and is associated with tumor progression and invasiveness 79, therefore we confirmed the
stimulatory role of Rho A for lamellipodia formation using Rho A depletion experiment (figure 4) then we investigated the possible effect of LARP4 on Rho
40
GTPases demonstrating that there was a significant enhancement of Rho A protein expression with LARP4 depletion as shown in figure 8A and C. In consequence, for the first time, we provide an evidence that LARP4 regulates cellular motility through targeting Rho A which is a well-established key player protein in cellular motility 81.
To further investigate the metastatic potential of LARP4 we established, for the first time, a LARP4 depleted xenograft model. Our results showed that there was a significant increase in the number of metastatic peritoneal nodules relative to the control which provide an evidence of the metastatic suppressor role of LARP4 and therefore delineate the future therapeutic role of LARP4 in ovarian cancer (Fig 9). To validate these results, I checked the persistence of LARP4 knockdown in SKOV3 cells by western blotting up to 96 h as the first few days are the most important in the development in metastatic nodules, more confirmation using the Hematoxylin-Eosin staining of the tumor tissue sample or visible metastatic nodule was not performed in the current study but it would be helpful in future studies.
The LARP4 gene is located on human chromosome 12q13.12. Over 130 LARP4 mutations in cancers are reported in the COSMIC. Of these, six mutations are located in the C-terminal region of LARP4, a part of the protein that appears to mediate interactions of LARP4 55. In seetharaman et al study they
investigated these LARP4 mutants in PC3 cells, none of the LARP4 mutations affected protein stability and localization. They reported that several cancer-associated mutations in LARP4 enhance its effects on cell shape, predominantly
41
by reducing cell elongation and increasing circularity, which would be predicted to reduce migration and invasion based on their previous studies with PC3 cells
82. This suggests that some of the mutants could be more active than wild-type
LARP4 32.
To summarize, we provide the first report demonstrating that only 3 isoforms of the LARP family can significantly regulate lamellipodia formation, from these three proteins LARP4 is a potent down-regulator for lamellipodia formation. We provide the first evidence that Rho A is a target for LARP4. Furthermore, our data elucidate the tumor metastasis suppressor role of LARP4, RNA chaperone, in ovarian cancer via establishing a novel LARP4 depleted xenograft model. Together the in vitro and in vivo results are consistent with the Kaplan Meier survival plot generated by www.kmplot.com/ovar which illustrates the positive correlation between LARP4 mRNA expression and ovarian cancer patients overall survival. All these data allow us to propose that LARP4 has a tumor suppressive potential in ovarian cancer metastasis and may itself behaves as a prognostic factor and a novel therapeutic target to explore in the future.
To arrive at a complete description of the action of LARP4 in ovarian cancer cells, it is necessary to determine if the protein is regulating RhoA activity, whether this regulation is transcriptional or posttranscriptional regulation and to describe if LARP4 is interacting directly or indirectly with RhoA protein. Future studies are required to illustrate and answer these questions.
43
VII.
SUMMARY
The La-related proteins (LARPs) are a family of RNA binding proteins that
control the degradation and stabilization of RNAs. As emerging research reveals the biology of each LARP, it is evident that LARPs are dysregulated in some types of cancer. Upregulation of cell motility potentiates the metastatic potential of ovarian cancer cells; however, the roles of LARPs in cell motility remain unknown. In the present study, we investigated the roles of LARPs in the progression of ovarian cancer using SKOV3 human ovarian cancer cells and a public database that integrates microarray-based gene expression data and clinical data.
To explore the involvement of LARPs in the cell motility, we performed RNA interference screening for LARP mRNAs in SKOV3 cells. The screening identified LARP4 as a potential suppressor of the formation of lamellipodia. Conversely, enforced expression of LARP4 suppressed the formation of lamellipodia. Moreover, cell migration was significantly increased in LARP4-depleted SKOV3 cells. Mechanistically, LARP4 depletion was associated with the decrease in RhoA protein expression. These results suggest that LARP4 may limit RhoA-dependent cell motility. In a mouse xenograft model with SKOV3 cells, LARP4 depletion potentiated peritoneal metastasis. Upon analysis of a public database of patients with ovarian cancer, the LARP4 mRNA-high expression group (n = 166) showed longer overall survival compared with the LARP4 mRNA-low expression group (n = 489), implying a positive correlation of LARP4 mRNA levels in ovarian cancer tissues with patient prognosis. Taken together, we
44
propose that LARP4 could suppress motility and metastatic potential of ovarian cancer cells.
VIII. CONCLUSIONS
LARP4 pathobiology in ovarian cancer is still unclear and we demonstrate an anti-metastatic role of LARP4 in preclinical studies and clinical significance of LARP4 expression in cancer progression. In light of these results, LARP4 is possible to serve as a novel prognostic biomarker in ovarian cancer.
In conclusion, LARP4 is suggested to function as a RhoA-driven motility suppressor in ovarian cancer (fig.11).
46 IX. REFRENCES
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA: a cancer journal for clinicians. 2018;68(1):7-30.
2. Torre LA, Bray F, Siegel RL, et al. Global cancer statistics, 2012. CA: a cancer journal for clinicians. 2015;65(2):87-108.
3. Amin RW, Ross AM, Lee J, et al. Patterns of ovarian cancer and uterine cancer mortality and incidence in the contiguous USA. The Science of the total environment. 2019;697:134128.
4. Yeung TL, Leung CS, Yip KP, et al. Cellular and molecular processes in ovarian cancer metastasis. A Review in the Theme: Cell and Molecular Processes in Cancer Metastasis. American journal of
physiology Cell physiology. 2015;309(7):C444-456.
5. Guan X. Cancer metastases: challenges and opportunities. Acta Pharm Sin B. 2015;5(5):402-418. 6. Petrie RJ, Doyle AD, Yamada KM. Random versus directionally persistent cell migration. Nature reviews Molecular cell biology. 2009;10(8):538-549.
7. Bisi S, Disanza A, Malinverno C, et al. Membrane and actin dynamics interplay at lamellipodia leading edge. Curr Opin Cell Biol. 2013;25(5):565-573.
8. Machesky LM. Lamellipodia and filopodia in metastasis and invasion. FEBS Lett. 2008;582(14):2102-2111.
9. Roche J. The Epithelial-to-Mesenchymal Transition in Cancer. Cancers. 2018;10(2).
10. Xue G, Hemmings BA. PKB/Akt-dependent regulation of cell motility. Journal of the National Cancer Institute. 2013;105(6):393-404.
11. Bast RC, Jr., Hennessy B, Mills GB. The biology of ovarian cancer: new opportunities for translation. Nature reviews Cancer. 2009;9(6):415-428.
12. Ip CK, Cheung AN, Ngan HY, et al. p70 S6 kinase in the control of actin cytoskeleton dynamics and directed migration of ovarian cancer cells. Oncogene. 2011;30(21):2420-2432.
13. Yoshizaki H, Mochizuki N, Gotoh Y, et al. Akt-PDK1 complex mediates epidermal growth factor-induced membrane protrusion through Ral activation. Molecular biology of the cell. 2007;18(1):119-128. 14. Kwiatkowska A, Symons M. Signaling determinants of glioma cell invasion. Advances in
experimental medicine and biology. 2013;986:121-141.
15. Takami Y, Higashi M, Kumagai S, et al. The activity of RhoA is correlated with lymph node metastasis in human colorectal cancer. Digestive diseases and sciences. 2008;53(2):467-473.
16. Konstantinopoulos PA, Spentzos D, Cannistra SA. Gene-expression profiling in epithelial ovarian cancer. Nature clinical practice Oncology. 2008;5(10):577-587.
17. Schwanhausser B, Busse D, Li N, et al. Corrigendum: Global quantification of mammalian gene expression control. Nature. 2013;495(7439):126-127.
18. Wurth L. Versatility of RNA-Binding Proteins in Cancer. Comparative and functional genomics. 2012;2012:178525.
19. Glisovic T, Bachorik JL, Yong J, et al. RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett. 2008;582(14):1977-1986.
20. Raineri I, Wegmueller D, Gross B, et al. Roles of AUF1 isoforms, HuR and BRF1 in ARE-dependent mRNA turnover studied by RNA interference. Nucleic acids research. 2004;32(4):1279-1288.
21. Rajkowitsch L, Chen D, Stampfl S, et al. RNA chaperones, RNA annealers and RNA helicases. RNA Biol. 2007;4(3):118-130.
22. Liao B, Hu Y, Brewer G. Competitive binding of AUF1 and TIAR to MYC mRNA controls its translation. Nat Struct Mol Biol. 2007;14(6):511-518.
47 23. Bayfield MA, Yang R, Maraia RJ. Conserved and divergent features of the structure and function of La and La-related proteins (LARPs). Biochim Biophys Acta. 2010;1799(5-6):365-378.
24. Kechavarzi B, Janga SC. Dissecting the expression landscape of RNA-binding proteins in human cancers. Genome biology. 2014;15(1):R14.
25. Guo Y, Tian P, Yang C, et al. Silencing the double-stranded RNA binding protein DGCR8 inhibits ovarian cancer cell proliferation, migration, and invasion. Pharmaceutical research. 2015;32(3):769-778. 26. Sommer G, Rossa C, Chi AC, et al. Implication of RNA-binding protein La in proliferation,
migration and invasion of lymph node-metastasized hypopharyngeal SCC cells. PloS one. 2011;6(10):e25402.
27. Petz M, Them N, Huber H, et al. La enhances IRES-mediated translation of laminin B1 during malignant epithelial to mesenchymal transition. Nucleic acids research. 2012;40(1):290-302.
28. Hopkins TG, Mura M, Al-Ashtal HA, et al. The RNA-binding protein LARP1 is a post-transcriptional regulator of survival and tumorigenesis in ovarian cancer. Nucleic acids research. 2016;44(3):1227-1246.
29. Ye L, Lin ST, Mi YS, et al. Overexpression of LARP1 predicts poor prognosis of colorectal cancer and is expected to be a potential therapeutic target. Tumour Biol. 2016;37(11):14585-14594.
30. Cheng Y, Jin Z, Agarwal R, et al. LARP7 is a potential tumor suppressor gene in gastric cancer. Laboratory investigation; a journal of technical methods and pathology. 2012;92(7):1013-1019. 31. Bai SW, Herrera-Abreu MT, Rohn JL, et al. Identification and characterization of a set of
conserved and new regulators of cytoskeletal organization, cell morphology and migration. BMC biology. 2011;9:54.
32. Seetharaman S, Flemyng E, Shen J, et al. The RNA-binding protein LARP4 regulates cancer cell migration and invasion. Cytoskeleton (Hoboken). 2016;73(11):680-690.
33. Reichlin M. Current perspectives on serological reactions in SLE patients. Clinical and experimental immunology. 1981;44(1):1-10.
34. Sommer G, Dittmann J, Kuehnert J, et al. The RNA-binding protein La contributes to cell proliferation and CCND1 expression. Oncogene. 2011;30(4):434-444.
35. Al-Ejeh F, Darby JM, Brown MP. The La autoantigen is a malignancy-associated cell death target that is induced by DNA-damaging drugs. Clinical cancer research : an official journal of the American Association for Cancer Research. 2007;13(18 Pt 2):5509s-5518s.
36. Brenet F, Socci ND, Sonenberg N, et al. Akt phosphorylation of La regulates specific mRNA translation in glial progenitors. Oncogene. 2009;28(1):128-139.
37. Trotta R, Vignudelli T, Candini O, et al. BCR/ABL activates mdm2 mRNA translation via the La antigen. Cancer cell. 2003;3(2):145-160.
38. Tang J, Huang ZM, Chen YY, et al. A novel inhibitor of human La protein with anti-HBV activity discovered by structure-based virtual screening and in vitro evaluation. PloS one. 2012;7(4):e36363. 39. Bousquet-Antonelli C, Deragon JM. A comprehensive analysis of the La-motif protein superfamily. Rna. 2009;15(5):750-764.
40. Xie C, Huang L, Xie S, et al. LARP1 predict the prognosis for early-stage and AFP-normal hepatocellular carcinoma. Journal of translational medicine. 2013;11:272.
41. Burrows C, Abd Latip N, Lam SJ, et al. The RNA binding protein Larp1 regulates cell division, apoptosis and cell migration. Nucleic acids research. 2010;38(16):5542-5553.
42. Tcherkezian J, Cargnello M, Romeo Y, et al. Proteomic analysis of cap-dependent translation identifies LARP1 as a key regulator of 5'TOP mRNA translation. Genes & development. 2014;28(4):357-371.
43. Matsuoka S, Ballif BA, Smogorzewska A, et al. ATM and ATR substrate analysis reveals extensive protein networks responsive to DNA damage. Science. 2007;316(5828):1160-1166.
48 44. Hsu PP, Kang SA, Rameseder J, et al. The mTOR-regulated phosphoproteome reveals a
mechanism of mTORC1-mediated inhibition of growth factor signaling. Science. 2011;332(6035):1317-1322.
45. Yu Y, Yoon SO, Poulogiannis G, et al. Phosphoproteomic analysis identifies Grb10 as an mTORC1 substrate that negatively regulates insulin signaling. Science. 2011;332(6035):1322-1326.
46. Hsieh AC, Liu Y, Edlind MP, et al. The translational landscape of mTOR signalling steers cancer initiation and metastasis. Nature. 2012;485(7396):55-61.
47. Valavanis C, Wang Z, Sun D, et al. Acheron, a novel member of the Lupus Antigen family, is induced during the programmed cell death of skeletal muscles in the moth Manduca sexta. Gene. 2007;393(1-2):101-109.
48. Glenn HL, Wang Z, Schwartz LM. Acheron, a Lupus antigen family member, regulates integrin expression, adhesion, and motility in differentiating myoblasts. American journal of physiology Cell physiology. 2010;298(1):C46-55.
49. Wang Z, Glenn H, Brown C, et al. Regulation of muscle differentiation and survival by Acheron. Mechanisms of development. 2009;126(8-9):700-709.
50. Shao R, Scully SJ, Jr., Yan W, et al. The novel lupus antigen related protein acheron enhances the development of human breast cancer. International journal of cancer. 2012;130(3):544-554.
51. Ji X, Lu H, Zhou Q, et al. LARP7 suppresses P-TEFb activity to inhibit breast cancer progression and metastasis. eLife. 2014;3:e02907.
52. Biewenga P, Buist MR, Moerland PD, et al. Gene expression in early stage cervical cancer. Gynecologic oncology. 2008;108(3):520-526.
53. Merret R, Martino L, Bousquet-Antonelli C, et al. The association of a La module with the PABP-interacting motif PAM2 is a recurrent evolutionary process that led to the neofunctionalization of La-related proteins. Rna. 2013;19(1):36-50.
54. Schaffler K, Schulz K, Hirmer A, et al. A stimulatory role for the La-related protein 4B in translation. Rna. 2010;16(8):1488-1499.
55. Yang R, Gaidamakov SA, Xie J, et al. La-related protein 4 binds poly(A), interacts with the poly(A)-binding protein MLLE domain via a variant PAM2w motif, and can promote mRNA stability. Molecular and cellular biology. 2011;31(3):542-556.
56. Albrecht M, Lengauer T. Survey on the PABC recognition motif PAM2. Biochemical and biophysical research communications. 2004;316(1):129-138.
57. Amrani N, Ghosh S, Mangus DA, et al. Translation factors promote the formation of two states of the closed-loop mRNP. Nature. 2008;453(7199):1276-1280.
58. Goss DJ, Kleiman FE. Poly(A) binding proteins: are they all created equal? Wiley interdisciplinary reviews RNA. 2013;4(2):167-179.
59. Adams DR, Ron D, Kiely PA. RACK1, A multifaceted scaffolding protein: Structure and function. Cell communication and signaling : CCS. 2011;9:22.
60. Yap TA, Carden CP, Kaye SB. Beyond chemotherapy: targeted therapies in ovarian cancer. Nature reviews Cancer. 2009;9(3):167-181.
61. Bai H, Li H, Li W, et al. The PI3K/AKT/mTOR pathway is a potential predictor of distinct invasive and migratory capacities in human ovarian cancer cell lines. Oncotarget. 2015;6(28):25520-25532. 62. Kitatani K, Usui T, Sriraman SK, et al. Ceramide limits phosphatidylinositol-3-kinase C2beta-controlled cell motility in ovarian cancer: potential of ceramide as a metastasis-suppressor lipid. Oncogene. 2016;35(21):2801-2812.
63. Dhillon AS, Hagan S, Rath O, et al. MAP kinase signalling pathways in cancer. Oncogene. 2007;26(22):3279-3290.
49 65. Lee YK, Park NH. Prognostic value and clinicopathological significance of p53 and PTEN in
epithelial ovarian cancers. Gynecologic oncology. 2009;112(3):475-480.
66. Ridley AJ. Rho GTPase signalling in cell migration. Curr Opin Cell Biol. 2015;36:103-112.
67. Schmitz AA, Govek EE, Bottner B, et al. Rho GTPases: signaling, migration, and invasion. Exp Cell Res. 2000;261(1):1-12.
68. Gyorffy B, Lanczky A, Szallasi Z. Implementing an online tool for genome-wide validation of survival-associated biomarkers in ovarian-cancer using microarray data from 1287 patients. Endocr Relat Cancer. 2012;19(2):197-208.
69. Zhu Y, Tian Y, Du J, et al. Dvl2-dependent activation of Daam1 and RhoA regulates Wnt5a-induced breast cancer cell migration. PloS one. 2012;7(5):e37823.
70. Pille JY, Denoyelle C, Varet J, et al. Anti-RhoA and anti-RhoC siRNAs inhibit the proliferation and invasiveness of MDA-MB-231 breast cancer cells in vitro and in vivo. Mol Ther. 2005;11(2):267-274. 71. Hwang YS, Hodge JC, Sivapurapu N, et al. Lysophosphatidic acid stimulates PC-3 prostate cancer cell Matrigel invasion through activation of RhoA and NF-kappaB activity. Mol Carcinog. 2006;45(7):518-529.
72. Vogelstein B, Papadopoulos N, Velculescu VE, et al. Cancer genome landscapes. Science. 2013;339(6127):1546-1558.
73. Faried A, Faried LS, Usman N, et al. Clinical and prognostic significance of RhoA and RhoC gene expression in esophageal squamous cell carcinoma. Ann Surg Oncol. 2007;14(12):3593-3601.
74. Fritz G, Brachetti C, Bahlmann F, et al. Rho GTPases in human breast tumours: expression and mutation analyses and correlation with clinical parameters. Br J Cancer. 2002;87(6):635-644.
75. Kamai T, Yamanishi T, Shirataki H, et al. Overexpression of RhoA, Rac1, and Cdc42 GTPases is associated with progression in testicular cancer. Clinical cancer research : an official journal of the American Association for Cancer Research. 2004;10(14):4799-4805.
76. Somlyo AV, Bradshaw D, Ramos S, et al. Rho-kinase inhibitor retards migration and in vivo dissemination of human prostate cancer cells. Biochemical and biophysical research communications. 2000;269(3):652-659.
77. Matsuoka T, Yashiro M. Rho/ROCK signaling in motility and metastasis of gastric cancer. World J Gastroenterol. 2014;20(38):13756-13766.
78. Chen S, Wang J, Gou WF, et al. The involvement of RhoA and Wnt-5a in the tumorigenesis and progression of ovarian epithelial carcinoma. Int J Mol Sci. 2013;14(12):24187-24199.
79. Horiuchi A, Imai T, Wang C, et al. Up-regulation of small GTPases, RhoA and RhoC, is associated with tumor progression in ovarian carcinoma. Laboratory investigation; a journal of technical methods and pathology. 2003;83(6):861-870.
80. Horiuchi A, Kikuchi N, Osada R, et al. Overexpression of RhoA enhances peritoneal dissemination: RhoA suppression with Lovastatin may be useful for ovarian cancer. Cancer Sci. 2008;99(12):2532-2539.
81. Wang X, Jiang W, Kang J, et al. Knockdown of RhoA expression alters ovarian cancer biological behavior in vitro and in nude mice. Oncol Rep. 2015;34(2):891-899.
82. Vega FM, Fruhwirth G, Ng T, et al. RhoA and RhoC have distinct roles in migration and invasion by acting through different targets. The Journal of cell biology. 2011;193(4):655-665.
51 A
B
52
Figure 1. Identification of LARP isoforms regulating lamellipodia formation.
(A) LARPs siRNAs efficiently decrease the expression of LARPs mRNA in SKOV3 cells. LARPs mRNA expression levels were determined by quantitative real-time PCR. Data shown (mean ± s.e., n = 3) are the mRNA values normalized to β- actin. * P <0.05, ** P < 0.005, *** P <0.001 (B) SKOV3 cells (2 × 104) grown on glass-bottom dishes were
transfected with 5 nM siRNAs for 48 h. Cells were fixed followed by staining with TRITC-conjugated phalloidin (red) and Hoechst 33342 (blue). Confocal microscopy representative images are shown and yellow arrows point to lamellipodia forming cell. The data showed (mean ± s.e., n = 4) are the percentages of cells forming lamellipodia. (C) Data shown (mean ± s.e., n = 4) are the percentages of cells forming lamellipodia, *** P < 0.0001. Three LARPs isoforms regulate lamellipodia formation in SKOV3 cells (LARP3, LARP4 and LARP4b).
53
Figure 2. LARP4 knockdown promotes lamellipodia formation in SKOV3 cells
(A) Protein expression demonstrated by western blot showing LARP4 knockdown effect of siRNA. (B) Cells were fixed followed by staining with TRITC-conjugated phalloidin (red) and Hoechst 33342 (blue). Confocal microscopy representative images are shown and yellow arrows point to lamellipodia. (C) The data showed (mean ± s.e., n = 11) are the percentages of cells forming lamellipodia. * P < 0.05, *** P < 0.0001.
C B
54
Figure 3. LARP4 overexpression decrease lamellipodia formation in SKOV3 cells
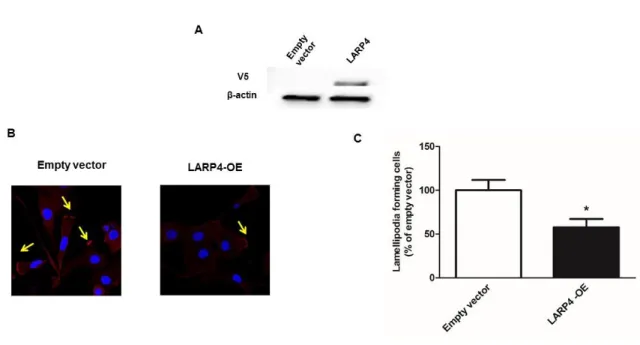
(A) Protein expression demonstrated by western blot showing LARP4 overexpression effect of LARP4 vector. (B) Cells were fixed followed by staining with TRITC-conjugated phalloidin (red) and Hoechst 33342 (blue). Confocal microscopy representative images are shown and yellow arrows point to lamellipodia. (C) The data showed (mean ± s.e., n = 4) are the percentages of cells forming lamellipodia. * P < 0.05, *** P < 0.0001.
55
Figure 4. RhoA knockdown decreases lamellipodia formation in SKOV3 cells
Representative data for the percentage of lamellipodia forming cells counted by confocal microscopy after cell fixation and staining with TRITC-conjugated phalloidin (red) and Hoechst 33342 (blue). The data shown (mean ± s.e., n = 11) are the percentages of cells forming lamellipodia. *** P < 0.0001.