107 魚類学雑誌 63(2):107–118 2016 年 11 月 5 日発行
本論文・Full Paper
1〒 501–1193 岐阜県岐阜市柳戸 1–1 岐阜大学地域科学部 2〒 514–8507 三重県津市栗真町屋町 1577 三重大学大学院生物資源学研究科 (2016 年 6 月 9 日受付;2016 年 8 月 25 日改訂;2016 年 8 月 29 日受理) キーワード:国内外来種 , 遺伝的撹乱 , カワヒガイ , ビワヒガイ , ミトコンドリア DNA Japanese Journal of Ichthyology © The Ichthyological Society of Japan 2016Miyu Suzuki, Shigeru Kitanishi*, Taiga Yodo and Takahiko Mukai. 2016. Population genetic structure of Sarcocheilichthys variegatus variegatus in the Tokai region, Japan and invasion of a non-indigenous mitochondrial DNA lineage. Japan. J. Ichthyol., 63(2): 107– 118.
Abstract Sarcocheilichthys variegatus variegatus, an endemic Japanese cyprinid fish
distributed from western Honshu (Tokai region) to Kyushu, has recently declined due to environmental damage and reduction of host mussels, and has been designated as a near threatened species in the Red List of the Japan Ministry of Environment. In addition, a recent study has noted the artificial introduction of S. v. microoculus from Lake Biwa into the Chubu, Kinki and Kyushu regions, suggesting a threat of genetic introgression. The present study, to investigate the population genetic structure of S. v. variegatus and identify invasions of a non-indigenous mtDNA lineage in the Tokai region, involved phylogeographic and population genetic analyses using cytochrome b gene sequences of 253 individuals of S. variegatus from Gifu, Mie and Shiga Prefectures. In addition, morphological characteristics of S. v. variegatus in the Tokai region, and S. v. microoculus in and around Lake Biwa, were used to identify instances of hybridization between the two subspecies. Phylogeographic analyses revealed greatest genetic divergence between populations of S. v. variegatus in the eastern and western regions of the Suzuka mountain range, and the presence of a non-indigenous mtDNA lineage in S. v. variegatus in the Tokai region. Clearly, the latter is characterized by important endemic genetic features, although already suffering from a loss of genetic identity due to the invasion of non-indigenous individuals. Steps are needed for future conservation of native S. v. variegatus populations in the Tokai region, including risk assessment and protection from invasive non-indigenous forms. *Corresponding author: Faculty of Regional Studies, Gifu University, 1–1 Yanagido, Gifu, 501–1193, Japan (e-mail: [email protected])
地
域集団の遺伝的構造や集団間の系統関係を 明らかにすることは,その集団の変遷過程 を明らかにするうえでも,また適切な保全策を立 案 す る う え で も 重 要 で あ る(Frankham et al., 2004).特に,純淡水魚は地域間での遺伝的分化 が進んでいることから,その保全のためには,各 地域固有の遺伝子系統の特定や集団の単位(たと え ば,Evolutionarily Significant Unit や Management Unit)の設定など,地域ごとの遺伝的特性に留意 した保護管理策の立案および実践が不可欠である(Moritz, 1994).一方,さまざまな国内の淡水魚類 において,異なる地域由来個体の人為移殖による遺 伝的撹乱が報告されており(たとえばメダカ種群 Oryzias latipes complex:Takehana et al., 2003;小山・ 北川,2009;ドジョウ Misgurnus anguillicaudatus:清 水・高木,2010;アブラボテ Tanakia limbata:松葉 ほか,2014),地域固有の遺伝子資源の消失が懸念 されている.しかし,このような国内外来種によ る遺伝的撹乱に対する知見は未だ十分ではなく, その実態把握は喫緊の課題となっている(瀬能,
東海地方におけるヒガイ属魚類の遺伝的集団構造と撹乱
鈴木美優
1・北西 滋
1・淀 太我
2・向井貴彦
12013). コ イ 科 カ ワ ヒ ガ イ Sarcocheilichthys variegatus variegatus は,愛知県豊川以西の濃尾平野から九 州北西部までの西日本地域に広く分布している日 本固有種であり,河川の中・下流域などの,水深 1–3 m の緩やかな流れのある環境に生息している (細谷,2001;細谷,2013).しかし近年,生息環 境の悪化や産卵に必要な二枚貝類の消失により, その生息数は減少し,環境省第 4 次レッドリスト において準絶滅危惧(NT)に選定されている(環 境省,2015).また,ミトコンドリア DNA(mtDNA) およびマイクロサテライト DNA を用いた先行研 究により,カワヒガイは鈴鹿山脈を境として東西 2 つの系統に分けられることが明らかとなってい る(Komiya et al., 2014).しかし,カワヒガイには, 琵琶湖および瀬田川に分布する亜種ビワヒガイ Sarcocheilichthys variegatus microocuhus が知られて おり(Hosoya, 1982),その資源増殖を目的とした 移殖放流が古くから日本各地で実施されてきた(中 村,1955;細谷,2001).ビワヒガイの人為的分 布拡大は,琵琶湖産アユ種苗の放流への混入の可 能性も考えられるが(瀬能,2013),いずれの要 因であったとしても,人為移殖されたビワヒガイ との交雑による各地域在来のカワヒガイの遺伝的 撹乱が危惧される(向井ほか,2014). カワヒガイの在来遺伝子資源を保全していくた めには,各地域在来のカワヒガイの詳細な遺伝的 集団構造の把握と,ビワヒガイ(および他地域産 カワヒガイ)の侵入・定着の現状把握,そして, それら非在来個体との交雑の有無などを明らかに することが不可欠である.特に,東海地方のカワ ヒガイ集団は,自然分布域の東端という地理的な 重要性に加え,系統的に滋賀県以西のビワヒガイ・ カワヒガイとは大きく異なることが示されており (Komiya et al., 2014),遺伝的固有性が非常に高い. さらに,東海地方においても,岐阜県には大規模 河川と濃尾平野が広がっている一方,三重県は比 較的小規模な河川のみという地勢的な相違が顕著 なことから,東海地方内部においても多様な遺伝 的構造が認められる可能性がある.これら地域固 有の遺伝的特性を詳細に把握し,その遺伝子資源 を保全することは,本種全体の遺伝的多様性保全 のためにも重要であると考えられる.そこで本研 究では,岐阜県および三重県のカワヒガイ集団を 対象とした, mtDNA のチトクローム b(cyt b)遺 伝子領域の塩基配列の分析により,東海地方にお けるカワヒガイの詳細な遺伝的集団構造の解明を おこなうとともに,ビワヒガイおよび他地域産カ ワヒガイの侵入について検討した.また,東海地 方における在来カワヒガイとビワヒガイの形態形 質の比較をおこない,交雑の可能性について検討 した. なお,滋賀県以西のビワヒガイとカワヒガイは mtDNA の み で は 必 ず し も 判 別 で き な い こ と (Komiya et al., 2014),調査地域において両亜種が 交雑している可能性があることから,本研究で採 集した個体は両亜種を区別せず “ ヒガイ属魚類 ” とし,非在来系統の mtDNA を持つ個体は “ 侵入 ヒガイ ” とした.また,西日本地域のヒガイ属(カ ワヒガイおよびビワヒガイ)のハプロタイプを “ 西 日本系統 ” とし,東海地方在来のヒガイ属(カワ ヒガイ)のハプロタイプを “ 東海系統 ” とする. 材 料 と 方 法 調査は東海地方の岐阜県,三重県の 26 地点, および比較に用いた滋賀県の 3 地点を対象とし, 2006–2008 年,2012 年,2014–2015 年 に か け て, 投網,たも網等を用いてヒガイ属魚類 253 個体を 採集した(Table 1, Fig. 1).カワヒガイの自然分 布域とされる愛知県では,近年ほとんどカワヒガ イが確認されておらず(荒尾,2015;梅村ほか, 2015),本研究では対象としなかった.採集個体 は,FA100(田村製薬株式会社)を用いて麻酔し た後, 展鰭し 10% 中性ホルマリンで固定した. 固定した標本は,デジタルノギス(スーパーキャ リパー CD67-S15PM,株式会社ミツトヨ)で各部 位の計測をおこなった後,70% エタノール中で保 存した.これらの標本のうち,岐阜県産標本は岐 阜県博物館,三重県産標本は三重大学大学院水産 実験所に登録,保管した(Appendix 1).また,各 個体の胸鰭,腹鰭,または体側筋の一部を DNA 解析用試料として 99.5% エタノール中に保存した. DNA 抽出は,エタノールで保存された試料の一 部から,QuickGene DNA Tissue Kit S(KURABO 社) を用いておこなった.mtDNA の cyt b 遺伝子領域 の PCR 増幅と,塩基配列の決定は,L14690-AH(5'-GGT CAT AAT TCT TGC TCG GA-3')と H15913-Thr-AH(5'-CCG ATC TTC GGA TTA CAA GAC CG-3')(梅 村ほか,2012)を用いておこなった.PCR には Ex Taq DNA Polymerase(タカラバイオ株式会社)を用 い, プ ロ ト コ ル に 従 っ て,95˚C1 分,55˚C1 分, 72˚C2 分 の 温 度 サ イ ク ル を 30 回 繰 り 返 し た. BigDye Terminator Cycle Sequencing Kit ver. 3.1
ヒガイ属魚類の遺伝的集団構造 109
(ThermoFisher Scientific 社)によるサイクルシーク エンス反応をおこなった後,磁気ビーズ Clean SEQ (Beckman Coulter 社 ) で 精 製 し,ABI3130 Genetic
Analyzer を用いて塩基配列の決定をおこなった. 得られた塩基配列は,CLUSTAL X(Tompson et al., 1997)を用いて多重整列をおこない,ハプロタイ プの決定後に,国際塩基配列データベース(INSD: International Nucleotide Sequence Database)に登録し た(登録番号:LC151414–LC151449). 系統樹の作成は 2 つのデータセットでおこなっ た.まず,日本産ヒガイ属全体における東海地方 のハプロタイプの位置付けを確認するため,本研 究 で 採 集 し た 253 個 体 と,Komiya et al.(2014) で報告された三重県から九州におけるハプロタイ プの塩基配列(581 bp)を用いて系統樹を作成し た(データセット 1).系統樹の作成には MEGA 6 (Tamura et al., 2013)を使用し,近隣結合法による 系統樹を作成した.遺伝距離のモデルは,内群の ハプロタイプ間に多重置換がほとんど無いことと, Komiya et al.(2014)のハプロタイプと本研究で見 出したハプロタイプの異同を図示することを目的 としたため,最もシンプルな p-distance を選択した. 外群には中国の Sarcocheilichthys kigngsiensis(登録 番号 EF193427;Zhang et al., 2008),Sarcocheilichthys p a r v u s ( E F19 3 4 5 9 ; Z h a n g e t a l . , 2 0 0 8 ), Sarcocheilichthys czerskii(EF193466;Zhang et al., 2008),Sarcocheilichthys nigripinnis(KF013979;Liu et al., 2013)を用いた.各枝の信頼性については 1,000 回のブートストラップ確率を求めた. 次に,東海地方におけるヒガイ属魚類の遺伝的 集団構造を把握するため,本研究で得られた 1135 bp の塩基配列(データセット 2)を用いた系統樹
Fig. 1. Maps showing sampling sites. Circle size reflects number of individuals collected at each site. Open and grey
circles represent haplotypes belonging to Tokai and Western clades of Sarcocheilichthys variegatus, respectively. Numbers correspond to those in Table 1.
作成を行った.また,比較 対 象として,三重県 (AB366542,LC098159,LC098160) お よび 滋 賀 県 (LC098163,LC098163,LC098164,LC098167– LC098169,LC098173)の既報データ(Kawamura et al., 2014;Tabata et al., 2016)も解析に加えた.系統 樹はデータセット 1 と同様の手法により作製した. また,東海地方で確認されたハプロタイプを対象 と し,Arlequin ver. 3.5(Excoffier and Lischer, 2010) および Hapster(Teacher and Griffiths, 2011)を用いて, minimum spanning network を作製した.
西日本系統のハプロタイプの侵入による,東海 地方における集団構造の撹乱の程度を推定するた め,本研究で得られた 253 個体の 1135 bp の塩基 配列(上述のデータセット 2)を用いて,分子分 散分析(AMOVA)をおこなった.AMOVA では, 標本を東海地方(岐阜県および三重県,232 個体) と 滋 賀 県(21 個体) の 2 つのグループに分け, 東海地方の西日本系統のハプロタイプを含むデー タ(データセット 2a,253 個体)と含まないデー タ(データセット 2b,221 個体)におけるグルー プ 間 の 遺 伝 的 分 化 を 求 め た.AMOVA に は Arlequin を用いた.さらに,東海地方におけるヒ ガイ属魚類における集団サイズの急速な拡大の有 無を検討するため,データセット 2b を用いて, mismatch distribution test(Rogers and Harpending, 1992),Tajimaʼs D(Tajima, 1989),Fuʼs Fs(Fu, 1997)を求めた.これらの解析では,岐阜県の集 団を,長良川(地点番号 1–12),揖斐川(地点番
Table 1. Sampling information and genetic diversity indices in Sarcocheilichthys populations. For site no., see Fig. 1
Site River name Locality N N’ Sampling year A A’ h h’ π π’
1 Nagara R. Gifu City, Gifu 44 6 2015 13 11 0.871 0.841 0.0079 0.0027 2 Ijira R. Nagara River system, Gifu City, Gifu 12 2 2012, 2015 7 6 0.909 0.889 0.0099 0.0035 3 Ishida R. Nagara River system, Gifu City, Gifu 12 0 2015 5 5 0.727 0.727 0.0019 0.0019 4 Toba R. Nagara River system, Gifu City, Gifu 13 0 2015 8 8 0.910 0.910 0.0031 0.0031 5 Toishi R. Nagara River system, Gifu City, Gifu 15 0 2015 10 10 0.933 0.933 0.0037 0.0037 6 Kawaura R. Nagara River system, Tomika, Gifu 2 0 2015 2 2 1.000 1.000 0.0053 0.0053 7 Seki R. Nagara River system, Seki, Gifu 10 0 2015 4 4 0.733 0.733 0.0029 0.0029 8 Goroku R. Nagara River system, Mizuho City, Gifu 9 0 2015 7 7 0.944 0.944 0.0038 0.0038
9 Sai R. Nagara River system, Mizuho, Gifu 1 0 2008 1 1 - - -
-10 Chougoji R. Nagara River system, Mizuho, Gifu 1 0 2008 1 1 - - - -11 Houe R. Nagara River system, Mizuho City, Gifu 6 0 2015 3 3 0.800 0.800 0.0033 0.0033 12 Kiso Sansen Park Nagara River system, Kaizu, Gifu 1 0 2006 1 1 - - - -13 Neo R. Ibi River system, Godo, Gifu 16 0 2015 8 8 0.858 0.858 0.0030 0.0030 14 Mimizu R. Ibi River system, Ono, Gifu 4 0 2015 4 4 1.000 1.000 0.0034 0.0034
15 Kudase R. Ibi River system, Ibigawa, Gifu 1 0 2015 1 1 - - -
-16 Oku R. Ibi River system, Ogaki, Gifu 9 0 2015 5 5 0.861 0.861 0.0028 0.0028 17 Doro R. Ibi River system, Tarui, Gifu 3 0 2015 3 3 1.000 1.000 0.0023 0.0023 18 Nakasu R. Ibi River system, Anpachi, Gifu 7 1 2015 6 5 0.952 0.933 0.0089 0.0027 19 Kawabe Dam Hida River, Kiso River system, Kawabe, Gifu 32 19 2015 7 2 0.730 0.462 0.0129 0.0020
20 Kiso R. Sakahogi, Gifu 2 0 2015 1 1 0 0 0 0
21 Shinsakai R. Kiso River system, Kakamigahara, Gifu 4 3 2015 3 1 0.833 - 0.0137 -22 Shirutani R. Miya River system, Ise, Mie 20 0 2014–2015 1 1 0 0 0 0
23 Oohori R. Meiwa, Mie 2 0 2014 1 1 0 0 0 0
24 Sasabue R. Meiwa, Mie 2 1 2015 2 1 1.000 - 0.0238
-25 Kongo R. Matsuzaka, Mie 1 0 2015 1 1 - - -
-26 Kumozu R. Tsu, Mie 3 0 2014–2015 2 2 0.667 0.667 0.0006 0.0006
27 Anonimous Creek Lake Biwa, Omihachiman, Shiga 17 17 2015 2 0 0.382 - 0.0040 -28 Nishinoko Lake Biwa, Omihachiman, Shiga 3 3 2015 2 0 0.667 - 0.0070
-29 Uso R. Hikone, Shiga 1 1 2007 1 0 - - -
-N: number of individuals; N': number of individuals possessed western haplotypes; A: number of haplotypes; A': number of haplotypes without western haplotypes; h: gene diversity; h': gene diversity without western haplotypes; π: nucleotide diversity; π': nucleotide diversity without western haplotypes
ヒガイ属魚類の遺伝的集団構造 111
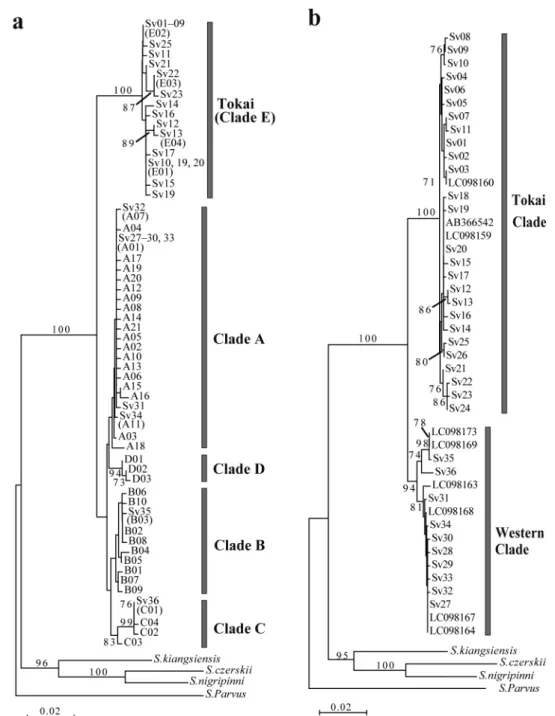
Fig. 2. a. Neighbor-joining (NJ) tree of Sarcocheilichthys variegatus variegatus, S. v. microoculus and S. biwaensis
based on partial mitochondrial cytochrome b (cyt b) sequences (581 bp) of 42 haplotypes (A01–21, B01–10, C01–04, D01–03 and E01–04) from Komiya et al. (2014), plus 36 haplotypes (Sv01–36) from 253 individuals of S. variegatus (this study). Short sequence haplotypes that could not be identified are represented by the same node (e.g., Sv01–09). Clades A–E correspond to Komiya et al. (2014). Two species and two subspecies of Japanese Sarcocheilichthys could not be identified from the mitochondrial phylogeny (Komiya et al., 2014). b. NJ tree based on mitochondrial cyt b sequences (1135 bp) of 36 haplotypes from 253 individuals of S. variegatus (this study), plus cited sequences from Kawamura et al. (2014) and Tabata et al. (2016). Numbers on nodes in both trees represent bootstrap probabilities (>70 %) of 1,000 pseudoreplicates.
号 13–18),木曽川(地点番号 19–21)の 3 水系に まとめ,また三重県 5 地点(地点番号 22–26)をま と め て 1 集 団 とし, 合 計 4 集 団 を 対 象 とし た. Mismatch distribution test では,spatial expansion model と sudden expansion model について解析し,モデル との差異は sum of square deviation を用いて判定し た.これらの解析には Arlequin を用いた.さらに, もし過去に個体群サイズの急速な拡大があったこ とが示唆された場合,その時期(t)を推定した. 推定には,t = τ/2u を用い,u の計算には u = 2 × 突然変異率×塩基配列長(1135 bp)を用いた(Rogers and Harpending, 1992; Schenekar and Weiss, 2011).ま た, 突 然 変 異 率 は 0.76%/Myr/ 系 統(Zardoya and Doadrio, 1999)を用いた. さらに,カワヒガイとビワヒガイの亜種判別形 質とされている尾柄高 / 頭長(カワヒガイ:≧ 49%,ビワヒガイ:< 49%;細谷,2013)を用いて, 亜種間の形態的差異と侵入ヒガイの形態的特徴を 求めた.解析では雌雄を区別せず,成魚 94 個体 (東海地方:88 個体,滋賀県:6 個体)を用いた. 成魚の基準は,中村(1969)に従って全長 8 cm 以上の個体とした.尾柄高および頭長の計測は, 中坊・中山(2013)に従い,標準体長に対する尾 柄高 / 頭長を求めた.解析では,東海地方におけ る在来系統のハプロタイプを持つ個体(72 個体) と東海地方で見出された西日本系統のハプロタイ プを持つ個体(侵入ヒガイ,16 個体)を区別し, また,比較のために,上記の滋賀県 6 個体に加え て琵琶湖博物館所蔵のビワヒガイ標本 81 個体 (Appendix 1)を用いた. 結 果 岐阜県,三重県,滋賀県から得られた 253 個体 のヒガイ属魚類の mtDNA の cyt b 領域 1,135 bp の 塩基配列を決定した結果, 36 ハプロタイプ(Sv01– 36)が検出された(Appendix 2).各地点における ハプロタイプ多様度(h)および塩基多様度(π)は, それぞれ,0–1.000,0–0.0137 であり,三重県およ び滋賀県の各地点と比べ,岐阜県の各地点は高い 遺伝的多様性を示す傾向にあった(Table 1).また, 西日本系統のハプロタイプ(Sv27–36)を除いたデー タセット 2b の場合,各地点のハプロタイプ多様 度(hʼ)および塩基多様度(πʼ)は,それぞれ, 0–1.000,0–0.0089 であり,塩基多様度の低下が認 められた(Table 1). データセット 1(581 bp)を用いた場合,上述 の 36 種類のハプロタイプは必ずしも区別されず,
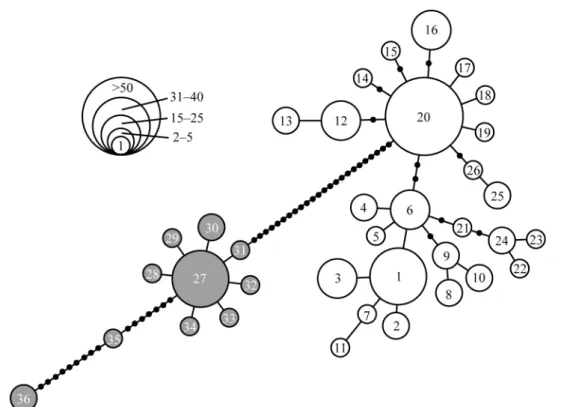
Fig. 3. Minimum spanning network of Sarcocheilichthys variegatus derived from cytochrome b
ヒガイ属魚類の遺伝的集団構造 113
20 種類のハプロタイプとなった.それらのデー タと Komiya et al. (2014)のデータを用いて作製 した近隣結合樹から,ヒガイ属のハプロタイプは 大きく 2 つのクレード群に分かれることが示され た[Clade A–D と Clade E, クレード名は Komiya et al.(2014)に従った;Fig. 2a].これらの間の塩 基 置 換 率 は 2.6–4.1%( 平 均 ± 標 準 偏 差:3.4 ± 0.3%)であった.本研究で採集した東海地方のヒ ガイ属からは,Clade D を除く,すべてのクレー ドのハプロタイプが認められた(Fig. 2a). 次に,データセット 2(1135 bp)を用いて近隣 結合樹を作製した結果,データセット 1 の Clade E に相当する東海系統(本研究の Sv01–26 ハプロ タイプおよび既報の三重県産標本からなるクレー ド)と,データセット 1 の Clade A–D に相当する 西日本系統(本研究の Sv27–36 ハプロタイプと既 報の琵琶湖産データからなるクレード)の 2 つに 分けられた(Fig. 2b).これらのクレード間の塩 基 置 換 率 は 2.2–2.8%( 平 均 ± 標 準 偏 差:2.5 ± 0.1%)であり,東海系統と西日本系統との間には 大きな遺伝的差異が存在していること明らかと なった.西日本系統のハプロタイプは,東海地方 では岐阜県の 5 地点および三重県 1 地点の合計 32 個体(13.8%)から認められた(Table 1,Fig. 1). 東海系統のハプロタイプは,さらに 2 つのサブク レードに分かれたが(それぞれ,Sv01–11,Sv12– 26),各ハプロタイプとその地理的分布との間に 明瞭な構造は認められなかった.しかし,岐阜県 と三重県の両地域におけるハプロタイプの分布パ ターンは大きく異なっており,優占するハプロタ イプ(Sv20)は両地域で共通しているものの,三 重県下では 2 種類の在来ハプロタイプ(Sv17 と Sv20)のみが見出され,また,殆どの個体(26/27 個 体 ) が 同 じ ハ プ ロ タ イ プ(Sv20) で あ っ た (Appendix 2).データセット 2 を用いてハプロタ イプネットワークを作製したところ,東海系統は Sv01,Sv06,Sv20 を中心とする灌木型の樹形を 示し,これら 3 つのハプロタイプは東海地方の河
Fig. 4. Mismatch distribution based on cytochrome b sequences of four Sarcocheilichthys variegatus
populations in the Tokai region. Solid and dotted lines represent the expected distribution under the spatial expansion model and 95% confidence intervals, respectively. Filled circles represent observed frequencies of pairwise differences.
川に広く分布していた(Fig. 3, Appendix 2).一方, 西日本系統は,Sv27 を中心とする花火型,およ びそれより大きく離れた 2 つのハプロタイプ(Sv35 と Sv36) からなっていた(Fig. 3).Sv27 は, 本 研究の琵琶湖水系産サンプルにおいても主要なハ プロタイプであり,東海地方においても広く認め られた(Appendix 2). 分子分散分析(AMOVA)をおこなった結果, 東海地方(岐阜県および三重県,232 個体)と滋 賀県(21 個体)の 2 つのグループ間に全遺伝分 散の 64.5%(FCT = 0.645, P < 0.001)が存在していた. 次に,東海地方における西日本系統のハプロタイ プを除いた場合,グループ間の遺伝分散は 86.6% (FCT = 0.866, P < 0.001)となり,西日本系統のハ プロタイプの侵入により,東海地方と西日本地域 との間の遺伝的差異が低下していることが示され た.
Mismatch distribution test の 結 果,4 集 団 す べ て において spatial expansion model の期待値の分布と の間に有意な差は無かった(長良川:τ = 3.523, SSD = 0.015, P = 0.425; 揖 斐 川:τ = 3.230, SSD = 0.004, P = 0.814;木曽川:τ = 5.529, SSD = 0.099, P = 0.276;三重県:τ = 0.054, SSD < 0.001, P = 0.249; Fig. 4).Sudden expansion model の場合は,木曽川 集団がモデルと適合しなかった(木曽川:τ< 0.001, SSD = 0.386, P < 0.001).また,長良川,揖斐川, 木曽川における Tajimaʼs D および Fuʼs Fs は,それ ぞれ,-0.983(P > 0.05)および -3.682(P > 0.05), -0.089(P > 0.05)および -1.697(P > 0.05),0.382(P > 0.05)および 3.828(P > 0.05)であり,岐阜県 内の河川はすべてゼロとの間に有意な差がなかっ た.しかし,三重県集団では,Tajimaʼs D および Fuʼs Fs は, そ れ ぞ れ -1.154(P > 0.05) お よ び -1.125(P < 0.05)であり,Fuʼs Fs はゼロから有意 に差があった.そこで,三重県集団では,過去の 集団サイズの急増が示されたため,その時期を推 定したところ,約 1,600 年前となった. 尾柄高 / 頭長の標準体長に対する割合を求めた 結果,侵入ヒガイは,東海系統と滋賀県産の中間 付近にプロットされる個体が多く,一部は東海系 統および滋賀県産の中にもプロットされた(Fig. 5). 考 察 東海地方におけるヒガイ属魚類の遺伝的特性 本研究では,日本産ヒガイ属魚類の mtDNA が東海 系統と西日本系統の 2 つに大別されることが示され た.同様のことは,先行研究によっても報告されて おり(Komiya et al., 2014),ヒガイ属魚類の遺伝的集 団構造として,鈴鹿山脈を境に,琵琶湖から九州ま でを含む西日本系統と,伊勢湾周辺の東海系統の 2 つの系統に分かれることが明らかにされている.鈴 鹿山脈を境とした遺伝的な分岐は,ミナミメダカ Oryzias latipes(Matsuda et al., 1997;Takehana et al., 2003)やカマツカ Pseudogobio esocinus(Tominaga et al., 2009, 2016),オイカワ Opsariichthys platypus(Kitanishi et al., 2016)など,他の多くの日本産淡水魚において も報告されており,ヒガイ属魚類の遺伝的分化にお いても,この山脈が非常に重要な地理的隔離要因で あったと考えられる.また,琵琶湖水系と同じ(ま たは類似した)ハプロタイプが,東海地方の河川で 不規則に分布している事例は,ゼゼラやシロヒレタ ビラ,カマツカで報告されているが,それらはいず れも人為移殖によるものと考えられている(堀川・ 向井,2007; Tominaga et al., 2009;梅村ほか,2012). したがって,東海地方のヒガイ属魚類における西日 本系統の mtDNA も,人為的攪乱に由来する可能性 が高いと考えられる. 東海地方のヒガイ属魚類において在来と考えら れる系統の中には 26 種類のハプロタイプが見出さ れ,また,ハプロタイプ間の遺伝的差異も大きい ことから,東海地方のヒガイ集団が,遺伝的多様 性を保持していることが示された.しかし,岐阜県 と三重県の両地域におけるハプロタイプの分布パ ターンは大きく異なっており,三重県下では殆どの
Fig. 5. Relationship between body length (mm) and
caudal-peduncle depth/head length (%) of Tokai region Sarcocheilichthys variegatus characterized by Tokai clade haplotypes (open circles), Tokai region S. variegatus characterized by Western clade non-indigenous haplotypes (grey squares), and Shiga Prefecture S. variegatus (crosses).
ヒガイ属魚類の遺伝的集団構造 115 個体(26/27 個体)が同じハプロタイプ(Sv20)であっ た.同様の分布パターンは Komiya et al.(2014)に おいても報告されており,1 種類のハプロタイプが 優占する低い遺伝的多様性は,三重県のヒガイ集 団の特性であると考えられる.一方,岐阜県下で は 25 種類のハプロタイプが認められ,また各地点 に固有のハプロタイプも複数認められたことから (Appendix 2),高い遺伝的多様性を持つことが示さ れた.こうした遺伝的構造をもたらした要因として, 氷期の古水系と,最終氷期後の集団サイズの変動 が考えられる.伊勢湾周辺地域では,海水面の低 下した氷期に伊勢湾周辺全域を集水域とする古水 系が存在したと考えられており(森山,2004),実 際にカワバタモロコ Hemigrammocypris rasborella や ウシモツゴ Pseudorasbora pugnax では,この古水系 を介した伊勢湾の東西における遺伝的交流が示唆 されている(Watanabe and Mori, 2008).東海地方の ヒガイ属においても,広域で優占的に分布する Sv20 のようなハプロタイプの存在は,伊勢湾周辺 地域における広範囲の遺伝的交流が生じていたこ とを示唆している.その後,最終氷期後の海面上 昇により,各地域の河川が分離することで,河川 規模が大きい木曽川水系(長良川,揖斐川を含む) では,遺伝的多様性を維持するのに十分な集団サ イズが維持されたが,三重県ではヒガイ属の生息 環境である河川中流域の面積が小さいことから, 最終氷期以降の縄文海進(約 6000 年前)などで集 団サイズの減少が著しく進み,遺伝的多様性が大 きく減 少したと考えられる.Mismatch distribution test の結果も,岐阜県の河川集団が比較的安定して いた一方,三重県集団においては縄文海進のよう な地質年代的に非常に新しい時代にボトルネック が生じたことを示唆している. 東海地方への西日本系統ヒガイの侵入 東海地 方から得られたヒガイ属 232 個体のうち, 岐阜県 の 5 地点 31 個体および三重県の 1 地点 1 個体が 西日本系統のハプロタイプを有しており(13.8%; 侵入ヒガイ),東海地方の複数の地点において西 日本からのヒガイ属魚類の侵入が生じていること が明らかとなった.このような東海地方への侵入 要因として,食用魚としての意図的な放流や,琵 琶 湖 産 ア ユ Plecoglossus altivelis altivelis 種 苗 へ の 混入,観賞魚の遺棄などが考えられる.食用魚と しての放流については,明治天皇が琵琶湖産のヒ ガイの味を,ことのほか好んだことから日本各地 に移殖されたことが知られているが(滋賀県立琵 琶湖文化館,1986),岐阜県揖斐郡においても, 江戸時代初期に揖斐川の治水を行った岡田将監が 琵琶湖から移入したとして「ショウゲンモロコ」 と呼ばれているとされている(揖斐郡教育会, 1993).揖斐郡において江戸時代に移殖したとす ることの真偽はともかく,在来のカワヒガイが分 布する岐阜県や三重県の河川において,食用魚と してのビワヒガイの移殖が過去に行なわれた可能 性は十分に考えられる. また,アユは,その水産業上の重要性から,毎 年日本各地で琵琶湖産種苗の放流が実施されてお り,多くの淡水魚でアユ種苗への混入による人為 的分布拡大が報告されている(瀬能,2013).本 研究で最も多くの侵入ヒガイが認められた川辺ダ ムを含む飛騨川流域では,2014 年に約 6,000 kg, 2015 年に約 5,500 kg の琵琶湖産アユ種苗の放流が 実施されている(飛騨川漁業協同組合ホームページ: http://hidagawa.com/).琵琶湖産アユ種苗への混入に よると考えられる琵琶湖産魚類の東海地方への侵入 は,さまざまな魚種について報告されており(ハス Opsariichthys uncirostris uncirostris:丹羽,1957;ゼゼ ラ Biwia zezera:堀川・向井,2007;シロヒレタビラ Acheilognathus tabira tabira:梅村ほか,2012;ビワヨ シノボリ Rhinogobius sp. BW:向井ほか,2015;オイ カワ Opsariichthys platypus:Kitanishi et al., 2016),東 海地方における侵入ヒガイの場合も,琵琶湖産アユ 種苗への混入による侵入が,十分に考えられる. こうした要因のほか,近年観賞魚として多くの 日本産淡水魚が広く販売・流通しており,そのよ うな観賞魚由来の淡水魚の侵入・定着もいくつか の 地 域 で 報 告 さ れ て い る た め( 向 井・ 西 田, 2003;小山・北川,2009;金尾,2013),ヒガイ 属魚類においても,同様に飼育魚の遺棄や意図的 放流の可能性が考えられる.データセット 1 にお いて侵入ヒガイが含まれたクレード A およびク レード B のハプロタイプは,近畿地方や中国地 方由来の個体の可能性もあるため(Komiya et al., 2014),本研究の結果からは観賞用のヒガイ属の 放流の可能性も排除できない. 母系遺伝するミトコンドリア DNA の解析では, 外来のミトコンドリア DNA を持つ侵入ヒガイが, 放流された移入個体そのものなのか,在来ヒガイ 属との交雑個体なのか,あるいは放流された移入 個体同士の子孫なのかを判別することはできない. しかし,いくつかの侵入ヒガイは,カワヒガイと 似た形態(尾柄高 / 標準体長が 49%以上)を有し ていた.さらに,僅かではあるが,東海系統の個 体の中に尾柄高 / 標準体長が 49%未満の個体が認
められた(5/72 個体).これらのことは,侵入ヒ ガイと在来の東海系統との間での交雑の可能性を 示している. 在来遺伝子系統の保全 東海地方のヒガイ属と 西日本のヒガイ属魚類との間の大きな遺伝的差異 (Komiya et al., 2014)を考えた場合,東海地方の ヒガイ属魚類の遺伝的な固有性は際立っている. しかし,本研究により,既に東海地方の自然水域 においても非在来のヒガイ属魚類が広く侵入して いること,そして,侵入ヒガイにより,西日本と 東海地方の集団間の遺伝的構造が不明瞭化してい ることが明らかとなった.さらに,侵入ヒガイと 在来個体との交雑の可能性も示された.これらの ことは,外来遺伝子の侵入や遺伝的撹乱により, 保護管理すべき単位として本来認識されるはずの 地域の在来系統や遺伝的特性が消失する可能性を 示している.愛知県においては,すでにカワヒガ イと同定される個体がほとんど確認されないため, 絶滅危惧 IA 類とされている(荒尾,2015).その ため,東海地方の在来ヒガイ属の保全を行うなら ば,今後,侵入リスクの高い地域の推定や外来ヒ ガイの侵入予防と防除などが必要と思われる. 謝 辞 標本の計測にご協力いただいた琵琶湖博物館の 金尾滋史博士に深く感謝する.また,試料の採集 および魚体の計測に協力していただいた岐阜大学 大学院連合農学研究科の伊藤 玄氏,試料の採集 に協力していただいた三重大学生物資源学部およ び大学院生物資源学研究科の小松史弥氏,田野太 志氏,中西 彬氏,野々目亮氏,一野哲志氏,田 中智士氏,髙島万莉子氏,三重県総合博物館の北 村淳一博士,琵琶湖博物館うおの会の田中治男 氏,川辺町生物多様性自然環境調査会の栗山圀彦 氏,田原治明氏,岐阜・美濃生態系研究会の松田 道一氏,岐阜市自然環境課自然係の番 雅義氏, 岐阜大学地域科学部の赤塚詠司氏および峯下千明 氏,DNA 解析にご協力いただいた岐阜大学生命 科学総合研究支援センターゲノム分野の皆様,標 本の登録と利用に便宜をはかっていただいた岐阜 県博物館の説田健一氏と三重大学大学院水産実験 所の木村清志教授に謝意を表したい.また,2 名 の査読者からは,多くの有益なご指導をいただい た.本研究の一部は,JSPS 科研費 26250044 の助 成によりおこなわれた.試料は適宜特別採捕許可 を取得して採集,もしくは当該河川の漁業協同組 合員が採集をおこなうなど,各調査地点において 合法な方法で採集された. 引 用 文 献 荒尾一樹.2015.カワヒガイ.愛知県環境調査セ ンター(編),p. 5.第三次レッドリスト レッ ドリストあいち 2015 汽水・淡水魚類 掲載種 の解説.愛知県環境部.
Excoffier, L. and H. E. L. Lischer. 2010. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour., 10: 564–567.
Frankham, R., J. D. Ballou, D. A. Briscoe and K. H. Mclnnes. 2004. A primer of conservation genetics. Cambridge University Press, Cambridge. xii+220 pp. Fu, Y.-X.. 1997. Statistical tests of neutrality of mutations
against population growth, hitchhiking and background selection. Genetics, 147: 915–925.
堀川まりな・向井貴彦.2007.濃尾平野における ゼゼラのミトコンドリア DNA 二型の分布.日本 生物地理学会会報,62: 29–34.
Hosoya, K. 1982. Classification of the cyprinid genus Sarcocheilichthys from Japan, with description of a new species. Japan. J. Ichthyol., 29: 127–138.
細谷和海.2001.ヒガイ属.川那部浩哉・水野信 彦(編),pp. 310–313.山渓カラー図鑑 日本の 淡水魚,第三版.山と渓谷社,東京. 細谷和海.2013.コイ科.中坊徹次(編),pp. 308– 327.日本産魚類検索全種の同定,第三版.東海 大学出版会,秦野. 揖斐郡教育会.1993.岐阜県揖斐郡 ふるさとの 自然.揖斐郡教育会,揖斐川.196 pp. 金尾滋史.2013.観賞魚店における日本産淡水魚 類の販売状況と課題.日本魚類学会自然保護委 員会(編),pp. 169–178.見えない脅威 “ 国内外 来種 ” どう守る地域の生物多様性.東海大学出 版会,秦野. 環 境 省.2015. 汽 水・ 淡 水 魚 の レ ッ ド リ ス ト. http://www.env.go.jp/press/press.php?serial=16264( 参 照 2016-5-9).
Kawamura, K., T. Ueda, R. Arai and C. Smith. 2014. Phylogenetic relationships of bitterling fishes (Teleostei: Cypriniformes: Acheilognathinae), inferred from mitochondrial cytochrome b sequences. Zool. Sci., 31: 321–329.
Kitanishi, S., A. Hayakawa, K. Takamura, J. Nakajima, Y. Kawaguchi, N. Onikura and T. Mukai. 2016. Phylogeography of Opsariichthys platypus in Japan based on mitochondrial DNA sequences. Ichthyol. Res., in press (DOI: 10.1007/s10228-016-0522-y).
Komiya, T., S. Fujita-Yanagibayashi and K. Watanabe. 2014. Multiple colonization of Lake Biwa by Sarcocheilichthys fishes and their population history.
ヒガイ属魚類の遺伝的集団構造 117 Environ. Biol. Fish., 97: 741–755.
小山直人・北川忠生.2009.奈良県大和川水系の メダカ集団から確認されたヒメダカ由来のミト コンドリア DNA.魚類学雑誌,56: 153–157. Lui, S. Q., Q. Y. Tang, X. J. Li and H. Z. Liu. 2013.
Plylogeographic analyses of Sarcocheilichthys nigripinnis (Teleostei: Cyrinidae) based on mitochondrial DNA cyt b gene sequences. Zool. Res., 34: 437–445.
松 葉 成 生・ 吉 見 翔 太 郎・ 井 上 幹 生・ 畑 啓 生. 2014.分子系統地理が示す愛媛県松山平野にお けるアブラボテの人為移入起源.魚類学雑誌, 61: 89–96.
Matsuda, M., H. Yonekawa, S. Hamaguchi and M. Sakaizumi. 1997. Geographic variation and diversity in the mitochondrial DNA of the medaka, Oryzias latipes, as determined by restriction endonuclease analysis. Zool. Sci., 14: 517–526.
Moritz, C. 1994. Defining ʻevolutionarily significant unitsʼ for conservation. Trends Ecol. Evol., 9: 373–375. 森山昭雄.2004.東海・三河湾の海底地形,とく に湾口部の海釜と砂堆地形.愛知教育大学研究 報告,53: 39–56. 向井貴彦・北原佳郎・森口宏明・酒井博嗣・浅香 智也・地村佳純.2015.西日本におけるビワヨシ ノボリ外来個体群の分布.日本生物地理学会会 報,70: 173–180. 向井貴彦・長野浩文・長野 光・宮島弘佳・千藤 克彦・説田健一.2014.岐阜県での分布が確認さ れたボウズハゼおよび証拠を伴う外来魚 5 種の 記録.岐阜県博物館調査研究報告,35: 1–9. 向井貴彦・西田 睦.2003.日本産ドンコにおけ るミトコンドリア DNA の系統と関東地方への人 為移植の分子的証拠.魚類学雑誌,50: 71–76. 中坊徹次・中山耕至.2013.魚類概説.中坊徹次 (編),pp. 3–30.日本産魚類検索全種の同定,第 三版.東海大学出版会,秦野. 中村守純.1955.関東平野に繁殖した移殖魚.日 本生物地理学会会報,16-19: 333–337. 中村守純.1969.ヒガイ.中村守純,pp. 149–156. 日本のコイ科魚類(日本産コイ科魚類の生活史 に関する研究).資源科学シリーズ 4,財団法人 資源科学研究所,東京. 丹羽 彌.1957.長良川の魚類.長良川の生物編 集委員会(編),pp. 190-215. 長良川の生物.岐阜 県.
R Core Team. 2014. R: A language and environment for statistical computing. Computing RFfS. http://www. R-project.org/
Rogers, A. R. and H. Harpending. 1992. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol., 9: 552–569.
Schenekar, T. and S. Weiss. 2011. High rate of calculation errors in mismatch distribution analysis results in numerous false inferences of biological importance.
Heredity, 107: 511–512. 瀬能 宏.2013.国内外来種とは何か.日本魚類 学会自然保護委員会(編),pp. 3–18.見えない脅 威 “ 国内外来種 ” どう守る地域の生物多様性.東 海大学出版会,秦野. 滋賀県立琵琶湖文化館.1986.湖国琵琶湖の魚た ち 改訂版.第一法規出版株式会社,東京.189 pp. 清水孝昭・高木基裕.2010.ミトコンドリア DNA による愛媛県を中心としたドジョウの遺伝的集 団構造と撹乱.魚類学雑誌,57: 13–26.
Tabata, R., R. Kakioka, K. Tominaga, T. Komiya and K. Watanabe. 2016. Phylogeny and historical demography of endemic fishes in Lake Biwa: the ancient lake as a promoter of evolution and diversification of freshwater fishes in western Japan. Ecol. Evol., 6: 2601–2623. Tajima, F. 1989. Statistical method for testing the natural
mutation hypothesis by DNA polymorphism. Genetics, 123: 585–595.
Takehana, Y., N. Nagai, M. Matsuda, K. Tsuchiya and M. Sakaizumi. 2003. Geographic variation and diversity of the cytochrome b gene in Japanese wild populations of Medaka, Oryzias latipes. Zool. Sci., 20: 1279–1291. Tamura, K., G. Stecher, D. Peterson, A. Filipski and S.
Kumar. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol., 30: 2725–2729. Teacher, A. G. F. and D. J. Griffiths. 2011. HapStar:
automated haplotype network layout and visualization. Mol. Ecol. Resour., 11: 151–153.
Tominaga, K., J. Nakajima and K. Watanabe. 2016. Cryptic divergence and phylogeography of the pike gudgeon Pseudogobio esocinus (Teleostei: Cyprinidae): a comprehansive case of freshwater phylogeography in Japan. Ichthyol. Res., 63: 79–93.
Tominaga, K., K. Watanabe, R. Kakioka, S. Mori and S. R. Jeon. 2009. Two highly divergent mitochondrial DNA lineages within Pseudogobio esocinus populations in central Honshu, Japan. Ichthyol. Res., 56: 195–199. Tompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin
and D. G. Higgins. 1997. The CLUSTAL X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res., 25: 4876–4882. 梅村啓太郎・二村 凌・高木雅紀・池谷幸樹・向 井貴彦.2012.岐阜県産シロヒレタビラにおける 外来ミトコンドリア DNA の分布.日本生物地理 学会会報,67: 169–174. 梅村錞二・酒井博嗣・内田良平・山本敏哉.2015. 阿摺ダム直下の淵のかいぼり調査結果.矢作川 研究,19:75–84.
Watanabe, K. and S. Mori. 2008. Comparison of genetic population structure between two cyprinids, Hemigrammocypris rasborella and Pseudorasbora pumila subsp., in the Ise Bay basin, central Honshu, Japan. Ichthyol. Res., 55: 309–320.
Zardoya, R. and I. Doadrio. 1999. Molecular evidence on the evolutionary and biogeographical patterns of European cyprinids. J. Mol. Evol., 49: 227–237. Zhang, L., Q. Y. Tang and H. Z. Liu. 2008. Phylogeny and
speciation of the eastern Asian cyprinid genus Sarcocheilichthys. J. Fish Biol., 72: 1122–1137.
Appendix 1. Museum voucher informations in this study.
Voucher specimens for mtDNA analyses of Sarcocheilichthys variegatus are deposited in the Gifu Prefectural Museum (GPM) and Fisheries Research Laboratory, Mie University (FRLM). The museum catalogue numbers are: GPM-Z 21852– 21896 and FRLM 52964–52985.
The specimens for morphological analyses were the same specimens in GPM, FRLM and museum collections in Lake Biwa Museum (LBM). The museum catalogue numbers of LBM are: 4003, 4051, 4057, 4062, 4831, 5818, 5828, 6018, 6044, 6150, 6192, 7257, 7450, 7922, 8130, 8234, 8569, 8976, 9232, 10224, 11132, 11279, 11285, 12448, 13282, 13313, 13519, 14361, 16162, 16934, 16957, 17070, 17169, 17224, 17643, 19459, 20392, 21362, 24801, 27118, 27643, 28826, 28940, 31202, 31482, 31491, 33271, 34280, 34901, 35093, 35401, 36255, 36593, 36601, 36914, 36948, 37654, 37663, 37762, 37942, 38506, 39330, 40530, 40532, 40534, 40536, 40538, 40540, 40923, 40945, 46924, 47307, 47309, 47319, 47337, 47338, 47339, 47340, 47342, 47345, 47346.
Appendix 2. Distribution of each haplotype for 29 sampling sites
Haplotype (Sv)
Site River name N 01 02 03 04 05 06 07 08 09 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36
1 Nagara R. 44 10 2 1 1 6 1 10 1 1 4 1 5 1 2 Ijira R. 12 2 2 1 1 3 1 2 3 Ishida R. 12 6 1 3 1 1 4 Toba R. 13 3 1 3 1 1 1 1 2 5 Toishi R. 15 3 1 1 1 3 1 1 1 2 1 6 Kawaura R. 2 1 1 7 Seki R. 10 2 5 2 1 8 Goroku R. 9 1 1 2 2 1 1 1 9 Sai R. 1 1 10 Chougoji R. 1 1 11 Houe R. 6 2 2 2
12 Kiso Sansen Park 1 1
13 Neo R. 16 1 2 5 1 4 1 1 1 14 Mimizu R. 4 1 1 1 1 15 Kudase R. 1 1 16 Oku R. 9 2 1 3 2 1 17 Doro R. 3 1 1 1 18 Nakasu R. 7 1 2 1 1 1 1 19 Kawabe Dam 32 4 9 14 2 1 1 1 20 Kiso R. 2 2 21 Shinsakai R. 4 1 2 1 22 Shirutani R. 20 20 23 Oohori R. 2 2 24 Sasabue R. 2 1 1 25 Kongo R. 1 1 26 Kumozu R. 3 1 2 27 Anonimous Creek 17 13 4 28 Nishinoko 3 2 1 29 Uso R. 1 1