九州大学学術情報リポジトリ
Kyushu University Institutional Repository
動的数理モデルを用いた抗体生産CHO細胞培養におけ るpH制御の最適化に関する研究
鳳桐, 智治
https://doi.org/10.15017/1931972
出版情報:Kyushu University, 2017, 博士(農学), 課程博士 バージョン:
権利関係:
全文
九州大学学術情報リポジトリ
Kyushu University Institutional Repository
動的数理モデルを用いた抗体生産CHO細胞培養におけ るpH制御の最適化に関する研究
鳳桐, 智治
https://doi.org/10.15017/1931972
出版情報:Kyushu University, 2017, 博士(農学), 課程博士 バージョン:
権利関係:
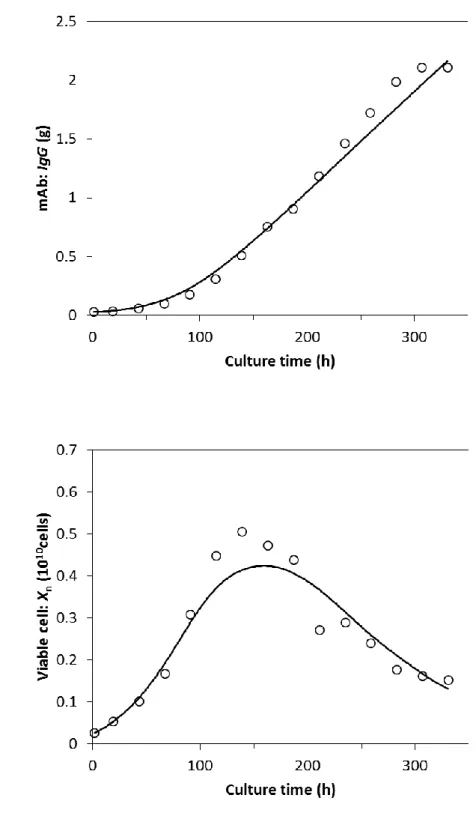
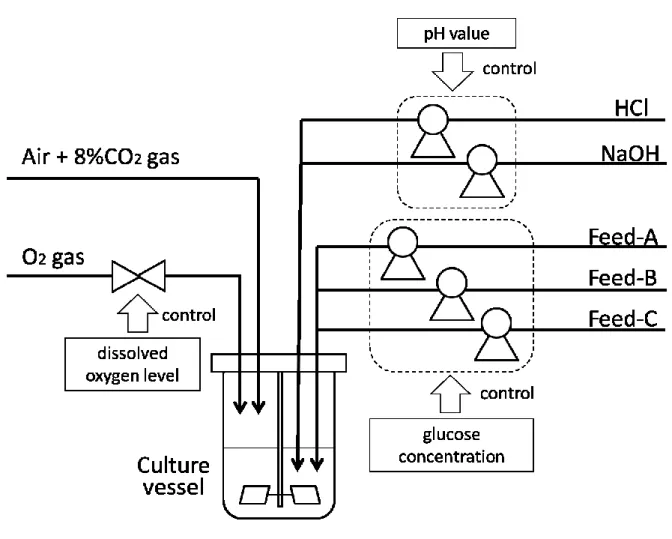
図



関連したドキュメント
For the thick case, this result was announced by Buekenhout, Delandtsheer, Doyen, Kleidman, Liebeck and Saxl, and in the thin case (where the lines have 2 points), it amounts to
In other words, the aggressive coarsening based on generalized aggregations is balanced by massive smoothing, and the resulting method is optimal in the following sense: for
It is suggested by our method that most of the quadratic algebras for all St¨ ackel equivalence classes of 3D second order quantum superintegrable systems on conformally flat
A generalization of Theorem 12.4.1 in [20] to the generalized eigenvalue problem for (A, M ) provides an upper bound for the approximation error of the smallest Ritz value in K k (x
In particular, we consider a reverse Lee decomposition for the deformation gra- dient and we choose an appropriate state space in which one of the variables, characterizing the
Atherosclerosis is a disease of the vasculature that is characterized by an accumulation of lipid-laden immune cells and apoptotic cells in the arterial wall.. Recently, the
In this paper, we focus on the existence and some properties of disease-free and endemic equilibrium points of a SVEIRS model subject to an eventual constant regular vaccination
(It is a standard convention to denote the unique line on two distinct collinear points x and y of a partial linear space by the symbol xy.) A linear space ðP ; LÞ with all lines