Some observations on flowering behaviors in strains of wild grass, Aegilops squarrosa, from different habitats of the South West Asia.
Mamoru SUGINO,
*
Kazuyohi KIMURA,**
Keizi MARUSHIGE***
7';L--;t:JJ.,.:Yr;J:DD-Y'/J.,.ttt;" l)tttc7)it:im.:JJ.,.:\" (/-<':/:JJ.,.:!f) AABBDD (7)!t1:.c7)lllJt;
(7)l--::>-c"<f:>~':'C1J{~Ghlv'~o 1955if*t~*~i]'7:J;L--J.,.· t.:/;(1~~1Jf.J~~ (~-&,
*
rn'::l:t]) 1J{1Thh, /-<'~A7':/, 77if::::"A7':/, 1'7:/tr:p,c.,t.:, :JJ.,.:¥'i:;J:tl~JU~fil!f&J(7)t*~
~1E1J{~~ht.:o~~G~;J:, *t~*~tt~~JF.!f, -'(-c7)-3 t;,(7)7';L--;t:JJ.,.:¥'~;b'fd\7tc7)flTc7)1:I!#f<t
-3 tt, Jt~~1t1:..fJ.~Jf5Et1T'?-c~t':1J{, ~@]~;J:, -'(-h G(7)rn":!¥±lBc, ~1tM'I'1c(7)~f*t.:--::>v,
-c;}(c7) J: -3 ~*fi*t ¥R-iS-L t.: 0
1) i5;i':'!J&.fJ.·
m
B!J&.fJ. (8' h B-&), -& B!J&.fJ. (-i!*'it7t) ~c"t':J:3T
~.:. hG~**7C(7)~1t&.r.tl;J:, Jt~B!J~ll:t':~h, ':'(7)':'C1J'G, :JJ.,.:¥(7)t.llJt;cL-cc7)tJ~1J{, +(7)~r.tB!J~~~t1
I.:i:;v'-c, ~1'ltGh~o
2) ~1t1JEi1tt 1t~5tll:(7)J2!~-3:.~~~(7)iJi;XY-C"I;J:1J' ~C, -~I':~t1J'G~",(7)*tJ3tB95t;fp C (7);tl3~1J{hGht.: o
3) 1:.W~ (growth habit) -c"h ~ Utn(7) l' 7:/, 77 if::::.. A 7' :/ (7) 1";L---fl;J: ~:JJ.,. :!f~,
~(7)/-<'~A 7' :/ (7) 1";L---7'I;J:*:J J.,. :¥'~t.:
h.z
~0 L 1J' L, 1~~';J:, ,16'1' L -c ~~B!J~t (7)-c"~
<,
15;1':' t ~<
-3 It ~ v' Cili B t.: J: '? -c ~1tiJ{1JE ~ h, -& B t.: J: '? -c~!fJ*1j ~ h ~ 04) .:. h G(7), B-& I.:
tiT
~ t#~ ~ &.r.t ,;J: , 1:.W±lB(7)~i':', ~%;7J<.ii(7)/ -<' 7' -:/c
B-&c
(7), ili v' 1:.W:ltNs't.:i:;tt ~ ~1*I:: J: '? -C, <f:> ~t¥.J3t~~Nl ~ h~ 0One of the key processes which determine the geographical distribution of plant species is the pattern of reproduction. Thus, for example, certain non-perenial plants are able to persist in the area by having the successful timing of flower initiation. followed by flower development and seed formation. On the other hand, since the day length is the most precise information of the seasonal change in nature for many plants and animals, sensitivity of plants to photo- period may be assumed to differ according to their lattitudinal distributions. Furthemore, the time of flowering is considerably modified by subjecting the plants to certain duration of cold.
The experiments described herein was conducted to examine the effect of cold treatment
*
Lab. of Plant Physiology, Dept. of Agriculture**
Inst. for Agricultural and Biological Sciences, Okayama University***
Lab. for Reproductive Biology, School of Medicine, University of N. C. U. S. A.and photo periods on flower initiation in some wild strains of Ae. squa"osa originated from Afganistan and its meighboring regions and to find possible relationships between their regional distribution and flowering behaviors.
Experimentals and Considerations
The materials used in the present experiments were seven strains ofAe. squarrosa collected originally at the various sites along the expeditional route in Pakistan, Afganistan and Iran by the Kyoto University Scientific Expedition to Karakoram indukush in 19552 (Fig.! and TableI). Those strains have been maintained at the esearch Institute of Agricultural Plant, Kyoto University and were offerred by the courtesy ofProf. Tananka for the present experiments.
40'
USSR
'---'---'--'200 - - - ' - ' 400km 39'38' N 37' 39'
- 38' 36'
35' 37'
KABUHL 36•
(AFGANISTAN
35'IRAN
34'
KANDAHAR 33'
, x2009 3 • .:. "'2001,2002,2003 2
QUETTA 31'
PAKISTAN
Fig. 1. Map showing the localities of original collection of the strains of Ae. squarrosa used in the present experiments. (cited from the report by Kihara et al) The figures in the map showed the strain number and the site of collection.
Table 1. The strain numbers coresponding to the original stock numbers, varieties and sites of collectin (1)
Strain (Symbols) Stock Original site
Variety
Nos. Nos. of collection
2001 (A) 2001-3 var. typica L. Quetta, PAKISTAN
2002 (B) 2001-6
2003 (C) 2003-4 var. anathem Ell,. "
2009 (D) 2017-2 Chaman,
2018 (E) 2040-2 var. typica L Ghazni, AFGANITAN
2089 (F) 2135-3 ssp. stmngulata ElG. Behshahr, I RAN 2100 (G) 2145-2 var. meyeri CRlSEB. Ramsar, IRAN
,. ',
39 The first experiment was started on March 1, 1960. In order to examine the effect of cold treatment on flowering, the germinating seed of each strains-were kept on wet sand in petri dishes at 2°to 3°C for 15 and 30 days in a refrigerator. The plants exposed to low temperature and non-chilling seedlings were planted on a medium of sand in a wooden box of 30x30xlO cm and cultured in the open air at Kyoto University, Kyoto. All the plants were grown under continuous light in which 100 watt incandescent lamps about 150 cm above the plant were used for supplemental illumination .during the night. To prevent tillering and simplify the observation, fertilizers were not applied to the plants throughout the experimental period.
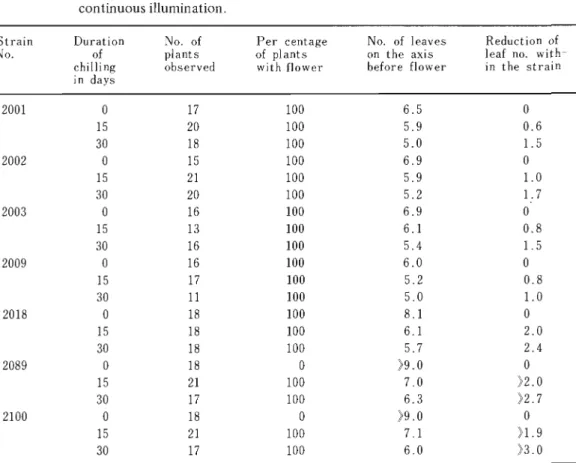
Two months after the start of culture, the flowering responses of the plants were examined and countered the number of leaves on the main axis before flower. Results are shown in Table 11.
Table 11. Flowering response of various strains of Ae. squarrosa subjected to chilling at 2-3°C for 15 and 30 days. After the chilling, plants were cultured under continuous illumination.
Strain Duration o. of Per centage No. of leaves Reduction of
No. of plants of plants on the axis leaf no. with-
chilling observed with flower before flower in the strain in days
2001 0 17 100 6.5 0
15 20 100 5.9 0.6
30 18 100 5.0 1.5
2002 0 15 100 6.9 0
15 21 100 5.9 1.0
30 20 100 5.2 1.7
2003 0 16 100 6.9 0
15 13 100 6.1 0.8
30 16 100 5.4 1.5
2009 0 16 100 6.0 0
15 17 100 5.2 0.8
30 11 100 5.0 1.0
2018 0 18 100 8.1 0
15 18 100 6.1 2.0
30 18 100 5.7 2.4
2089 0 18 0 )9.0 0
15 21 100 7.0 )2.0
30 17 100 6.3 )2.7
2100 0 18 0 )9.0 0
15 21 100 7.1 )1.9
30 17 100 6.0 )3.0
Five strains (2001,2002,2003,2009,2018) ofAe. squarrosa used in the present experiment flowered without chilling, but two (2089, 2100) of the northern strains did not. Acceleration of flowering by low temperature treatments were generally observed in all strains, and the number of leaves were decreased with increasing duration of chilling. With regard to the tendency of ealiness in flower initiation, which is indicated by the reduced number of leaves On axis before flower, the strains tested may be arranged as following order: 2009,2001,2002,2003, 2008,2089,2100
With somewhat exceptional case of the stra in 2009, the sequence is correlated to the latitudinal distribution of strains from south to north on. According to the degree of acceleration in
..
flowering by vernalization, these. strains may be divided into three groups. The first group includes the strains 2001, 2002, 2003, and 2009 in which the reduction of leaf number by chilling are relatively small. The second group includes .the strain 2018 which has more reduced number of leaves by chilling, and in the third group (strain 2089 and 2100) there are apparent difference in number of leaves between vernalized plants and non-chilling plants. As these groups also corespond to the three main habitats originally collected, they can be arbitrarily classified into Pakistanian, Afganistanian and Iranian strain, respectively.
The second experiment was conducted to examine the effect of photoperiods on flower initiation of these strains. In the first experiment, the plants were grown in the open air at Kyoto with relatively low temperature condition (Daily averages of the minimum and maximum temperature were 6.7-19.9°C in April and 12.0-24.0°C in May according to the meteologica1 data at Kyoto from 1931 to 1960). In the second experiment, plants were treated in the same way but subjected to the cold treatment for 30 days, after which they were grown in a green house with rougWy controlled temperature range of 20° to 28°C from July 29 to October 18.
Half of the plants were subjected to eight hours short photoperiods by exposing to day light from 9 A.M. to 5 P.M. every day. 'Long photoperiods (continuous light) were given to the remaining plants by supplementing illumination during the night as described in the first experiment. Observations were made 80 days after the start of the culture and the results are shown in Table Ill.
Table Ill. Flowering response of various strains of Ae. squarrosa subjected to short and long photoperiods following the chilling at 2_3°C for 30 days.
Strain Vernali- Photo- No. of Per cent age No. of leaves Reduction of No. zation period' plants of plants on the main leaf no. with-
observed with axis before in the strain flower flowering
2001 None Long 17 12 )11. 0 0
Short 17 100 7.6 3.4
Vern. Long 15 100 5.3 5.7
Short 18 100 7.4 3.6
2002 None Long 16 6 )11. 5 0
Short 17 100 7.3 )4.2
Vern. Long 16 100 5.4 )6.1
Short 16 100 7.6 )3.9
2003 None Long 8 0 )12.0 0
Short 10 100 9.5 )2.5
Vern. Long 15 100 5.5 )6.5
Short 16 100 7.1 )4.9
2009 None Long 8 100 9.8 0
Short 9 66 )10.5 -0.7
Vern. Long 7 100 5.1 4.7
Short 8 100 7.6 2.2
2018 None Long 17 0 )12.0 0
Short 16 0 )12.0
Vern. Long 14 100 6.4 )5.6
Short 15 80 10.2 )1.8
2089 None Long 16 0 )12.0 0
Short 17 12 ) 9.5 )2.5
Vern. Long 15 100 6.9 )5.1
Short 15 100 10.0 )2.0
• Long day: continuous light, Short day: eight- hour s photoperiod
.'
..
.,41
In non-vernalized plants, flower differentiation greatly w~re retarded and the plants initiated a larger number of leaves than did the plants in the first experiment, presumably ,owing to higher temperature conditions in a green house. The vegetative growth of the non-vernalized plants also was so accentuated that the effect ofvernalization on flower initiation was outstanding compared with that in the first experiment.
As shown in Table Ill, the Iranian strain, the Afganistanian strain and one of the Pakistanian strains did not flower at all without chilling. This indicates that the former two northern strains have a cold requirement for flower initiation and are classified into a typical winter type.
Although the Pakistarnian strains have been classified as a spring type, an acceleration of flowering occurred by short photoperiods if the plants were not exposed to low temperature with exception of the strain 2009 which seems to be a typical spring type.
It has been well established that Ae. squarrosa with genomes of DD is one of the ancestral wild species from which our present berad wheat with genomes of AABBDD has been derived 2
Since the strains used in the present experiment were originated from the localities within a limitted distributional range of Ae. squarrosa2 , they seem to be indigenous to the vast mountainous places in the South West Asia. Furthermore, from evolutional point of view, it is much likely that not only the differentiation of varieties but also many minor physiological differentiation adapted to diverse ecosystem have occurred in the species of Ae. squarrosa.
Recently, McMiIlan reported that the photoperiodic responses (critical photo periods) of American cocklebur population were diverse while those of wider spread Australian populations showed a uniform response to photoperiod, indicationg a indigenous distribution of the former Americans and newly introduction of the latter Australians.
The flowering behaviors of the strains of Ae. squarrosa tested in the present experiments showed a clear diversification in the three groups, corresponding to the site of collections in Pakistan, Afganistan and Iran. The earliness in flower irlitiation and the degree of acceleration by low temperature have a paralellism with their latitudinal distribution from south to north on.
The northern two groups are typical winter type, having a cold requirement for flower initiation.
The southern (Pakistanian) strains, except one strain (2009), are not the typical spring type, since flowering of them are suppressed by long days and accelerated by short days if the plants are not exposed to low temperature.
A short day induction of winter rye was first reported by Purvis et a14 , and later stated by the senior auther that spring rye and fully vernalized winter rye are typical long day plants while unvernalized winter rye is a short day plant in that its flowering is accelerated by limitted exposun" to short days5. In the case of some Pakistanian strains, however, flower initiation and further development to earing occurred in total short days without following long days.
Naturally, the flowering response of wild plants to photoperiod, especially in the long established endemic species, is thought to be manifold and also modified by local temperature conditions or water availability.It must be noted here that both promotive and inhibitory effect of photoperiod on the plant development are important for the normal growth and suvival of the concerned plants. From somewhat indirect meteological data shown in Table IV, a unique flowering response of the Pakistanian strains to short day may be explained as follows; Because of the severe dry season. (from May to November), the growing period of those wild strains in their native regions is restricted within the short duration from winter to spring season. Therefore .
.~
they (must) initiate flowers in shqrt days under low temperature conditions, and after being fulfy vernalized, further development of flower and the completion of their life cycle are hastened by longer photoperiods in the remaining short duration of spring season. But to make these relationship clear, more detailed experiments will be required.
Table IV. Monthly averages of temperature and precipitation for the 30 years (1931-1960) at Quetta, Kandahar, Kabuhl, Teheran and Kyoto6 . The average precipitation (in mm) was enclosed in parentheses.
Places Quetta Kandahar Kabuhl Teheran Kyoto
Latitude 30°10 N ~no' N 3nO' N 3nO' N 35° N
Altitude 1686 M 1038 m 1815 m 1220 m 43 m
Jan. 3.0 (52) 6.4 (79) -2.8 (30) 3.5 (35) 3.3 (51)
Feb. 6.0 (72) 9.5 (42) 0.6 (36) 5.2 (23) 3.8 (65)
Mar 9.0 (38) 13.9 (20) 6.4 (94) 10.2 (36) 6.9 (108)
Apr. 14.8 (22) 19.2 (8 ) 12.5 (102) 15.4 (31) 12.5 (141)
May 19.9 (12) 23.6 (5 ) 18.1 (20) 21. 2 (14) 17.6 (143)
June 24.4 (10) 27.0 (0 ) 22.0 ( 5 ) 26.1 ( 2) 21.7 (233)
July 26.6 (20) 28.9 ( 3) 24.7 ( 3) 29.5 ( 1 ) 26.0 (210)
Aug. 25.0 (4 ) 27.2 (0 ) 23.1 ( 3) 28.4 ( 1 ) 26.7 (160)
Sept. 19.8 (10) 22.0 (0 ) 20.0 (0 ) 24.6 ( 1 ) 22.0 (202)
Oct. 13.9 (0 ) 18.1 (0 ) 14.2 (15) 18.3 ( 5 ) 16.0 (124)
Nov. 8.9 ( 2 ) 12.5 ( 0 ) 8.6 (20) 10.6 (29) 10.9 (83)
Dec. 5.2 (28) 7.2 (20) 2.8 (10) 4.9 (27) 5.8 (55)
References
1) H.Kihara, K.Yamashita and M.Tanaka, Results of the Kyoto Universty Scientific Expedition to the Karakoram and Hindukush, 1955, Vol. 1,1965
2) H.Kihara, Cytologia 16, 1951.
3) C.McMillan, Sci. 171 (3975) : 1029-31, 1971.
4) O.N. Purvis and F.G.Gregory, Ann.Bot. N.S. 1: 569-592, 1937
5) O.N. Purvis, Handbuch der Pflanzenphysiologie, Bd.XVI , 76-117, 1961.
6) Tokyo Astronomical Observatory, Rika-Nenpyo (in Japanese), Maruzen, 1969.
"