Auxin-dependent compositional change in Mediator in ARF7- and ARF19-mediated transcription
39
0
0
全文
(2) Abstract Mediator is a multiprotein complex that integrates the signals from transcription factors binding to the promoter and transmits them to achieve gene transcription. The subunits of Mediator complex reside in four modules: the head, middle, tail, and dissociable CDK8 kinase module (CKM). The head, middle, and tail modules form the core Mediator complex, and the association of CKM can modify the function of Mediator in transcription. Here, we show genetic and biochemical evidence that CKM-associated Mediator transmits auxin-dependent transcriptional repression in lateral root (LR) formation. The AUXIN/INDOLE 3-ACETIC ACID 14 (Aux/IAA14) transcriptional repressor inhibits the transcriptional activity of its binding partners AUXIN RESPONSE FACTOR 7 (ARF7) and ARF19 by making a complex with the CKMassociated Mediator. In addition, TOPLESS (TPL), a transcriptional co-repressor, forms a bridge between IAA14 and the CKM component MED13 through the physical interaction. Chromatin immunoprecipitation assays show that auxin induces the dissociation of MED13, but not the tail module component MED25, from the ARF7 binding region upstream of its target gene. These findings indicate that auxin-induced degradation of IAA14 changes the module composition of Mediator interacting with ARF7 and ARF19 in the upstream region of their target genes involved in LR formation. We suggest that this regulation leads to a quick switch of signal transmission from ARFs to target gene expression in response to auxin. (219/250 words). 2.
(3) Significant Statement Mediator complex relays the information from transcription factors to RNA polymerase II. Our results show that Mediator transmits auxin-dependent transcription through its compositional change in lateral root formation. The AUXIN/INDOLE 3-ACETIC ACID 14 (Aux/IAA14) transcriptional repressor inhibits the transcriptional activity of its binding partner AUXIN RESPONSE FACTOR 7 (ARF7) and ARF19 by interacting with the dissociable CDK8 kinase module (CKM) of Mediator, a putative blocker of RNA polymerase II recruitment. Auxininduced degradation of IAA14 dissociates the CKM component, but not other Mediator subunit, from ARF7 binding to the upstream region of its target gene. We suggest that this compositional change in Mediator enables a quick switch of information transmission from ARFs to target gene expression in response to auxin. (119/120 words). 3.
(4) \body. Introduction Mediator is a multiprotein complex, which relays integrated information from transcriptional factors to RNA polymerase II (RNAPII) (1-3). Mediator consists of ~25 to 30 subunits, which are organized into three modules forming the core Mediator (head, middle and tail) and the dissociable CKM (3, 4). The Mediator structure, subunit organization and RNAPII interaction are conserved across eukaryotes, whereas Mediator is not a fixed complex. Structural analyses demonstrate that Mediator conformation and module organization are altered in accordance with the interaction with RNAPII (5, 6). RNAPII interacts with the middle module of the core Mediator, leading to a conformation change that could facilitate holoenzyme formation. CKM binds to the middle module, which interferes with the interaction of RNAPII with the core Mediator in vitro. In addition, recent accumulating evidence has demonstrated a wide range of Mediator functions involving in almost all stages of RNAPII transcription such as epigenetic regulation, transcriptional elongation, termination, mRNA processing, noncoding RNA activation and super enhancer formation (7, 8). In Arabidopsis, the core Mediator is composed of more than 21 subunits (9) and CKM consists of MED12/CRYPTIC PRECOCIOUS (CRP)/CENTER CITY (CCT), MED13/MACCHI-BOU2 (MAB2)/GRAND CENTRAL (GCT), CDK8/HUA ENHANCER3 (HEN3), and C-type cyclin (CycC) (10-13). Analysis of Arabidopsis Mediator subunits has shown that the subunits are important in regulating various developmental processes, phytohormone signaling pathways, developmental phase transition, and abiotic and biotic stress tolerance (14-16). Mutants of MED12/CRP/CCT and MED13/MAB2/GCT display embryonic development defects similar to those in auxin-related mutants and the genetic links between auxin-insensitive mutants, suggesting the involvement of Arabidopsis CKM in the auxin signaling pathway (10-12). The biological functions of the respective Mediator subunits are now emerging in plants; however, little is known about the molecular mechanism whereby Mediator integrates information from various transcription factors and transmits it to gene transcription. Auxin plays a crucial role in various aspects of physiological and developmental processes (17). To control these processes, auxin coordinates the transcription of numerous auxin-induced genes through actions of the auxin signaling module, the Aux/IAA transcriptional repressors and the ARF transcription factors (18, 19). The ARFs bind specific auxin response elements (AuxREs) in their target genes. The Aux/IAAs function as repressors of ARF-mediated transcription by forming multimers with ARFs (20, 21) and by recruiting the Groucho/Tup family co-repressor TPL and its family proteins (TPRs) (22, 23). Auxin induces the proteolysis of Aux/IAAs by the E3-ubiquitine ligase SCFTRANSPORT INHIBITOR RESPONSE 1/AUXIN SIGNALING F-BOX PROTEINS (TIR1/AFBs) complex to activate ARF-mediated transcription. However, the molecular mechanisms about how Mediator relays the information from the auxin signaling module ARFAux/IAA to general transcriptional machinery in response to auxin remain obscure. In this study, we describe the function of MAB2 and other Mediator subunits in LR initiation controlled by the auxin signaling module IAA14-ARF7-ARF19, and show the molecular mechanism of auxin responsive compositional changes in Mediator and its importance in their target gene transcription.. 4.
(5) Results and Discussion MAB2 Is Required for IAA14-dependent Repression of ARF7 and ARF19 Function To elucidate the molecular mechanism of the integration of signals from the Aux/IAA-ARF auxin signaling module and their transmission to target gene expression, we focus on the Mediator function in auxin-regulated LR formation. The LRs are initiated from the anticlinal cell division in the pericycle, a cell layer located deep within the primary root and attached to the outer side of the vascular bundle (24), and LRs grow through outer cell layers to appear on the surface of the root (Fig. S1A). Both ARF7 and ARF19 are required to activate the cell cycle in pericycle cells by inducing the expression of the target genes LATERAL ORGAN BOUNDARIES-DOMAIN16 (LBD16) and LBD29 (25, 26, 27, 28). The solitary-root-1 (slr-1) mutation in IAA14, which stabilizes the IAA14 protein (29), converts the ARF7 and ARF19 function from transcriptional activation to repression, leading to complete loss of LR formation (Fig. 1 A and B). In order to define the function of CKM in auxin signaling in LR formation, we examined genetic interaction between MAB2/MED13 and the Aux/IAA-ARF gene module. The mab2-4 single mutant exhibited a phenotype with shorter roots than wild type and showed a disordered LR distribution, but there was no difference in LR development and density between them (Fig. 1 A-C, and Fig. S1 A and B). The slr-1 mutant formed no LR initiation sites, whereas the mab2-4 slr-1 double mutant developed LRs showing a disorder distribution as the mab2 single mutant did (Fig. 1 A and B, and Fig. S1B). In addition, auxin treatment increased LR density in mab2-4 slr-1, but did not in slr-1 (Fig. S1C), demonstrating that the mab2 mutation restored the auxin responsiveness in the slr-1 background. The mab2-4 mutation also restored the expression of a direct target gene of ARF7 and ARF19 in LR formation, LBD16, in the slr-1 mutant backgrounds (Fig. 1 D-G). Similar restorations of LR formation with pericycle cell division and expression of ARF7 and ARF19 target genes in LR formation were found using transgenic plants expressing stabilized mIAA14-GFP protein under the control of the native IAA14 promoter and another mab2 allele, mab2-1 (Fig. 1 C and H, and Fig. S2A and S7A). In addition, the restoration was dissolved by complementation with pMAB2::MAB2-c-Myc (Fig. S3). Next, we examined the genetic relationship between ARF7 and ARF19, and MAB2 in LR formation. Like arf7 arf19 double mutants, the mab2-4 arf7-1 arf19-1 triple mutants had very few LRs (Fig. 1 A and B). The additional arf7-1 and arf19-1 mutations completely deleted LR formation in the mab2-4 slr-1 background (Fig. 1 A and B), indicating that the recovery of LR formation in slr-1 by mab2 depends on the ARF7 and ARF19 activity. In addition, to investigate whether mab2 affects the expression of IAA14, ARF7 and ARF19, we analyzed the expression of these genes in pIAA14::mIAA14-GFP-expressing Col and mab2-4. The expression of mIAA14GFP and ARFs were not affected by the mab2 mutations (Fig. S4). The mIAA14-GFP signal was also detected in the nuclei of the epidermal and stele cells in both genetic backgrounds (Fig. S2 B-G). These data indicate that MAB2 is involved in IAA14-dependent repression of ARF7 and ARF19 target genes. CKM-containing Mediator Is Involved in IAA14-dependent Repression To examine the involvement of other CKM subunits in IAA14-dependent repression of the ARF function in LR formation, we constructed crp/med12 slr-1 and hen3/cdk8 slr-1 double mutants. Like mab2 slr-1, crp slr-1 and hen3 slr-1 developed LR and showed the auxin sensitivity in LR formation, demonstrating that the crp and hen3 mutation restored the auxin responsiveness involved in LR formation in the slr-1 background (Fig. S5 A and B). These results indicate that CKM is involved in the IAA14-mediated block of LR formation. Furthermore, we investigated the function of the core Mediator in IAA14-mediated inhibition of LR formation. Mutations in MED17 and PHYTOCHROME AND FLOWERING TIME1 (PFT1)/MED25, which encode subunits of the head and tail Mediator module respectively (9), also rescued the LR-less phenotype in pIAA14::mIAA14-GFP expressing plants (Fig. S5C), indicating that not only CKM but also other Mediator modules are involved in IAA14-dependent LR-less phenotype. Our genetic data suggest that CKM-containing Mediator transmits a signal from IAA14 to repress the. 5.
(6) expression of ARF7 and ARF19 target genes. MAB2 Interacts with a Transcriptional Repressor Complex Including IAA14 and TPL To confirm the connection between IAA14 and CKM, we examined the physical interaction between IAA14 and MAB2. Although we observed no direct interaction between them in yeast (Fig. S6), we did observe interaction between stabilized mIAA14 and MAB2 by performing coimmunoprecipitation (Co-IP) assay using mab2-4 plants co-expressing MAB2-c-Myc and mIAA14-GFP (Fig. 2A). These results suggest that MAB2 indirectly interacts with IAA14. Recently, the co-repressor TPL/TPRs have been shown to mediate auxin-dependent transcriptional repression through direct interaction with Aux/IAA (23). In our study, we found that IAA14 interacted with TPL in yeast cells and plants (Fig. 2B and Fig. S6). We also found the tpl-1 mutation induced cell division in pericycle cells, resulting in LR formation in the presence of stabilized mIAA14-GFP as mab2 did (Fig. S7). In addition, the simultaneous mutation of TPL and MAB2 restored LR formation in plants expressing stabilized mIAA14 to the same extent as respective single mutations (Fig. S7 C and D). This indicates that TPL functions in the same pathway with MAB2 in IAA14-dependent repression of ARF7 and ARF19 target genes. Furthermore, we found direct interaction between MAB2 and TPL/TPR and the TPL linking between IAA14 and MAB2 in yeast and protoplast cells (Fig. 2 C-F). This is consistent with a previous report that HEN3, a component of CKM (12, 13), coexists with the transcription corepressor LEUNIG and the histone deacetylase HDA19 (30). However, we could not detect any interaction between MAB2 and TPL in plants co-expressing MAB2-c-Myc and TPL-HA under the normal growth condition (Fig. 2G). Only in the presence of the auxin antagonist auxinole, which stabilizes Aux/IAA by blocking TIR1-Aux/IAA interaction (31), did we find the stabilization of IAA14-GFP and the physical association with MAB2 and TPL in plants (Fig. 2G and Fig. S8), indicating that the interaction between MAB2 and TPL depends on Aux/IAA in plants. Our results suggest that the IAA14-TPL co-repressor complex recruits CKM to change the module composition of Mediator. IAA14 Recruits MAB2 to the Promoter Region of the Target Gene LBD16 To confirm IAA14-dependent recruitment of CKM to the upstream region of the target gene LBD16, the chromatin IP (ChIP) assays for MAB2 were performed in the presence of auxin, auxinole or mIAA14-GFP. We detected MAB2 binding weakly at the region I and II of LBD16 under the normal growth condition (Fig. 3A and B, and Fig. S9A). MAB2 dissociated from the 5’ upstream region of LBD16 in the presence of auxin (Fig. 3C and Fig. S9B). In contrast, in the presence of auxinole or mIAA14, MAB2 binding was strongly detected at the region I and II, weakly at the region III and IV close to TATA box upstream of the transcription start site of LBD16, where mIAA14 specifically bound at the region II containing AuxRE (Fig. 3 D and E, and Fig. S9 C and D). These results indicate IAA14-dependent MAB2 binding to the upstream region of LBD16. The region containing I to IV is shared by LBD16 and long non-coding RNA (lncRNA) At2g42425, whose expression was upregulated by auxin treatment and the mab2 mutation (Fig. 1H and Fig. S10). MAB2 binding at the region I and II under normal conditions suggests that these genes are ordinarily repressed by MAB2. MED25 Stably Binds to the Upstream Region of LBD16 with ARF7 We then investigated the connection of the core Mediator with the auxin-signaling module ARF7-ARF19-IAA14 in LR formation. We found the physical interaction of ARF7 and ARF19, but not of IAA14, with PFT1/MED25, the subunit of the tail Mediator module (9), and MED8 of the head module, but not with MED6 of the head module, in yeast (Fig. 4A and Fig. S11), and genetic interaction between ARF7 and MED8 (Fig. S12), whose mutation reportedly affects auxin response in LR formation (32). In addition, we observed interaction between stabilized mIAA14 and MED25 by performing Co-IP assay using mIAA14-GFP expressing plants (Fig. 4B). These results suggest that the core Mediator interacts with the auxin-signaling module. Next, to examine the behavior of the core Mediator in the upstream region of LBD16, ChIP. 6.
(7) assays for MED25 were performed using MED25-GFP expressing plants (33). MED25-GFP binding was detected at the region II containing AuxRE, where ARF7 specifically bound under normal conditions and in the presence of auxin (Fig. 4C and Fig. S9D). These results indicate that ARF7 is stably associated with AuxRE in the region II together with MED25 and that mIAA14, when bound to ARF7, recruits MAB2 to the upstream region of LBD16. Auxindependent degradation of IAA14 is suggested to cause the dissociation of MAB2 from the ARF7-containing transcriptional machinery. To confirm this, we performed Co-IP assay using arf7-1 arf19-1 mab2-4 co-expressing ARF7-GFP and MAB2-c-Myc. MAB2 and MED25 coexisted in the same protein complex as ARF7 in seedlings pretreated with auxinole (Fig. S13A). Subsequent exposure of these seedlings to auxin led to dissociation of MAB2, but not MED25, from the ARF7-containing protein complex, whereas subsequent exposure to auxinole caused no change in the interactions (Fig. S13A). We confirmed no change in the MAB2 and MED25 protein level with or without auxin (Fig. S13 B and C). These data indicate that auxindependent degradation of Aux/IAA dissociates MAB2 from ARF7-containing transcriptional machinery. Considering that all these factors are expressed in the pericycle attached to the outer side of the vascular bundle (Fig. S14) (25, 29), this event could be coupled to auxin-regulated LR formation. In conclusion, we have shown that CKM-associated Mediator transmits repressive signals from the ARFs-IAA14-TPL complex to target gene expression, resulting in inhibited LR formation (Fig. 4Di). Furthermore, auxin-induced proteolysis of IAA14 leads to the dissociation of CKM, but not the core Mediator, from ARF7 and ARF19 (Fig. 4Dii). As a result, the active form of ARFs induces the expression of target genes via the core Mediator (Fig. 4Diii). Considering that single mutations in the core Mediator subunits suppress mIAA14-dependent LR-less phenotype (Fig. S5C), the mutations might affect the interaction between CKM and the core Mediator, without much effect on the function of the core Mediator in auxin signaling (Fig. S12), leading to the dissociation of CKM from the core Mediator. If one considers that CKM structurally prevents RNAPII interaction with the core Mediator (5, 6), it follows that the IAA14-TPL repressor complex may inhibit the recruitment of RNAPII to target genes by associating CKM with the core Mediator, whereas CKM-dissociated Mediator may recruit RNAPII in the presence of auxin. This regulation can enable a quick transcriptional switch between activation and repression in response to auxin. Meanwhile, TPL controls the chromatin remodeling with histone deacetylases to repress transcription (22, 34, 35). Both repression mechanisms, CKM-dependent RNAPII exclusion and TPL-dependent chromatin remodeling, seem to be necessary to achieve auxin-dependent transcriptional repression because both the mab2 and tpl-1 mutations partially rescued slr-1 phenotypes respectively (Fig. 1 and Fig. S7). Thus, our findings provide novel insights into the role of TPL in repressing auxin signaling and propose a model for rapid auxin-dependent gene activation.. 7.
(8) Materials and Methods Plant materials and growth conditions. Arabidopsis thaliana accessions Columbia (Col-0) and Landsberg erecta (Ler) were used as wild types. Descriptions of mab2-1, mab2-4, arf7-1, arf191, slr-1, crp-4, hen3-1, med17, pft1-2, med8, pIAA14::IAA14-GFP, pIAA14::mIAA14-GFP, pLBD16::GUS and p35S::MED25-GFP lines have been made previously (11, 12, 25, 26, 33, 36). The tpl-1 and pTPL::TPL-HA lines we used were kindly provided by Jeff Long (University of California, Los Angels, USA), while hen3-1 mutant was obtained from Arabidopsis Biological Resource Center (37). Seeds were surface-sterilized, plated on Murashige and Skoog medium plates, and germinated as described previously (38). Plants were transferred to soil and grown at 23˚C under constant light as described previously (38). Transgenic plants. In order to construct pARF7::ARF7-GFP, ARF7 cDNA was subcloned into the pDONR201 vector (Invitrogen) and transferred to the pGWB-GFP(C) vector under a 2.5 kb ARF7 promoter (The pGWB-GFP(C) vector was constructed by inserting the GFP coding region into the Xba I/Sac I sites of the pGWB1 vector (39)). We transformed pARF7::ARF7-GFP into LR-less arf7-1 arf19-1 plants via floral dipping using Agrobacterium tumefaciens (strain MP90) (40). Four kanamycin-resistant transformants developed LR, but less than wild type, indicating that pARF7::ARF7-GFP partially complemented the double mutant phenotype (25). To generate pMAB2::MAB2-c-Myc, MAB2 cDNA were subcloned into the pDONR221 vector (Invitrogen) and transferred to the pGWB16 vector (39) under a 2.0 kb MAB2 promoter. We transformed pMAB2::MAB2-c-Myc into heterozygous mab2-4 plants via floral dipping using Agrobacterium tumefaciens (strain MP90) (40). Thirteen kanamycin-resistant transformants homozygous for the mab2-4 mutation displayed normal cotyledon development and fertility, indicating that pMAB2::MAB2-c-Myc complemented the mab2 phenotype (12). The transgene was introduced into mab2-4 pIAA14::mIAA14-GFP, tpl-1 pTPL::TPL-HA and arf7 arf19 pARF7::ARF7-GFP plants by crossing to construct the mab2-4 pMAB2::MAB2-c-Myc pIAA14::mIAA14-GFP, tpl-1 mab2-4 pTPL::TPL-HA pMAB2::MAB2-c-Myc, and arf7-1 arf19-1 mab2-4 pARF7::ARF7-GFP pMAB2::MAB2-c-Myc plants, respectively. Yeast two-hybrid (Y2H) and Y3H assays. The Y2H assay was performed as described previously (12). Empty vectors were used as negative controls and pBD-p53 and pAD-T7 were used as positive controls. In the Y3H assay, full-length TPL and TPL without CTLH domain (TPLΔCTLH) cDNAs were subcloned into p427-TEF. The plasmids were introduced into the AH109 strain together with pAD-GAL4-MAB2 and pBD-GAL4-IAA14. We selected p427TEF-introduced yeasts on the basis of their resistance to G418 (200 µg/l; Wako). The empty vector p427-TEF was used as negative controls. Interaction was detected by the expression of the HIS3 reporter gene. Co-IP and immunoblot. For Co-IP assays, root parts of 7-d-old seedlings were homogenized in a protein extraction buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1% NP40, 0.6 mM phenylmethyl sulphonyl fluoride, and Roche protease inhibitor cocktail). We conducted IP from protein extracts using the µMACS GFP-tagged protein isolation kit and the µMACS c-myctagged protein isolation kit (Miltenyi Biotec), according to the manufacturer's instructions. Immunoblot assays were performed using anti-GFP (Clontech JL-8), anti-HA (Roche 12CA5), anti-c-Myc (Santa Cruz Biotechnology 9E10) and anti-MED25 antibodies (33). The bands of interest were identified by proper molecular-weight bands of Precision Plus ProteinTM Dual Color Standards (Bio-Rad). ChIP assay. The ChIP assays were performed according to the previously-reported protocol (41) with slight modification. 2 or 4 g of 7 d-old seedlings were used as samples. In nuclei isolation buffer, Roche protease inhibitor cocktail was used in place of pepstatin A and aprotinin. After preclearing with salmon sperm DNA-protein G agarose beads (Millipore), cross-linked chromatin from seedlings was immunoprecipitated with anti-GFP antibody (Sigma-Aldrich. 8.
(9) GFP-20), anti-Myc antibody (Abcam 9E10) or normal mouse IgG (Santa Cruz Biotechnology). The ratio between the bound DNA after IP and the input DNA before IP was calculated for all the representative primer sets spanning the LBD16 genomic region, shown in the Supplementary Table.. 9.
(10) References 1. Malik S, Roeder RG (2005) Dynamic regulation of pol II transcription by the mammalian Mediator complex. Trends Biochem. Sci 30(5): 256-263. 2. Yin JW, Wang G (2014) The Mediator complex: a master coordinator of transcription and cell lineage development. Development 141(5): 977-987. 3. Carlsten JO, Zhu X, Gustafsson CM (2013) The multitalented Mediator complex. Trends Biochem Sci 38(11): 531-537. 4. Tsai KL, et al. (2014) Subunit architecture and functional modular rearrangements of the transcriptional mediator complex. Cell 157(6): 1430-1444. 5. Elmlund H, et al. (2006) The cyclin-dependent kinase 8 module sterically blocks Mediator interactions with RNA polymerase II. Proc Natl Acad Sci U S A 103(43): 15788-15793. 6. Tsai KL, et al. (2013) A conserved Mediator-CDK8 kinase module association regulates Mediator-RNA polymerase II interaction. Nat Struct Mol Biol 20(5): 611-619. 7. Allen BL, Taatjes DJ (2015) The Mediator complex: a central integrator of transcription. Nat Rev Mol Cell Biol 16(3): 155-166. 8. Carlsten JO, Zhu X, Gustafsson CM (2013) The multitalented Mediator complex. Trends Biochem Sci 38(11): 531-537. 9. Bäckström S, Elfving N, Nilsson R, Wingsle G, Björklund S (2007) Purification of a plant mediator from Arabidopsis thaliana identifies PFT1 as the Med25 subunit. Mol Cell 26(5): 717-729. 10. Gillmor CS, et al. (2010) The MED12-MED13 module of Mediator regulates the timing of embryo patterning in Arabidopsis. Development 137(1): 113-122. 11. Imura Y, et al. (2012) CRYPTIC PRECOCIOUS/MED12 is a novel flowering regulator with multiple target steps in Arabidopsis. Plant Cell Physiol 53(2): 287-303. 12. Ito J, Sono T, Tasaka M, Furutani M (2011) MACCHI-BOU 2 is required for early embryo patterning and cotyledon organogenesis in Arabidopsis. Plant Cell Physiol 52(3): 539-552. 13. Wang W, Chen X (2004) HUA ENHANCER3 reveals a role for a cyclin-dependent protein kinase in the specification of floral organ identity in Arabidopsis. Development 131(13): 3147-3156. 14. An C, Mou Z (2013) The function of the Mediator complex in plant immunity. Plant Signal Behav 8(3): e23182. 15. Kidd BN, Cahill DM, Manners JM, Schenk PM, Kazan K (2011) Diverse roles of the Mediator complex in plants. Semin Cell Dev Biol 22(7): 741-748. 16. Samanta S, Thakur JK (2015) Importance of Mediator complex in the regulation and integration of diverse signaling pathways in plants. Front Plant Sci 17(6): 757. 17. Vanneste S, Friml J (2009) Auxin: a trigger for change in plant development. Cell 136(6):1005-1016. 18. Salehin M, Bagchi R, Estelle M (2015) SCFTIR1/AFB-Based Auxin Perception: Mechanism and Role in Plant Growth and Development. Plant Cell 27(1):9-19. 19. Santner A, Calderon-Villalobos LI, Estelle M (2009) Plant hormones are versatile chemical regulators of plant growth. Nat Chem Biol 5(5): 301-307. 20. Korasick DA, et al. (2014) Molecular basis for AUXIN RESPONSE FACTOR protein interaction and the control of auxin response repression. Proc Natl Acad Sci USA 111(14): 5427-5432. 21. Nanao MH, et al. (2014) Structural basis for oligomerization of auxin transcriptional regulators. Nat Commun 7(5): 3617. 22. Long JA, Ohno C, Smith ZR, Meyerowitz EM (2006) TOPLESS regulates apical embryonic fate in Arabidopsis. Science 312(5779): 1520-1523. 23. Szemenyei H, Hannon M, Long JA (2008) TOPLESS mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis. Science 319(5868): 13841386. 24. Lavenus J, et al. (2013) Lateral root development in Arabidopsis: fifty shades of auxin. Trends Plant Sci 18(8): 450-458.. 10.
(11) 25. Okushima Y, et al. (2005) Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: unique and overlapping functions of ARF7 and ARF19. Plant Cell 17(2): 444-463. 26. Okushima Y, Fukaki H, Onoda M, Theologis A, Tasaka M (2007) ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 19(1): 118-130. 27. Iwakawa H, et al. (2002) The ASYMMETRIC LEAVES2 gene of Arabidopsis thaliana, required for formation of a symmetric flat leaf lamina, encodes a member of a novel family of proteins characterized by cysteine repeats and a leucine zipper. Plant Cell Physiol 43(5): 467-478. 28. Shuai B, Reynaga-Pena CG, Springer PS (2002) The lateral organ boundaries gene defines a novel, plant-specific gene family. Plant Physiol 129(2): 747-761. 29. Fukaki H, Tameda S, Masuda H, Tasaka M (2002) Lateral root formation is blocked by a gain-of-function mutation in the SOLITARY-ROOT/IAA14 gene of Arabidopsis. Plant J 29(2): 153-168. 30. Gonzalez D, Bowen AJ, Carroll TS, Conlan RS (2007) The transcription corepressor LEUNIG interacts with the histone deacetylase HDA19 and mediator components MED14 (SWP) and CDK8 (HEN3) to repress transcription. Mol Cell Biol 27(15): 5306-5315. 31. Hayashi K, et al. (2012) Rational design of an auxin antagonist of the SCFTIR1 auxin receptor complex. ACS Chem Biol 7(3): 590-598. 32. Raya-González J, Ortiz-Castro R, Ruíz-Herrera LF, Kazan K, López-Bucio J (2014) PHYTOCHROME AND FLOWERING TIME1/MEDIATOR25 Regulates Lateral Root Formation via Auxin Signaling in Arabidopsis. Plant Physiol 165(2): 880-894. 33. Chen R, et al. (2012) The Arabidopsis mediator subunit MED25 differentially regulates jasmonate and abscisic acid signaling through interacting with the MYC2 and ABI5 transcription factors. Plant Cell 24(7): 2898-2916. 34. Krogan NT, Hogan K, Long JA (2012) APETALA2 negatively regulates multiple floral organ identity genes in Arabidopsis by recruiting the co-repressor TOPLESS and the histone deacetylase HDA19. Development 139(22): 4180-4190. 35. Wang L, Kim J, Somers DE (2013) Transcriptional corepressor TOPLESS complexes with pseudoresponse regulator proteins and histone deacetylases to regulate circadian transcription. Proc Natl Acad Sci U S A 110(2): 761-766. 36. Xu R, Li Y (2012) The Mediator complex subunit 8 regulates organ size in Arabidopsis thaliana. Plant Signal Behav 7(2): 182-183. 37. Alonso JM, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301(5633): 653-657. 38. Fukaki H, Fujisawa H, Tasaka M (1996) Gravitropic response of inflorescence stems in Arabidopsis thaliana. Plant Physiol 110(3): 933-943. 39. Nakagawa T, et al. (2007) Development of series of gateway binary vectors, pGWBs, for realizing efficient construction of fusion genes for plant transformation. J Biosci Bioeng 104(1): 34-41. 40. Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16(6): 735-743. 41. Saleh A, Alvarez-Venegas R, Avramova Z. (2008) An efficient chromatin immunoprecipitation (ChIP) protocol for studying histone modifications in Arabidopsis plants. Nature Protoc 3(6): 1018-1025. 42. Paciorek T, Sauer M, Balla J, Wisniewska J, Friml J (2006) Immunocytochemical technique for protein localization in sections of plant tissues. Nat protoc 1(1): 104-107. 43. Malamy JE, Benfey PN (1997) Organization and cell differentiation in lateral roots of Arabidopsis thaliana. Development 124(1): 33-44. 44. Taoka K, et al. (2011) 14-3-3 proteins act as intracellular receptors for rice Hd3a florigen. Nature 476(7360): 332-335.. 11.
(12) 45. Robatzek S, Somssich IE (2001) A new member of the Arabidopsis WRKY transcription factor family, AtWRKY6, is associated with both senescence- and defence-related processes. Plant J 28(2): 123-133. 46. Takeuchi M, Ueda T, Sato K, Abe H, Nagata T, Nakano A (2000) A dominant negative mutant of Sar1 GTPase inhibits protein transport from the endoplasmic reticulum to the Golgi apparatus in tobacco and Arabidopsis cultured cells. Plant J 23(4): 517-525.. 12.
(13) Acknowledgments We thank Jeff Long for providing tpl-1 and pTPL::TPL-HA lines, Takashi Araki for providing crp-4, Taku Demura for providing pAD-GWRFC and pBD-GWRFC vectors and Kenichiro Hayashi for providing auxinole. This work was partly supported by the Ministry of Education, Culture, Sports, Science and Technology (MEXT) through Grants-in-Aid for Scientific Research on Priority Areas (14036222 and 19060007) (to M.T.), Grant-in-Aid for Young Scientists (B) (20770034), Grant-in-Aid for Scientific Research on Innovative Areas (26113513) and the Global Center of Excellence Program in Nara Institute of Science and Technology (Frontier Biosciences: strategies for survival and adaptation in a changing global environment), MEXT, Japan (to M.F.).. 13.
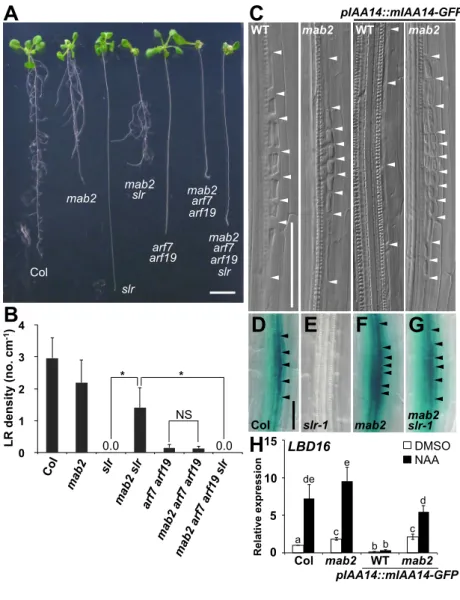
(14) Figure legends Fig. 1. The effect of mab2 on the SLR/IAA14-ARF7-ARF19 auxin signaling module in LR formation. (A) Fourteen-day-old seedlings of Col, mab2-4, slr-1, mab2-4 slr-1, arf7-1 arf19-1, mab2-4 arf71 arf19-1 and mab2-4 arf7-1 arf19-1 slr-1. Scale bar, 1 mm. (B) LR density (emerged LR number per primary root length) for Col (n=20), mab2-4 (n=21), slr-1 (n=20), mab2-4 slr-1 (n=22), arf7-1 arf19-1 (n=20), mab2-4 arf7-1 arf19-1 (n=21), and mab2-4 arf7-1 arf19-1 slr-1 (n=20). Seven-day-old seedlings were used in this analysis. Data are presented as means ± SD. A two-tailed, Student’s t test was performed, and asterisks represent significant differences: *P < 0.05; NS, not significant. (C) Nomarski images of roots of WT, mab2-1, and pIAA14::mIAA14GFP transgenic plants in WT and mab2-1 backgrounds at 8 d. The arrowheads indicate the cell wall. Scale bar, 50 µm. (D-G) Expression of pLBD16::GUS in the mature root region of Col (D), slr-1 (E), mab2-4 (F) and mab2-4 slr-1 (G) for 10-d-old seedlings. The arrowheads indicate the cell wall of LR primordia. Scale bar, 50 µm. (H) Expression of LBD16 in Col and mab2-4 expressing pIAA14::mIAA14-GFP. The roots at 7 d were harvested after auxin treatment (black) and without auxin treatment (white). The relative abundance of LBD16 mRNA to ACT8 mRNA was measured by quantitative RT-PCR. The value measured for Col without auxin treatment was set at 1. Data represent means ± SD (n = 3 independent biological replicates). Different letters in each graph indicate the statistical differences (P<0.05, Student’s t-test). Fig. 2. Protein-protein interaction between MAB2 and the IAA14-TPL repressor complex. (A and B) Co-IP assays showing that mIAA14 interacts with MAB2 (A) and TPL (B). Protein extracts from 7-d-old-roots of mab2-4 expressing MAB2-c-Myc, tpl-1 expressing TPL-HA, Col expressing mIAA14-GFP, mab2-4 coexpressing mIAA14-GFP and MAB2-c-Myc, and tpl coexpressing TPL-HA and mIAA14-GFP were subjected to IP using an anti-GFP antibody. Input and IP fractions of MAB2-c-Myc (A), TPL-HA (B) and mIAA14-GFP were detected by immunoblots (IB) using anti-c-Myc, anti-HA and anti-GFP antibodies, respectively. The outlined arrowheads indicate the predicted position of mIAA14-GFP, MAB2-c-Myc, and TPL-HA, respectively, identified by proper molecular-weight bands (Fig. S15). (C) Y2H assay showing that MAB2 interacts with TPL and TPR2. The assay was performed with SD-Leu-Trp as well as SD-Leu-Trp-His. BD: binding domain; AD: activation domain. (D) Y3H assay showing that TPL forms a bridge between IAA14 and MAB2 and that the C-terminal to the lissencephaly homology (CTLH) domain of TPL, essential for interaction with Aux/IAA, is required for the coexistence of three proteins. (E and F) Bimolecular fluorescence complementation (BiFC) assay for TPL-dependent interaction between mIAA14 and MAB2 in Arabidopsis protoplast cells. Bright field (left), BiFC (second from left) and ECFP (second from right) fluorescence images of protoplast cells, into which BiFC (Venus-N-mIAA14/MAB2-Venus-C) and additional constructs (TPL-ECFP or nuclear localization signal (NLS)-ECFP) were introduced. Merged image (right) of BiFC and ECFP. Scale bars represent 10 µm. (G) Co-IP assays showing that MAB2 associates with TPL only in the presence of the auxin antagonist auxinole. Protein extracts from 7-d-old-seedlings of mab2-4 expressing MAB2-c-Myc, tpl-1 expressing TPL-HA, and mab2 tpl coexpressing MAB2-c-Myc and TPL-HA pretreated with or without 10 µM auxinole for 18 h were subjected to IP using an anti-c-Myc antibody. Input and IP fractions of TPL-HA and MAB2-c-Myc were detected by IBs using anti-HA and anti-c-Myc antibodies, respectively. The outlined arrowheads indicate the predicted position of TPL-HA and MAB2-cMyc, respectively, by proper molecular-weight bands (Fig. S15). Fig. 3. MAB2 binding in the upstream region of LBD16. (A) Schematic diagram of the LBD16 locus and amplified fragments for ChIP assays (I-V and NC). The locus which gives rise to auxin-induced lncRNA is located upstream of the region I. (B -E) ChIP assays using 7-d-old seedlings of mab2-4 pMAB2::MAB2-c-Myc under the normal. 14.
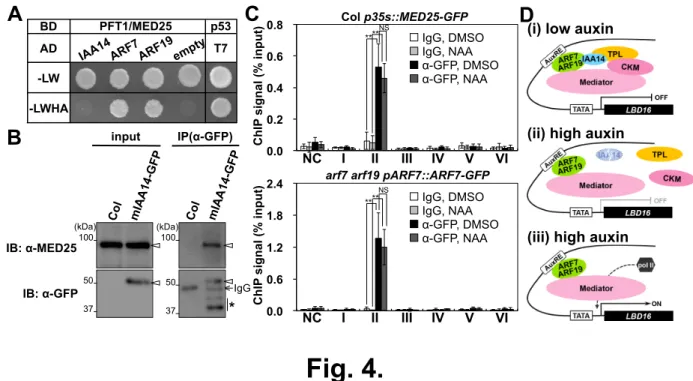
(15) condition (B), treated with 1 µM NAA for 2 h (C) and 10 µM auxinole for 18 h (D), and mab24 pMAB2::MAB2-c-Myc pIAA14::mIAA14-GFP (E). Control reaction was processed in parallel with mouse IgG. The y axis represents the ratio between the enriched DNA after IP and the input DNA before IP. Data represent means ± SD (n = 3 independent biological replicates). Asterisks indicate the statistical differences (*P < 0.05, **P < 0.01, Student’s t-test). A region approximately 3000 bp from the transcriptional start site of LBD16 was used as a negative control (NC). Fig. 4. The interaction between ARF7 and Mediator subunits. (A) Y2H assay showing that MED25 interacts with IAA14, ARF7 and ARF19. The assay was performed with SD-Leu-Trp as well as SD-Leu-Trp-His-Ade supplemented with 10 mM 3-AT. BD: binding domain; AD: activation domain. (B) Co-IP assays showing that mIAA14 interacts with MED25. Protein extracts from 7-d-old-roots of Col expressing mIAA14-GFP were subjected to IP using an anti-GFP antibody. Input and IP fractions of MED25 and mIAA14-GFP were detected by immunoblots (IB) using anti-MED25 and anti-GFP antibodies, respectively. The outlined arrowheads indicate the predicted position of MED25 and mIAA14-GFP, respectively. The asterisks show bands of the putative degradation products of mIAA14-GFP, identified by proper molecular-weight bands (Figure. S15). (C) ChIP assays using 7-d-old seedlings of Col p35s::MED25-GFP (top) and arf7 arf19 pARF7::ARF7-GFP (bottom). Control reaction was processed in parallel with mouse IgG. The y axis represents the ratio between the enriched DNA after IP and the input DNA before IP. Data represent means ± SD (n = 3 independent biological replicates). Asterisks indicate the statistical differences (**P < 0.01, Student’s t-test). A region approximately 3000 bp from the transcriptional start site of LBD16 was used as a negative control (NC). (D) A model for auxin responsive transcription through IAA14-ARF7-ARF19. (i) Under low auxin condition, IAA14 forms a repressor complex with TPL and CKM to inactivate the ARF7 and ARF19 function. Mediator interacts with ARF7 and ARF19 as well as CKM, thus preventing Mediator from associating with RNA pol II. (ii) When auxin concentrations are high, auxin promotes proteolysis of IAA14 and release of TPL and CKM, leading to derepression of both ARF7 and ARF19 and Mediator. (iii) Releasing ARF7 and ARF19 activates the transcription of target genes through the recruitment of RNA pol II via CKM-dissociated Mediator.. 15.
(16) A . C . mab2 slr . mab2 . WT . mab2 . D . E . pIAA14::mIAA14-GFP WT mab2 . mab2 arf7 arf19 mab2 arf7 arf19 slr . arf7 arf19 Col slr . F . G . mab2 . mab2 slr-1 . 3. * . * . 2. NS . 1 0. 0.0 1". 2". 3". 0.0 4". 5". 6". 7". Col slr-1 15 15 LBD16 . H Relative expression . LR density (no. cm-1) . B 4. DMSO NAA . e 10 10 . de d . 5 5 . a 0 0 . Fig. 1. . c c . c b b . 1" mab2 2" 3" mab2 4" WT Col pIAA14::mIAA14-GFP .
(17) A . B . tpl Col tpl mab2 Col mab2 TPL-HA + — + MAB2-Myc + — + (kDa) + mIAA14-GFP — + (kDa) mIAA14-GFP — + + IP: α-GFP IB: α-c-Myc input IB: α-c-Myc IP: α-GFP IB: α-GFP input IB: α-GFP . p53. C BD . 250 . 250 . T7. TPR2. G . 50 . 50 . D BD AD IAA14. MAB2. 100 . input IB: α-GFP . — TPL. 150 . IP: α-GFP IB: α-GFP . 50 . F . 100 . input IB: α-HA . 50 . AD -LW -LWH . 150 . IP: α-GFP IB: α-HA . E . linker . MAB2. — . MAB2. TPL. -LW -LWH . MAB2 TPLΔCTLH. Fig. 2. . mab2 tpl mab2 tpl MAB2-Myc + — — + + + TPL-HA — — + + + + auxinole — + — + — + (kDa) . IP: α-c-Myc IB: α-HA input IB: α-HA . 150 . 100 150 100 250 . IP: α-c-Myc IB: α-c-Myc 250 . input IB: α-c-Myc.
(18) lncRNA . ChIP signal (% input) . B . 0.5. II . III . IgG α-c-Myc (MAB2) . 0.3 0.2. * * . 0.1 0.0. 0.5. 1" 2" NC I . 6" V . 7" VI . IgG α-c-Myc (MAB2) . ** . 0.3. * * . 0.1 0.0. 4" 5" III IV . mab2 pMAB2::MAB2-Myc + auxinole . 0.4. 0.2. 3" II . 1" 2" NC I . 3" II . * . 4" 5" III IV . 6" V . IV . C . mab2 pMAB2::MAB2-Myc . 0.4. D ChIP signal (% input) . I . ChIP signal (% input) . NC . LBD16 . TATA . AuxRE . 7" VI . 0.5. V . VI . 500 bp mab2 pMAB2::MAB2-Myc + NAA IgG α-c-Myc (MAB2) . 0.4 0.3 0.2 0.1 0.0. 1" NC . E ChIP signal (% input) . A . 2" I . 3" II . 4" 5" III IV . ** . IgG α-c-Myc (MAB2) α-GFP (IAA14) . 0.4 0.3. ** . 0.2. ** * . 0.1. Fig. 3. . 7" VI . mab2 pMAB2::MAB2-Myc pIAA14::mIAA14-GFP . 0.5. 0.0. 6" V . 1" NC . 2" I . 3" II . * . 4" 5" III IV . 6" V . 7" VI .
(19) p53 T7 . AD -LW -LWHA . B . input . (kDa) 100 . IB: α-MED25 . 50 . IB: α-GFP 37 . IP(α-GFP) . (kDa) 100 . 50 37 . C 0.8 ChIP signal (% input) . PFT1/MED25 . BD . ChIP signal (% input) . A . IgG . * . D . Col p35s::MED25-GFP NS . ** ** . 0.6 0.4. IgG, DMSO IgG, NAA α-GFP, DMSO α-GFP, NAA . 0.2 0.0. 2.4. (ii) high auxin 1" 2" NC I . 3" II . 4" 5" III IV . 6" V . 7" VI . arf7 arf19 pARF7::ARF7-GFP NS . ** ** . 1.8 1.2. IgG, DMSO IgG, NAA α-GFP, DMSO α-GFP, NAA . 0.6 0.0. (i) low auxin . 1" 2" NC I . 3" II . Fig. 4. . 4" 5" III IV . 6" V . 7" VI . (iii) high auxin .
(20) Supplemental information Supplementary Method Immunolocalization analysis. 5-d-old seedlings were fixed for 1 h with 4% paraformaldehyde in 1x microtubules stabilizing buffer and cut root tips of 7-d-old seedlings were fixed overnight at -20 °C in a methanol-acetic acid solution (3:1). After dehydration in ethanol series and infiltration with butanol, samples were embedded in paraffin. Subsequent processes were performed as previously described (42). Antibodies and nuclear fluorescent dye were diluted as followed: 1:150 for rabbit anti-GFP (Thermo Fisher Scientific), 1:200 for mouse anti-HA (Roche 12CA5), 1:100 for anti-c-Myc (Santa Cruz Biotechnology 9E10), 1:300 for anti-MED25 antibodies, 1:500 for Alexa 488-conjugated anti-rabbit secondary antibody (Invitrogen), and 1:1000 for SYTOX Orange (Life technology), respectively. Microscopy. The GUS staining, fixation, and whole-mount clearing preparation of roots were performed in essentially the same manner as has been described previously (43), and samples were observed with an Olympus BX-52 microscope equipped with Nomarski optics (Olympus). For confocal microscopy, roots were analyzed with an Olympus FV1000 confocal microscope (Olympus). RNA extraction and quantitative RT-PCR. Total RNA was extracted from roots of 7-d-old seedlings treated with dimethyl sulfoxide (DMSO) or 1 µM NAA for 2 h using RNeasy Plant Mini Kit (Qiagen). Each treatment was done in three replicates. First-strand cDNA was synthesized from DNase-treated total RNA (2 µg) using SuperScript II reverse transcriptase (Invitrogen) with an oligo (dT)24 primer (Invitrogen) for UBC and LBD16 or with At2g42425-r2 primer (5’-GTACACGAGCACTAAGTGAAGA-3’) for lncRNA. Quantitative RT-PCR was performed on the LightCycler 96 (Roche) with the KAPA SYBR FAST qPCR kit (KAPA BIOSYSTEMS) and the gene-specific primer sets according to the manufacture’s instructions. ACT8 (Fig. S4) and UBC (Fig. 1H) were measured for an internal control and used to normalize the data. Relative amount of LBD16 and lncRNA (At2g42425) transcripts was calculated as fold change relative to values measured for wild type plants (Col) without auxin treatment, which was arbitrarily set to 1. All primer sequences are listed in the Supplementary Table. Bimolecular fluorescence complementation (BiFC). MAB2 and mIAA14 open reading frames were cloned into BiFC vectors (44). TPL open reading frame and AtWRKY6 NLS sequence (45), corresponding to the region spanning amino acid 1-543, were cloned into ECFP-tagged fluorescent protein expression vector. For the BiFC experiments, 3 µg of each Venus-Nterminal- and Venus-C-terminal-tagged protein expression vector (VN-mIAA14 and MAB2VC), and 3 µg of ECFP-tagged protein expression plasmid (TPL-ECFP or NLS-ECFP), were co-transformed into Arabidopsis protoplasts as previously described (46). After incubation at 23 °C for 12 hours in the dark, the Venus (BiFC) and ECFP fluorescence was analyzed by the confocal microscopy..
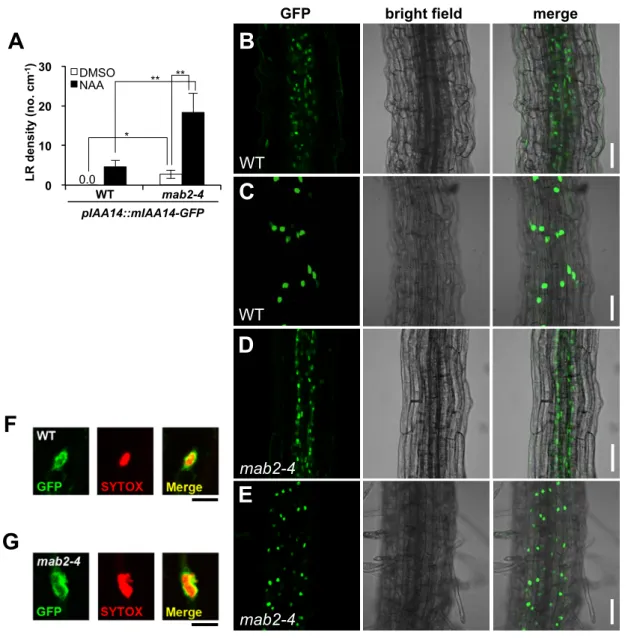
(21) Supplemental Figure Legends Fig. S1. LR formation in Arabidopsis. (A) Nomarski images of LR development in Arabidopsis Col (top) and mab2-4 (bottom). LRs are initiated from the anticlinal cell division (white arrows) in the pericycle (black arrowheads) and grow through outer cell layers. (B) The frequency distribution of cell number of pericycle between neighboring LR primordia of WT (n = 16), mab2-4 (n = 16) and mab2-4 slr-1 (n = 16). (C) LR density for WT (n = 20), slr-1 (n = 20), mab2-4 (n = 21) and mab2-4 slr-1 (n = 22). Data are means ± SD. Student’s t test was performed, and asterisks represent significant differences: *P < 0.05, **P < 0.01. Fig. S2. The effects of mab2 on auxin responsibility in LR formation and stabilized-IAA14 expression. (A) LR density for WT (n = 22), mab2-4 (n = 22) and pIAA14::mIAA14-GFP transgenic plants in WT (n = 22) and mab2-4 (n = 21) backgrounds. Four-day-old seedlings were transferred to the media either with or without 1 µM NAA and incubated for an additional 72 h. Data are means ± SD. Student’s t test was performed, and asterisks represent significant differences: *P < 0.05, **P < 0.01. (B-E) Expression of mIAA14-GFP driven by its own promoter in the stele (B and D) and epidermis (C and E) of WT (B and C) and mab2-4 (D and E) roots. Scale bars: 50 µm. (F and G) Immunolocalization of mIAA14-GFP (green) in nuclei (red) of WT (F) and mab2-4 (G). Scale bars: 10 µm. Fig. S3. Complementation test with pMAB2::MAB2-c-Myc. pMAB2::MAB2-c-Myc can fully complement LR phenotypes of the mab2-4 mutants expressing pIAA14::mIAA14-GFP. Pictures show 14-d-old seedlings of Col, mab2-4, mab2-4 pMAB2::MAB2-c-Myc, pIAA14::mIAA14-GFP, mab2-4 pIAA14::mIAA14-GFP, and mab2-4 pMAB2::MAB2-c-Myc pIAA14::mIAA14-GFP. Red arrows indicate the restoration of LR formation. Scale bars: 1 mm. The graphs show LR density for each background (n = 20) using 7-d-old seedlings. Data are presented as means ± SD. Student’s t test was performed, and asterisks represent significant differences: *P < 0.05. Fig. S4. Expressions of ARF7 and ARF19 target genes and auxin signaling module genes in Col and mab2-4 expressing pIAA14::mIAA14-GFP. The roots at 7 d were harvested after auxin treatment (white box) and without auxin treatment (black box). The relative abundance of LBD16, LBD29, ARF7, ARF19, IAA14 and GFP mRNA to ACT8 mRNA were measured by quantitative RT-PCR. The value measured for Col expressing pIAA14::mIAA14-GFP without auxin treatment was set at 1. Data represent means ± SD (n = 3 independent biological replicates). Different letters in each graph indicate the statistical differences (P<0.05, Student’s t-test). Fig. S5. The genetic interaction between other CKM genes and SLR/IAA14. (A) LR density for Col (n=20), slr-1 (n=20), crp-4 (n=20) and crp-4 slr-1 (n=20). (B) LR densitiy for Ler (n=20), slr-1 (n=20), hen3-1 (n=20) and hen3-1 slr-1 (n=22). Four-day-old seedlings were transferred to the media either with or without 1 µM NAA and incubated for an additional 72 h. (C) LR density for Col (n=20), med17 (n=20), pft1-2 (n=20), and pIAA14::mIAA14-GFP transgenic plants in Col (n=20), med17 (n=20), pft1-2 (n=20) backgrounds. Data are means ± SD. Student’s t test was performed, and asterisks represent significant differences: *P < 0.05, **P < 0.01. Fig. S6. The Y2H assay to examine the interaction between IAA14 and the subunits of the CKM module. IAA14 cannot interact with the subunits of the CKM module, but it can with TPL and TPR2, in.
(22) yeasts. The assay was performed with SD-Leu-Trp as well as SD-Leu-Trp-His. BD: binding domain; AD: activation domain. Fig. S7. The genetic interaction between TPL and mIAA14 in LR development. (A) 14-d-old seedlings of mIAA14-GFP expressing WT, mab2-1, tpl-1, and mab2-1 tpl-1. (B and C) LR density for WT (n = 20), mab2-1 (n = 20), tpl-1 (n = 20), mab2-1 tpl-1 (n = 20) and transgenic plants in WT (n = 20), mab2-1 (n = 20), tpl-1 (n = 20) and mab2-1 tpl-1 (n = 20) backgrounds. Four-day-old seedlings were transferred to the media either with DMSO (white) or 1 µM NAA (black) and incubated for an additional 72 h. Data are means ± SD. Student’s t test was performed, and asterisks represent significant differences: *P < 0.05, ** P < 0.01; NS, not significant. (D) Nomarski images of roots of WT, mab2-1, tpl-1, mab2-1 tpl-1 and pIAA14::mIAA14-GFP expressing plants in WT, mab2-1 , tpl-1 and mab2-1 tpl-1 backgrounds at 8 d. The arrowheads indicate the cell wall. In mIAA14-GFP expressing mab2-1 tpl-1, LR primordia (yellow asterisks) were adjacently located. Scale bars: 50 µm. Fig. S8. The effect of auxinole on the stability of IAA14-GFP Accumulation of IAA14-GFP protein in response to auxinole application. Protein extracts from 7 d-old-seedlings of pIAA14::IAA14-GFP expressing Col treated with 10 µM auxinole or DMSO for 18 h were subjected to an immunoblot assay with anti-GFP antibody. Fig. S9. ChIP analysis using Col (A-D) ChIP assays using 7-d-old seedlings of Col under the normal condition with an anti-cMyc or anti-GFP antibodies (A), auxin-treated Col (1 µM NAA for 2 h) with an anti-c-Myc antibody (B), auxinole-treated Col (10 µM auxinole for 18 h) with an anti-c-Myc antibody (C), and auxin- or DMSO-treated Col with an anti-GFP antibody (D). Nuclear proteins were immunoprecipitated with respective antibodies, and the enriched DNA fragments were used for quantitative RT-PCR analysis. Control reaction was processed in parallel with mouse IgG. The y axis represents the ratio between the enriched DNA after IP and the input DNA before IP. Data represent means ± SD (n = 3 independent biological replicates). Fig. S10. Auxin responsibility of lncRNA expression. Expressions of At2g42425 lncRNA transcribed from the locus upstream of LBD16 in Col, mab2-4 and auxin-treated Col roots. Wild-type roots at 7 d were harvested after treatment with 1 µM NAA for 2 h. The relative abundance of lncRNA mRNA to UBC mRNA were measured by quantitative RT-PCR. The value measured for Col without auxin treatment was set at 1. Data represent means ± SD (n = 3 independent biological replicates). Student’s t test was performed, and asterisks represent significant differences: ** P < 0.01. Fig. S11. The Y2H assay to examine the interaction of the subunits of the core Mediator with IAA14, ARF7 and ARF19. The assay was performed with SD-Leu-Trp as well as SD-Leu-Trp-His, using MED8 and MED6, components of the head module, as the bait. BD: binding domain; AD: activation domain. Fig. S12. The genetic interaction of MED genes with ARF7 and ARF19. (A) LR density for WT (n = 20), arf7-1 (n = 20), arf19-1 (n = 20), pft1-2 (n = 20), arf7-1 arf191 (n = 22), pft1-2 arf7-1 (n = 22) and pft1-2 arf19-1 (n = 22). (B) LR density for WT (n = 20), arf7-1 (n = 20), arf19-1 (n = 20), med8 (n = 21), arf7-1 arf19-1 (n = 22), med8 arf7-1 (n = 22) and med8 arf19-1 (n = 22). The graphs show LR density for each background using 8-d-old seedlings. Data are means ± SD. Student’s t test was performed, and an asterisk represents a significant difference: *P < 0.05..
(23) Fig. S13. The effect of auxin on the interaction between ARF7 and Mediator subunits. (A) Co-IP results showing the association of Mediator subunits with ARF7 in response to auxin treatment. Protein extracts from 7-d-old seedlings of Col and arf7-1 arf19-1 mab2-4 coexpressing pARF7::ARF7-GFP and pMAB2::MAB2-c-Myc, pretreated with 10 µM auxinole for 16 h (lane: pre) prior to incubating with 10 µM auxinole (lane: auxinole) or 1 µM NAA (NAA) for 2 h, were subjected to IP using an anti-GFP antibody. Input and IP fractions of MAB2-c-Myc (the two top columns), MED25 (the two middle columns) and mIAA14-GFP (the two bottom columns) were detected by IBs using anti-c-Myc, anti-MED25 and anti-GFP antibodies, respectively. The bands of interest were identified by proper moleculer-weight bands of Precision Plus ProteinTM Dual Color Standards (Bio-Rad), and were indicated by arrowheads. Colored dots indicate the position of the moleculer-weight bands, respectively. (B and C) Accumulation of MAB2 and MED25 proteins in response to auxin application. Protein extracts from 7 d-old-seedlings of mab2-4 pMAB2::MAB2-c-Myc and Col treated with 1 µM NAA or DMSO for 2 h were subjected to an immunoblot assay with anti-c-Myc (B) and anti-MED25 (C) antibodies, respectively. Fig. S14. Expression of auxin-signaling module, MAB2 and MED25 in LR formation (A) Relative expression of ARF7, ARF19, IAA14, TPL, MAB2 and MED25 in Arabidopsis root tips, displayed by Arabidopsis eFP browser (www.bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi). (BG) Immunolocalization of TPL-HA in roots of tpl-1 expressing TPL-HA (B) and Col (C), MAB2-c-Myc in roots of mab2-4 expressing MAB2-c-Myc (D) and Col (E), MED25 in roots of Col (F) and a null allele pft1-2 (G), respectively. Nuclei were stained by SYTOX (blue). Scale bars: 20 µm. Fig. S15. Identification of the bands of interest. The bands of interest in Fig. 2A, B, G and 4B were identified by proper molecular-weight bands of Precision Plus ProteinTM Dual Color Standards (Bio-Rad), and were indicated by outlined arrowheads. Colored dots indicate the position of the molecular-weight bands, respectively..
(24) A . Col . Frequency . B . 10. Col mab2-4 mab2-4 slr. 8 6 4 2 0. 0 0 . 1 1 . 2 2 . 3 3 . 44. 55. 66. 77. C . LR density (no. cm-1) . mab2-4 30. DMSO NAA. * . ** * . 20. 10. 0. Number of PC between two LRP . Fig. S1. . Col . 1 . 0.0 slr-1 mab2-4 . 2 . 3 . slr-1 mab2-4 4 .
(25) GFP LR density (no. cm-1) . A . B 30. DMSO NAA. ** ** . 20 * . 10. WT 0. 0.0 WT . 1 . mab2-4 . 2 . C . pIAA14::mIAA14-GFP . WT . D F mab2-4 . E G mab2-4 . Fig. S2. . bright field . merge .
(26) LR density (no. cm-1) . pMAB2::MAB2-Myc pMAB2:: MAB2-Myc pIAA14::mIAA14-GFP Col mab2-4 mab2-4 WT mab2-4 mab2-4 . 4 3. * . * . 2 1 0.0 0.0 Col mab2 mab2 WT mab2 mab2 1 2 MAB23 4 mIAA14-GFP 5 6 Myc MAB2Myc . 0. Fig. S3. . Relative expression level (normalized by ACT8 expression) . 6. 2. LBD16 . c . 4. b 2 0 8. a 1". 0. 2". 2. LBD29 . 4. 0. a . a . IAA14 . a . a . 1. a . a . a . a . a . c . 6. 2. 1. 2. ARF7 . ab . b . 1". a 1. 2. ARF19 a . 0. 2". a a 1. 1". 2". GFP a . a . a . a . a 0. 0. 1" 2" 1" 2" 1" 2" WT mab2-4 WT mab2-4 WT mab2-4 pIAA14::mIAA14-GFP pIAA14::mIAA14-GFP pIAA14::mIAA14-GFP . Fig. S4. .
(27) B DMSO NAA. 20. * . ** . * . 10 0. Col . 1 . 0.0 slr-1 crp-4 crp-4 2 3 4 slr-1. 30. C DMSO NAA. 20. * . * . * . 10 0. Ler . 1 . 0.0 slr-1 hen3-1 hen3-1 2 3 4 slr-1. LR density (no. cm-1) . 30. LR density (no. cm-1) . LR density (no. cm-1) . A . DM. 5. * . 4. * . 3 2 1 0 1 . 2 . 3 . 4 . 5 . 6 . pIAA14::mIAA14-GFP . Fig. S5. .
(28) BD . IAA14. p53 T7 . AD . -LW . -LWH . Fig. S6. .
(29) B LR density (no. cm-1) . A pIAA14::mIAA14-GFP . LR density (no. cm-1) . C . 35 30. 20. * . 15 10 5 0. Ler 1 mab2-1 2 tpl 3 . 0.0 WT 4 mab2-1 5 tpl 6 . pIAA14::mIAA14-GFP . 6. NS NS NS . 4. 2. D . 0.0 tpl Ler tpl WT 1" mab2 2" 3" mab2 4" 5" mab2-1 6" 7" mab2 8" tpl tpl pIAA14::mIAA14-GFP . pIAA14::mIAA14-GFP mab2-1 . tpl . mab2-1 tpl . WT . ** . 25. 0. WT . * ** . DMSO NAA . mab2-1 . Fig. S7. . tpl . mab2-1 tpl .
(30) Col pIAA14::IAA14-GFP auxinole . — . + . 100 75 . IAA14-GFP . 50 37 25 (kDa) . IB: α-GFP . Fig. S8. .
(31) 0.3 0.2 0.1 0.0. 1" 2" NC I . 3" II . 4" 5" III IV . 6" V . 7" VI . Col + auxinole . 0.5. IgG α-c-Myc . 0.4 0.3 0.2 0.1 0.0. 1" NC . 2" I . 3" II . 4" 5" III IV . 6" V . 7" VI . ChIP signal (% input) . IgG α-c-Myc α-GFP . 0.4. C ChIP signal (% input) . B . Col . 0.5. Col + NAA . 0.5. IgG α-c-Myc . 0.4 0.3 0.2 0.1 0.0. D 0.8 ChIP signal (% input) . ChIP signal (% input) . A . 1" NC . 2" I . 3" II . 4" 5" III IV . 6" V . 7" VI . Col IgG, DMSO IgG, NAA α-GFP, DMSO α-GFP, NAA . 0.6 0.4 0.2 0.0. Fig. S9. . 1" 2" NC I . 3" II . 4" 5" III IV . 6" V . 7" VI .
(32) Relative expression level. (normalized by UBC expression) . 60. ** . 0 1". ** . 2". ** . 40. 20. 3". Fig. S10. .
(33) BD . MED8 . MED6 . AD . p53 T7 . -LW . -LWH . Fig. S11. .
(34) LR density (no. cm-1) . 5 4. * . 3 2 1 0. Col arf7 arf19 med8 pft1-2 arf7 arf7 arf7 arf19 arf19 1 2 3 4 5 arf19 6 med8 7 pft1-2 8 med8 9 pft1-2 10 . Fig. S12. .
(35) A . input arf7 arf19 mab2 ARF7-GFP Col MAB2-Myc. IP(α-GFP) arf7 arf19 mab2 ARF7-GFP Col MAB2-Myc. IB: α-c-Myc . IB: α-MED25 250 150 100 . IB: α-GFP . Marker (kDa) . B . mab2 pMAB2::MAB2-Myc 1 µM NAA pre — + . IB: α-c-Myc . C . Col 1 µM NAA . — . pre . IB: α-MED25 . Fig. S13. . + .
(36) Fig. S14. .
(37) Fig. 2A . Fig. 2B . mab2 Col mab2 MAB2-Myc + — + mIAA14-GFP — + + . Fig. 2G tpl. TPL-HA + mIAA14-GFP — . IP: α-GFP IB: α-c-Myc . Col tpl — + . + + . IP: α-GFP IB: α-HA . input IB: α-GFP . input IB: α-HA . IP: α-GFP IB: α-GFP . IP: α-GFP IB: α-GFP . input IB: α-GFP . input IB: α-GFP . mab2 tpl mab2 tpl MAB2-Myc + — — + + + TPL-HA — — + + + + auxinole — + — + — + . IP: α-c-Myc IB: α-HA . input IB: α-HA . IP: α-c-Myc IB: α-c-Myc. input IB: α-c-Myc. Fig. 4B . 250 150 . IP: α-GFP IB: α-MED25 . 100 75 50 37 25 . input IB: α-MED25 . Marker (kDa) . IP: α-GFP IB: α-GFP . input IB: α-GFP . Fig. S15. .
(38) Supplementary Table Name Cloning pMab2-F3 pMAB2-R3 MAB2 F cMAB2 R2 TPL F TPL R TPR2 F TPR2 R IAA14 F IAA14 R ARF7 F ARF7 R ARF19 F ARF19 R PFT1 F PFT1 R MED6 F MED6 R MED8 F MED8 R TPL CTLHout F TPL CTLHout R qRT-PCR LBD16-LC-F LBD16-LC-R LBD29-LC-F LBD29-LC-R ARF7-LC-F ARF7-LC-R ARF19-LC-F ARF19-LC-R IAA14-L1 IAA14-R1 GFP-LC-f GFP-LC-r ACT8-LC-f ACT8-LC-r At2g42425RT-f At2g42425RT-rev UBC-F UBC-R. Sequence (5'-to-3') AAGCTTGACGATGTTCTCTGGCTTTCTTC AAGCTTCACTTGGGGATAGATACAACAAG AAAAAGCAGGCTTGATGTGGACTAATGTTTTCAGAATTGGTG AGAAAGCTGGGTAAACTCCTGTCTTATCAGGTATCCTGG AAAAAGCAGGCTACATGTCTTCTCTTAGTAGAGAGCTCG AGAAAGCTGGGTATCTCTGAGGCTGATCAGATGCAG AAAAAGCAGGCTACATGTCGTCTTTGAGCAGAGAG AGAAAGCTGGGTACCTTTGAATCTGATCCGAACTTG AAAAAGCAGGCTCAATGAACCTTAAGGAGACGGAGC AGAAAGCTGGGTATGATCTGTTCTTGAACTTCTCC AAAAAGCAGGCTCAATGAAAGCTCCTTCATCAAATGGA AGAAAGCTGGGTATCACCGGTTAAACGAAGTGGCTG AAAAAGCAGGCTCAATGAAAGCTCCATCAAATGGATTTC AGAAAGCTGGGTACTATCTGTTGAAAGAAGCTGCAGC AAAAAGCAGGCTCTATGTCGTCGGAGGTGAAACAG AGAAAGCTGGGTATCCCATGAAGCCAGCTCCAG AAAAAGCAGGCTCCATGGATTCCTCTCTGCTCTCG AGAAAGCTGGGTAGAACTTCATTCTTTTAGCAGGAC AAAAAGCAGGCTTTATGGAGACACAGCCGCAGCAAC AGAAAGCTGGGTATTGAGGATTTTGGTGCCTTTG TAGTATATCCACAGCCCCAGATTCTTGTTCAAG GAGCAAGAATCTGGCGCTGTTGAGATATTAGCC TGACCCTGTTTATGGATG TGATTGCAAGAAAGCCACCT GCTAGGCTTCAAGATCCCATC TGTGCTGCTTGTTGCTTTAGA GCTCATATGCATGCTCCACA GCAATGCATCTCTGTCATATTTG CACCGATCACGAAAACGATA TGTTCTGCACGCAGTTCAC CAAAGATGGTGACTGGATGC GCATGACTCGACAAACATCG TGGTCCTGCTGGAGTTCG CTTGTACAGCTCGTCCATGC TCAGCACTTTCCAGCAGATG ATGCCTGGACCTGCTTCAT ACATTCATGGCATTAGCATTTG TGTAACCAAATATTCCTAAGCCAC CTGCGACTCAGGGAATCTTCTAA TTGTGCCATTGAATTGAACCC.
(39) ChIP-qPCR LBD16-nonARE_f3 LBD16-nonARE_r3 LBD16-I_f LBD16-I_r LBD16-II_f LBD16-II_r LBD16-III_f LBD16-III_r LBD16-IV_f LBD16-IV_r LBD16-V_f LBD16-V_r LBD16-VI_f LBD16-VI_r. CCTATGTGGGCTTTAACGGC ATGTGGTTGGGAGGAAGGAC TGAAGCTCCAAGAACCAAACG ATGAGGTAGTCTTAGGTCTTCT CCCAATAAATTAGAAGTCTCATGTTGC CAAATTTATCGAGTGAGCCAAAGG GTGCAAACCAATTTTGGTTGTTATAC GACTATAGTAAAATTGCTTACACGACG CTCTTTCTCTCTCATGATAATGTTCC GAGATGTGCACGAGCGAACTA CTTTCAGCAACACGAGCAGT ACTCCTCCTCCGGTTTGATG AGTAGCGATTTAGGTGAGCT ACATCTCATTTGTTCTCTGACTGTC.
(40)
図

+7






関連したドキュメント