近畿大学学術情報リポジトリ
67
0
0
全文
(2) DoctoriaI Dissertation. Torularhodin biosynt:hesis o f lac-t:a-t:e assiIllila-t:iog yeast: RhodororuLa gLurinis 00.21. Hideyuki Sakaki. March, 2002. Senjyu Pharmaceutical Co. LTD., Division of Pre-Clinical Study (Advisor: Prof. Sadao Komemushi). Submitted to the Graduate School, Kinki University, to fulfill the requirement for the Doctorate Degree..
(3) (~)(m§). Torularhodio biosyo1:hesis o f lac1:a1:e yeas1: Rhodo~oruLa gLu~ini.s 00.21. assiID.ila1:iog. Hideyuki Sakaki. . March, 2002. Graduate School, Kinki University Division of Agricultural Science (Advisor: Prof. Sadao Komemushi). (~/J)(m§ ). ~LItJ({lJM:M£l. Rhodotorula glutinis no.21,~;: s~t Q torularhodin ~tt~. ------------------------------------------------------ ------------------------------------------~. Submitted to the Graduate School, Kinki University, to fulfill the requirement for the Doctorate Degree..
(4) TABLE OF CONTENTS 1) Preface. 2. 2) Peculiar properties of assimilating lactic acid. 6. 2.1 Introduction 2.2 Optimum conditions to assimilate DL-lactic acid 2.3 Assimilation of D-lactic acid and growth inhibition by L-Lactic acid 2.4 Growth acceleration by barium ion. 3) Effect of culture conditions on the carotenoids biosynthesis. 21. 3.1 Introduction 3.2 Carbon sources and culture conditions 3.3 Dissolved oxygen 3.4 Discussion. 4) Activation of torularhodin productivity and enhancement of tolerance. 33. 4.1 Introduction 4.2 Effect of active oxygen species generators 4.3 Tolerance against oxygen stress of torularhodin higher production mutant. 5) Scavenging of peroxyl radicals by torularhodin. 42. 5.1 Introduction 5.2 Relation of growth inhibition and carotenoid content 5.3 Peroxyl radicals scavenging 5.4 Inhibitory effect on lipid peroxide formation 5.5 Discussion. 6) Effect of weak white light irradiation on yeast growth and torularhodin. 52. 6.1 Introduction 6.2 Growth inhibition and activation of torularhodin productivity by weak white light. 7) Discussion. 59. 8) Acknowledgement. 64. Also including "MARETIALS AND METHODS" in the end of chapters 2)-6) and "REFERENCES" in chapters 1)-7).. 1.
(5) PREFACE Carotenoids, classified as terpenoids, are mainly C 40 tetraterpen, and mainly distribute to the body of fish and vegetable, microorganisms.. Carotenoids take color. with prolongation of conjugated polyene chain, and p-carotene, representative carotenoid, shows a yellow color. in nature besides p-carotene.. Over 700 kinds of carotenoids have been detected It became clear that carotenoids contain various. functions in addition to antenna function in photosynthesis.. Effect of decreasing risk. of tumor formation has been found in epidemiological study (1).. Much attention has. therefore focused on research on the biological activity of p-carotene, but a large-scale clinical study revealed no evidence of antitumor activity of p-carotene (2). indicated that p-carotene is not an epoch-making drug for human health.. This. On the other. hand, (X-carotene was shown to more effectively prevent carcinogenesis than. P-. carotene in an in vivo study (3), and strong anti oxidative activity was reported in astaxanthin that classified into highly oxidized xanthophyll (4).. Thus, active research. has been carried out on carotenoids. Major functions have been indicated in fungi and non-photosynthetic bacteria, which clearly lack the antenna function of carotenoids.. Schrott (5) proposed that. carotenoids protect against active oxygen species, mainly singlet oxygen produced under irradiation of strong sunlight.. Several studies have reported on the role of. carotenoids in yeast in the past few years (6,7).. Prevention of oxygen stress was. clearly demonstrated by results showing increase in carotenoid content by oxygen stress loading (6).. This study, however, did not evaluate the fluctuation of individual. carotenoid although yeast accumulated multiple carotenoids as final products.. Thus,. difference of contribution of each carotenoid was not defined. In 1983, to examine the physiological character in the low pH range of the microorganism, a yeast which is viable in the medium including lactic acid as single carbon souses has been isolated from soil sample in the Kinki area, Japan.. The strain. no. 21 in which higher growth was exhibited has been selected out of 34 strains. Yeast obtained (no. 21) was identified by comparing their morphological, biological and cultivating properties with those described in The yeast, A Taxonomic Study (8) and yeast: Characteristics and identification (9).. Characteristics of strain no. 21 are. summarized in Table 1-1 and they showed that properties of strain no. 21 were consistent with that of Rhodotorula glutinis. 2. Thus, this yeast was identified with.
(6) Rhodotorula glutinis and was named Rhodotorula glutinis no. 21.. The yeast isolated. had been deposited with institute for fermentation, Osaka.. Rhodotorula sp., red yeast, biosynthesizes carotenoids in its cell. (8). of these, finally synthesizes two kinds of carotenoid,. ~-carotene. R. glutinis,. and torularhodin.. As regards R. glutinis, it is known that a change in culture temperature has an influence on the production of carotenoids (10).. So far, however, only comparison. was made between carotenoids in produced quantity, and no mention has been made about concrete significance of the comparison.. In view of recent remarkable studies. on carotenoids, author has been paying attention to actions of carotenoids produced by. R. glutinis, especially torularhodin. carboxylic acid.. Torularhodin is a unique carotenoid with. This carotenoid is biosynthesized from y-carotene, precursor of ~. carotene, via tolurene,.. Although many carotenoids were taken up as subjects of. study, torularhodin has not been taken up for study. In this thesis, lactate-assimilating properties of R. glutinis no.21 was described at first and physiological properties of carotenoids in this yeast was mentioned. Consequently, it has become apparent that torularhodin has interesting characteristics, compared with. ~-carotene.. It can therefor be presumed that torularhodin play an. important role in R. glutinis.. 3.
(7) Table 1-1 no.21. Morphological characteristics and physiological properties of Rhodotorula glutinis. Rhodotorula glutinis. 8). Rhodotorula glutinis no.21. Reproduction. Multilateral budding. Multilateral budding. Cell Shape. Oval, Globose. Oval, Globose, Elliposid. Cell Size. (2.3-5.0)*(4.0-10.0) Jlm. (2.0-4.5)*(4.0-9.0) Jlm. Spore formation. Negative. Negative. Pseodo-, TrueMycelium formation. Negative. Negative. Ascospore formation. Negative. Negative. Teliospore formation. Negative. Negative. Ballistospore formation. Negative. Negative. Sediment formation. Positive. Positive. Pellicle formation. Positive. Positive. Starch formation. Negative. Negative. Streak culture. Coral red, Salmon pink, Smooth Shiny. Light pink, Smooth shiny. Fermentation. Negative. Negative. Urease activity. Positive. Positive. KN0 3. Positive. Positive. Melibiose. Negative. Negative. Raffinose. Negative. Negative. Melezitose. Positive. Positive. Growth in vitamin-free medium. Positive. Positive. Assimirate of. 4.
(8) REFERENCES 1) Ziegler, R.G.: A review of epidemiologic evidence that carotenoids reduce the risk of cancer. J. Nutr, 119, 116-122 (1989). 2) Mayne, S.T.:. ~-Carotene,. carotenoids and disease prevention in human. FASEB. J.,. 10, 690-701(1996). 3) Murakoshi, M., Nishino, H., Satomi, Y., Takayasu, J., Hasegawa, T., Tokuda, H., Iwashima, A., Okuzumi, J., Okabe, H., Kitano, H., and Iwasaki, R.: Potent preventive action of a-carotene against carcinogenesis. Cancer Res., 52, 65836587(1992). 4) Miki, W.: Biological functions and activities of animal carotenoids. Pure App!. Chern., 63, 141-146(1991). 5) Schrott, E.L.: Carotenoids in plant photoprotection. Pure Appl. Chern., 57,729-734 (1985). 6) Moore, M.M., Breedveld, M.W., and Autor, A.P.: The role of carotenoids in preventing oxidative damage in the pigmented yeast, Rhodotorula mucilaginosa. Arch. Biochm. Biophys., 270, 419-431(1989). 7) Mendez-Alvarez, S., Rufenacht, K., and Eggen, R.I.: The oxidative stress-sensitive yap1 null strain of Saccharomyces cerevisiae becomes resistant due to increased carotenoid levels upon the introduction of the Chlamydomonas reinhardtii cDNA, coding for the 60S ribosomal protein L10a. Biochem. Biophys. Res. Commun., 267, 953-959(2000). 8) Kreger-Van Rij, N.J.W.:The Yeast, a Taxonomic Study (3 rd. Rev. and Enlarg.Ed.), p.897-899, Elsevier Science Publisher, Amsterdam(1984). 9) Barnett,J.A., Payne R.W. and Yarrow D.: Yeast: Characteristics and Identification, 573-574, Cambridge UniversIty press,Cambridge(1990). 10) Simpson, K.L., Nakayama, T.O.M., and Chichester, C.O.: Biosynthesis of yeast carotenoids. J.Bacteriol.,88,1688-1694(1964).. 5.
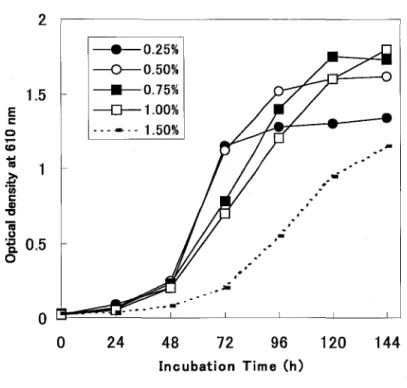
(9) PECULIAR PROPERTIES WHEN ASSIMILATING LACTIC ACID. 2.1 Introduction Lactic acid possesses potent antibacterial activity (1,2) and has been utilized as a bactericidal or a bacteriostatic agent for industrial use.. A lactic acid-assimilating. yeast, Candida utilis has been used to remove lactic acid from wastewater (3) or. Saccharomyces cerevisiae to transport lactate (4).. Though much attention has been. paid to lactic acid and yeast by industries, there are few reports concerning lactic acidassimilating yeast.. Lactic acid has two optical isomers, L- and D- 1 actic acid.. When a microorganism utilizes lactic acid, which isomer it assimilates depends on the type of lactate dehydrogenase (LDH) the microorganism has.. In yeasts, different. lactic acid isomers are assimilated under aerobic conditions from those under anaerobic conditions: in aerobic conditions, they utilize L-Iactic acid rather than Dlactic acid.. L-LDH, which catalyzes the reaction from L-Iactate to L-pyruvate, is. predominant in yeasts (such as S. cerevisiae) in aerobic conditions (5,6).. Humans. also utilize L-Iactate as an energy source, while D-Iactate is poorly absorbed and has little nutritional value (7). Surprisingly, there are few reports on the availability of D-Iactate for assimilation by yeasts in spite of considerable interest in wastewater treatments with yeasts (3).. In. this chapter, selectivity of optical isomer and growth acceleration by barium ion as the characteristics of R. glutinis no.21 incubated in lactate medium was described.. 2.2 Optimum conditions to assimilate DL-lactic acid 1) DL-Lactic acid concentration. R. glutinis no. 21 was grown in the medium including lactic acid as a carbon source with concentration from 0.05 % to 2.0 %.. As the plots of optical density (OD). versus time were almost equal to those of number of viable cells versus time, some growth curves measured by OD are shown in Fig.2-1. The maximum cell number (MeN) was increased and doubling time (DT) was decreased with an increase of lactic acid concentration up to 0.3 %. 6. The yeast.
(10) revealed the greatest MCN at concentrations between 0.5 % and 1.0 %. DT was obtained between 0.3 % and 0.5 %. obtained when lactic acid was over 1.0 %.. The smallest. A longer lag time and a larger DT were The author therefore determined 0.5 % as. the optimum lactic acid concentration for growth of R. glutinis no. 21, in lactic acid medium.. Though a strain of C. utilis was reported to have lactic acid assimilation. capability at 3.5 % (3), no yeasts in this study showed a capability greater than that. (7). 2 • 0.25% --0--- 0.50% 0.75% • -0-1.00% .. - - - - 1.50%. 1.5 E t::. o .co. .'. "Ii:. .-. ~ 'iii t::. CD "t:I. iii (.). '-E. 0.5 o. .. ,. ,. .'. .'. ,-. ". o o. 24. 48. 72. 96. 120. 144. Incubation Time (h). Fig. 2-1 DL-Lactate concentration on the growth of R. glutinis no.21. 2) Temperature and pH profiles. R. glutinis no. 21, showed a growth yield/temperature dependency between 20°C and 45. t. (Table 2-1).. The optimum temperature of around 30. t. for growth of the. strain was similar to that for most yeasts. The optimal pH for growth of the isolate was between 3.0 and 3.5 with reference to DT in log phase and MCN in stationary phase during the growth in lactic acid medium including 0.3 % lactic acid (Table 2-2). different in the 2.5 to 4.5 pH range.. Cell growth was not significantly. Growth was most pronounced at pH 3.5.. 7.
(11) Table2-1. Effect of temperature on the growth yield of R. glutinis no.2l Temperature (C) Optical density at 610 nm 0.50 1.02 1.40 1.05 0.20 0.03. 20 25 30 35 40 45. Incubation was continued until the stationary phase was reached in the 0.3% lactic acid medium.. Table2-2. Effect of pH on the growth yield of R. glutinis no.21 pH Doubling Time MCN. 144 30 24 21 32 32. 2.0 2.5 3.0 3.5 4.0 4.5. 2.0 7.9 7.9. 8.2 7.8 7.7. Incubation was continued until the stationary phase was reached in the 0.3% lactic acid medium. MCN: Maximum cells number (X 107 /ml). 2.3 Assimilation of D-lactic acid and growth inhibition by L-lactic acid The yeast revealed the shortest lag phase when D-Iactic acid was supplied. the other hand, the strain could not grown on L-Iactic acid medium.. On. The OD 610 value. at a stationary phase was 2-fold higher DL-Iactic acid than those for each comparable isomer (Fig. 2-2). These results indicate that the isolate efficiently assimilated the supplied D-Iactic acid as well as the D-Iactic acid in DL-Iactic acid, but not L-Iactic acid.. Since 0.5 %. DL-Iactic acid contains 0.25 % each or D- and L-Iactic acid, the doubled OD value for DL-Iactic acid under the stationary phase interpreted as indicating that L-Iactic acid was promptly assimilated in the presence of D-Iactic acid.. It might be inferred that. L-Iactic acid was converted intracellulafly or extracellularly into D-Iactic acid. The growth rate was compared in the presence of various ratios of D- and L-Iactic acid (Fig. 2-3).. The maximum specific growth rate decreased with an increase of L-. lactic acid in lactic acid medium. increase in total lactic acid.. On the other hand, the OD 610 increased with an. These observations were confirmed by the experiments 8.
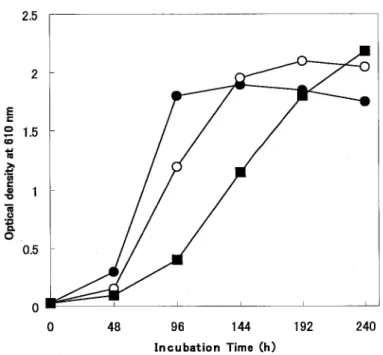
(12) shown in Fig. 2-4.. D- and L-Iactic acid were added to lactic acid medium.. Conditions of adding D- and L-Iactic acid were. = 0.25 %. Case 1. D-:L-. : 0.5 %. Case 2. D-: L- = 0.375 % : 0.375 %. Case 1 had a longer lag phase than that of Case 2.. But a value of OD 610 at Case. 1's stationary phase was similar to that of Case 2. Figure 2-5 shows the change of D- or L-Iactic acid in the medium during the growth of R. glutinis,. strain no. 21.. The growth curve of the strain and D-Iactic acid. reduction curve were patterned symmetrical mirror images.. The L-Iactic acid. decrease was observed on day 4 when cells were presumably in a late log phase.. The. isolate, R. glutinis, strain no. 21, should have D-LDH than L-LDH activity like a sake yeast S. cerevisiae (5,6).. It is supposed from Fig. 2-3 to 2-5 that L-Iactic acid. inhibited yeast growth in an early growth phase whatever D-Iactic acid was present at the same time, and that the yeast might have converted L-Iactic acid into D-Iactic acid and assimilated the resulting D-Iactic acid.. However, further enzymatic study of the. isolate should be done to confirm the observations.. 2.0 0.5% DL-lactic acid. E 1.5 c:: c .,... CD. 1i: .~. 'iii c::. 1.0 '/. iii u. o'-R. /. 0._-. II) "g. -0.._-. -0-- __. 0.25% D-Iactic acid. 0.5 0.25% L-lactic acid. o. 48. 96. 144. 192. 240. Incubation time (h). Fig.. 2-2 Effect of optical isomer for lactate on the growth of R. glutinis no.21. 9.
(13) 2 E. c. 0. CD. 1.5. "t;. ... .~ C. ell -.;,. iii () +I. c.. 0. 0.5. o. 48. 96. 144. 192. 240. Incubation Time (h). Fig. 2-3 Growth inhibition and increase of maximun cell density by L-Iactate -'-0.5% DL-Iactic acid -0- 0.5% DL-lactic acid + 0.25 % L-lactic acid ___ 0.5% DL-lactic acid + 0.5 % L-lactic acid. 10.
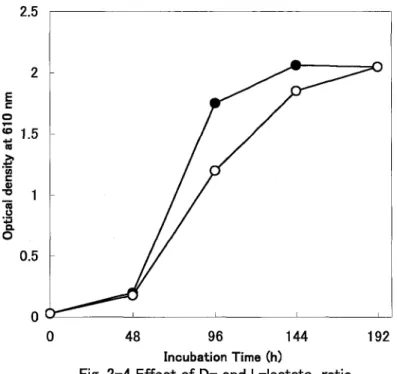
(14) 2.5. 2 E. I::. .... o. co. "Iii. 1.5. .~ Ul I::. CI) "g. ~. "i o 0.5. o. ~------~--------~------~--------~. o. 96. 48. 144. 192. Incubation Time (h). Fig. 2-4 Effect of D- and L-lactate ratio The medium contained 0.75% DL-lactate (e). or Llactate was added at 0.25 %to the 0.5% DL-lactate medium (0). Thus. the amounts of the whole lactate in the medium are united to 0.75%. _D-:L-= 1:1 -o-D-:L-. =2:1. 11.
(15) 2.0. 2.0 L-Iactate. ,....... •. E c. 1.5. 1.5. ~. o .,..... co ......... ~ .iii. ti. D-Iactate 1.0. c. "0. "(3. (JJ. '.p. C-. r::. 8. 1.0. cu. "0. ~. ':::J. (). 0.5. 0.5. 0.0. 0.0. "';;. j. O. o. 24. 48 72 96 120 144 168 Incubation time (h). Fig.2-5 Change of lactate concentration during the growth of R. glutinis no.21. 2.4 Growth acceleration by barium ion 1) Effects of metal ions on the growth Table 2-3 shows the effects of metal ions on the growth of R. glutinis no. 21 in lactate basal broth (LBB) expressed as doubling time (h).. The growth of R. glutinis. no. 21 in LBB was slightly depressed in the presence of several kinds of univalent metal ion whether its compound was lactate or an inorganic salt. seemed to be less depressive to cell growth than univalent ions.. Divalent ions. Barium was the only. element that stimulated the growth of R. glutinis no. 21, i.e., shortened the doubling time of the yeast in LBB among the elements tested in this experiment. Several elements, each of which forms devaluation, showed a stimulation of cell growth at a certain concentration when they were supplemented to the LBB medium in the form of inorganic salts.. Table 2-4 presents the maximum growth rate (expressed. as doubling time: h) for each element and the concentration giving the maximum growth rate.. Table 2-4 includes the concentration range of each element exhibiting. good growth of microorganisms.. Fig.2-6 shows growth curves of R. glutinis no. 21 in 12.
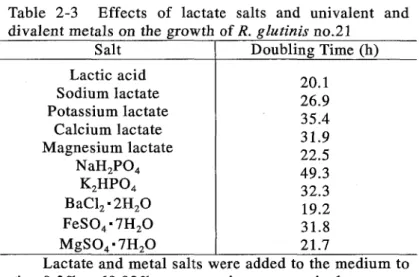
(16) the medium containing the optimum concentration of divalent metals; iron (0.01 mM), magnesium (4.0 mM) and barium (0.10 mM). Table 2-3 Effects of lactate salts and univalent and divalent metals on the growth of R. glutinis no.21 Salt Doubling Time (h) Lactic acid 20.1 Sodium lactate 26.9 Potassium lactate 35.4 Calcium lactate 31.9 Magnesium lactate 22.5 NaH 2 P0 4 49.3 K 2HP0 4 32.3 BaCI 2 "2H 2 0 19.2 FeS0 4 "7H 20 31.8 MgS0 4 "7H 20 21.7 Lactate and metal salts were added to the medium to give 0.3% andO.02% concentrations, respectively.. Optimal concentrations of metal salts for the growth of R. glutinis no.21 Cone. Optimal cone. Doubling Time Metal ion Salt (mM) (mM) (h) FeS0 "7H O 2 4 2 Fe + 0.006-0.014 14.3 0.01. Table 2-4.. Mg2+. MgS0 4 "7H 2O. 0.5-5.0. 4.0. 12.3. Ca2+. CaCI2"2H 2O. 0.1-5.0. 1.0. 14.6. ZnS0 4 "7H 2O. 0.0001-0.05. 0.001. 11.1. CoC1 2"2H 20. 0.0005-0.1. 0.001. 17.9. Zn 2+ C0 2+. Incubation was continued until the stationary phase was reached in the lactate salt.. One can assume that the increase in the growth rate may have been caused by the increase in pH from 3.0 to 3.5, the optimum pH for R. glutinis no. 21. supplemented to LBB had no effects on the initial pH of media.. Inorganic salts. The pHs of three. media, each of which was supplemented with Fe 2 +, MgH or BaH, were not increased but decreased at the end of cultivation from their initial values (Table 2-5).. These. facts indicate that growth enhancement was not caused by increase in pH but by supplementing the medium with such divalent ions as Fe2 +, MgH or BaH.. 13.
(17) 1.6 1.4 ____ Iron. E1.2. -0- Magnesium -X-Barium. -tV 0. cc. .~ 0.8 t:. Q). "'C. 0.6. (ij (). ~ 0.4 0. 0.2 0 0. 48. 192 96 144 Incubation time (h). 240. Fig. 2-6 Growth curve of R. glutinis no.21 in the medium containing divalent metals. The concentration of iron, magnesium and barium were 0.01, 4.0, 0.10 mM, respectively.. Table 2-5 Changes in pH during incubation of R. glutinis no.21 in a metal medium Initial pH Terminal pH Metal ion Fe 2+. 3.1. 2.5. Mg 2+. 3.1. 2.4. Ba 2+. 3.1. 2.5. Blank. 3.1. 3.1. Fe 2+, Mg2+, and Ba2+ were added to the LBB medium in 0.01,5.0 and O.lmM, respectively.. 2) Effects of barium ions The growth of R. glutinis no. 21 was monitored in both LBB and glucose basal broth (GBB; see "materials and methods) to determine if the growth enhancement with Ba2+ shown in Table 2-3 would be observed only in LBB.. The growth rate of R.. glutinis no. 21 was found to be lower in LBB than in GBB.. Increase in the growth. rate was again observed in LBB by the addition of Ba2+; however, no enhancement of. 14.
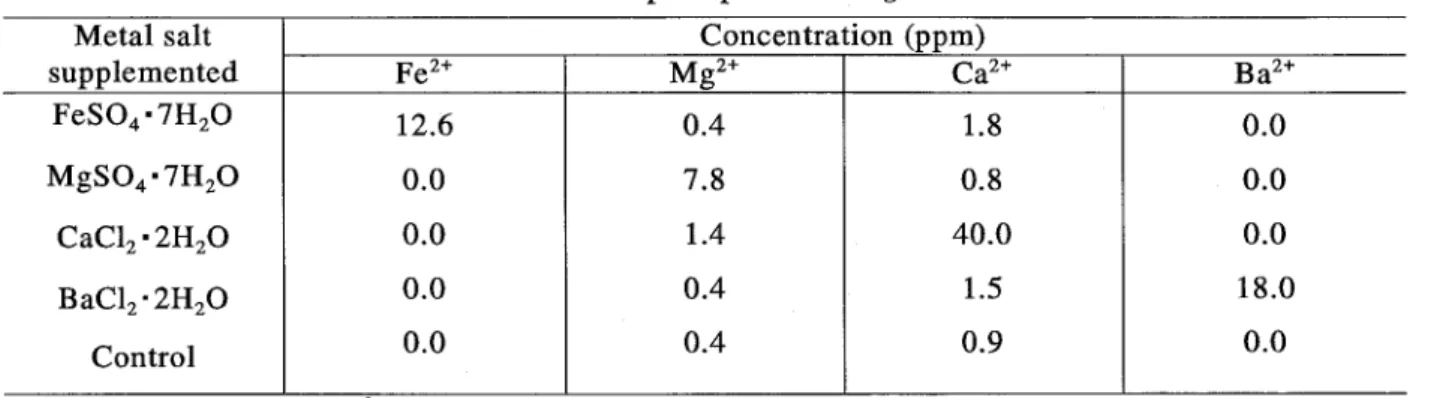
(18) the growth rate was observed in GBB supplemented with Ba2+ when compared with the growth rate in GBB without any supplement.. This suggested that BaH has some. relation to the lactate-assimilating system (s) in which some factors are required more strongly than in the glucose-assimilating system (s). Author then looked for the optimum concentration of Ba2 + for the growth enhancement of R. glutinis no. 21.. The optimal concentration was found in the range. between 0.05 and 0.3 mM Ba2 +, although the rate of growth enhancement varied from experiment to experiment (Table 2-6). Table 2-6. Effects of barium ion on the growth of R. glutinis no.21. Ba2+ concn. Exp 1 29.1. Doubling Time (h) Exp 3. Exp 4 Exp 5 0.01 ._---------------------- ------------------- ------------------ ------------------ -------------------------------------0.1 15.5 ._---------------------- ------------------- ------------------ ------------------ -------------------------------------0.05 28.0 18.2 14.8 (mM). Exp 2. .---------o.i0- -- ------ ------ -iC6 ---- -- - ------is. ~o-. -------------16.2------ --- -- --i6.-2 --------------i4j-------. ._---------------------- ------------------- ------------------ ------------------ -------------------------------------0.15 23.6 ._---------------------- ------------------- ------------------ ------------------ -------------------------------------0.20 26.0 12.5 13.3 14.6 ._---------------------- ------------------- ------------------ ------------------ --------------------------------------. 0.25 34.5 0.30 14.7 ._---------------------- ------------------- ------------------ ------------------ -------------------------------------0.40 21.0 16.4 ._---------------------- ------------------- ------------------ ------------------ -------------------------------------0.50 35.1 18.6 0.60 Incubation was continued until the stationary phase was reached in the LBB.. 3) Intracellular concentrations of metal ion Precipitate of barium sulfate was formed by the reaction of Ba2 + with SO/- in the medium when a barium chloride solution was added.. As described above, the growth. of R. glutinis no. 21 was stimulated in the presence of precipitate of barium sulfate. These findings suggest a connection between growth stimulation and incorporation of barium into the cells. The cells were grown in the medium supplemented with each metal ion at the optimum concentration shown in Tables 2-4 and 2-6.. After harvesting the cells, the. protoplasts were prepared to measure metals incorporated into the cells. detected at 18 ppm in the Ba. 2+. Barium was. -grown cell protoplasts (Table 2-7), indicating that. barium and three other metals as well were incorporated from the medium into R.. glutinis no. 21 cells.. It is still unknown how R. glutinis no. 21 incorporates barium. into the cells from the precipitate of barium sulfate. 15. Francis and Dodge (8) reported.
(19) that the precipitate of metal oxides was incorporated into the cells by direct actions of the microorganisms such as phagocytosis or after degrading the precipitate by catalysis of certain enzymes exorcised from the microorganisms. Table 2-7. Concentrations of metals in the protoplasts of R. g lutinis no.21 Concentration (ppm) Mgz+ Caz+. Metal salt supplemented FeS04 "7H zO. Fe z+ 12.6. 0.4. 1.8. 0.0. MgS04"7H zO. 0.0. 7.8. 0.8. 0.0. CaClz "2H zO. 0.0. 1.4. 40.0. 0.0. BaCl z "2H zO. 0.0. 0.4. 1.5. 18.0. Control. 0.0. 0.4. 0.9. 0.0. Ba2+. Protoplasts (l.5 X 1Q6 cells) were burst m l.Oml of delOlllzed water.. 4) The actions or barium in R. glutinis no. 21 cells Barium is an alkaline earth metal having chemical properties similar to that of calcium or strontium but is generally more stable.. The action of barium in organisms. is known as that of a replacement for other metals having an ion radius similar to that of barium (0.143 nm or 0.136 nm).. For example, strontium ion (ion radius: 0.116 nm). can replace calcium ion (ion radius: 0.0999 nm) in 25 % of cases requiring calcium (9). It was reported that certain metals in some enzymes produced by a microorganism. could be replaced with bariums (10-12).. Based on this, it can be assumed that a. percentage of some metals in the enzyme system related to cell growth was replaced by barium and incorporated into the cells.. The growth enhancement of R. glutinis no.. 21 in LBB by addition of barium salts may have been due to substitution of some other metal by barium, since R. glutinis no. 21 grows under such adverse growth conditions that lactic acid, an antibacterial agent, is the sole carbon source in the medium.. That. barium is an essential metal in the assimilation system of lactic acid in R. glutinis no. 21 is also suggested by the results. These findings seem to throw some light in resolving the intracellular action of barium, a very small amount of which was detected, but which does very important work in the growth of R. glutinis no. 21 cells.. 16.
(20) MATERIALS AND METHODS in Chapter 2 1) Culture conditions. In the growth studies, 500 ml Erlenmeyer flasks were used; Each flask contained 100 ml of lactic acid medium (Table 2-8).. The yeast cultures were preincubated at. 28 DC with reciprocal shaking (120 strokes/min, 5 cm span) until they reach a stationary phase.. After precultivation, the cells were collected by centrifugation. (3,000rpm, 10 min) and washed with physiological saline. for various experiments,. For inoculation to media. pre-cultures were diluted and adjusted to 0.1 units of optical. density at 610 nm (OD 610) with fresh broth.. Then, a portion of 4 ml of diluted pre-. culture was inoculated into fresh broth and cultivated with reciprocal shaking (120 strokes/min, 5 cm span). number of viable cells.. Cell growth was monitored by measuring OD 610 and/or For experiments on growth/pH dependency,. medium pH. was maintained with 10 % Clark-lubs buffer (13).. Table 2-8. Composition of lactic acid medium. Lactic acid (DL) (NH 4hS04 K 2 HP 04 MgS0 4 "7H 2 0 FeS04"7HzO Distilled water. 3.0g 2.0g 1.0g O.05g O.OOlg lOOOml. In the experiments which evaluated effect of metal ion on the yeast growth, lactic acid basal broth (LBB; Table 2-9) or glucose basal broth (GBB; Table 2-10) was used. In the experiments in which cells were grown in the media supplemented with various lactates, each lactate was added into LBB instead of lactic acid at the same molar concentration as 0.3 % lactic acid (33.3 mM) after which pH was adjusted to 3.0 with 1 N NaOH or 1 N HCI.. The pH was maintained constant by addition of Clark-Lubb's. buffer to each medium up to 10 %.. Media supplemented with metal salts were. prepared by addition of each metal salt to LBB or GBB at the predetermined concentrations.. 17.
(21) Table 2-9. Composition of lactic acid basal broth (LBB) Lactic acid (DL) (NH 4)2 S 04. 5.6g 2.0g 1.0g O.Olg 1000ml. K 2HP 04. FeS0 4 '7H 2 0 Distilled water. Table 2-10. Composition of glucose basal broth (GBB) Glucose (NH 4)2 S 04. 20.0g 2.0g 1.0g O.Olg 1000ml. K 2HP0 4. FeS0 4 '7H 2 0 Distilled water. 2) Effects of lactic acid isomers on the yeast growth Lactic acids of DL-, D- and L-form were added at various combinations to the mineral broth.. The medium pH was adjusted to 3.5.. 28°C with reciprocal shaking.. Cultivation was carried out at. Precultivation conditions and inoculums size of. precultures were the same as culture conditions. Specific growth rate was shown by doubling time (DT) calculated from OD values in terms of exponential growth.. And the growth yield was shown by the maximum. cell number (MCN) in the stationary growth phase.. 3) Determination of amount of lactic acid A portion of the cultures collected at a predetermined time was centrifuged at 3,000 rpm for 10 min.. Supernatant broth was incubated in a water bath at 80 "C for. 15 min in order to inactivate remaining enzymes in the broth.. Nicotinamide adenine. dinucleotide (NAD+) and lactic acid dehydrogenase (LDH) were then added to the heat-treated broth.. After the mixture was incubated at 25°C for 5 min, NADH. produced by the reaction with lactic acid and the additives was measured at absorbance at 340 nm (A 340 ).. The amount of lactic acid (C:[g/LD was calculated by the. following equation (14): C = (V X MWI. E. X d X v X 1000) X.L:1E. Where V =. volume of reaction mixture (L-form;2.24 ml, D-form; 2. 29 ml), MW=molecular 18.
(22) weight of lactic acid (90.1 g/mol), X. E. =absorption coefficient for NADH (6.3 mmot l. em-I), d = thickness of a cell (1.0 em), v = sample amount (0.1 ml),. LJE =. difference in A 340 values of the mixture before and after the reaction had occurred. 4) Measurement of the intracellular concentration of metal ions in yeast cells Cultures at the mid-log phase were collected to prepare protoplastizing cells. Cells were counted in every culture before collection to minimize the difference in the protoplast number among the preparations.. Protoplasts were prepared by the. following procedure: Cells were collected by centrifugation (3,000 rpm for 10 min) and resuspended in 0.2 M phosphate buffer (pH 5.7) supplemented with 0.6 M KCI and 0.1 % EDTA. (washing step).. The suspensions were slowly agitated for 2 min and then centrifuged The washing step was repeated several times (at least 3 times) to. avoid contamination with metals from the media.. Cells were then resuspended in 3. ml or phosphate buffer with 2 % Funcelase (Yakult Co., Ltd.) and incubated at 40°C for 12 h. membrane. m!.. After incubation, protoplasts were obtained by filtration through 1.2 /-Lm Protoplasts were washed on the filter with phosphate buffer of at least 10. They were burst by the addition of 1.0 ml of deionized water and the cell fluid. was collected for the assay by atomic absorption spectrophotometry for metals.. The. control was prepared by the same procedures described above with the same volume of culture media containing no metal salts.. 19.
(23) REFERENCES 1) Tanaka,H. and Yokota, S.: New Food Ind.,16,44-45(1974). 2) Yamamoto,T., Azuma,K., and Yoshii,H.: J.Jpn.Soc.Food Technol., 31, 525-530 (1984). 3) Oliva,R.U. and Hang Y.D.: Appl. Environ.Micobiol.,38,1027-1028(1979). 4) Cassio,F.,Leao,C. and van Uden N,: Appl. Envion, Microbiol.,53,509-513(1987). 5) Rara,S., Totsuka, A., Mizuno, A., and Sanuki,H.: J. Brew. Soc. Jpn., 74, 838-841 (1979). 6) Hara,S. and Shimazaki, T.: J. BrewSoc.Jpn.,76,587-591(1981). 7) Barth,C.A. and de Vrese M.: Kieler Milchw. Forsch. Ber.,36, 155-161(1984). 8) Francis,A.J. and Dodge,C.J.: Appl. Env.Microbiol.,54,1099-1014(1988). 9) Yamasaki,S.: Chem.Biol.,19,148-158(1981). 10) Okawa,Y. and Yamaguchi,T.: BioI. Chem.,40,277-283(1976). 11) Higashi,M.,Ito,F., and Masamune,Y.: J.Gen.AppI.Mirobiol.,26,203-216(1980). 12) Mabrouk,S,S.,Amr,A,S. and Abbel-Fattah,A.F.: Agr.Chem.,40,419-420(1976). 13) Clark,W.M. and Lubs R. A.: J.Bacteriol.,2,1-34(1917). 14) Sasaki,T.: Media Circ1e,31,545-550(1986). in Japanese. 20.
(24) EFFECT OF CULTURE CONDITIONS ON THE CAROTENOIDS BIOSYNTHESIS. 3.1 Introduction When lactic acid is used as a single carbon source, a change is observed in the color tone of R. glutinis no.21 as compared with yeast cells cultured on glucose.. glutinis finally synthesizes two kinds of carotenoids 0), (Fig.3-1).. ~-carotene. R.. and torularhodin. This difference is attributable to a change in carotenoid composition,. suggesting that the carbon source has an influence on the carotenoid production system of this yeast.. In this chapter, at first, carotenoids component after cultivation in lactic. acid and glucose was compared.. In addition, effect of culture conditions on the. carotenoids accumulation was investigated.. As a result of these studies of various. conditions for culture, environmental factors involved in increased production of torularhodin have become clear.. ,8-Carotene. Torularhodin. Fig.3-1. Chemical structure of ~-carotene and torularhodin.. 3.2 Carbon source and culture conditions 1) Carbon source No qualitative difference was observed in the resultant carotenoid regardless of 21.
(25) whether culture was performed with glucose or lactic acid as carbon source, and as main carotenoids in the yeast, there was observed f3-carotene and torularhodin as final biosynthetic products, in addition to their precursors.. When the two final products. were compared in carotenoid content, the ratio of torularhodin increased where culture was performed with lactic acid, its being more than half of the amount of main carotenoids.. No changes were observed in this tendency even when pH was changed. in each medium at the start of culture.. Thus, it was considered that the difference in. carbon source had an influence on the amount of carotenoid (Fig. 3-2).. The pH at the. completion of culture was around 3 in each medium. As its physiological property, this yeast has capability of assimilating some carbon sources (2).. However, in each of the carbon sources including fructose that. was studied in the present experiment, the production of torularhodin tended to increase unlike the case of glucose (Fig. 3-3).. Satisfactory growth was observed with. each carbon source.. "C. ·0. 2.9. pH3.2. IU (). ::T. 3.0. pH7.0. /3 -Carotene CD. en. :J: III r+ r+. ..g j. '0. ::s a. 0. T orularhodin 3.0. pH4.0. CD CD. 0. ""'0t:". r:+. <. o· ::s. III r+. (). ::s. a. 3.1. pH6.8 0. 3 2 Carotenoid content(mg/100 g cell dry weight). Fig.3-2 Effects of glucose and lactic acid on carotenoids biosynthesis of R. glutinis no.21. Lactic acid at 0.5% or glucose at 2.0% was added to the growth medium as a carbon source. The pH was adjusted with 1N H3P04 or K2HP04. Cells were harvested during the stationary ohase in each medium.. 22.
(26) Growth (x108cells) 2.2. Lactic acid Glycerol. 1.7. Pyruvic acid. 1.9. Gluconic acid. 2.0. Fructose. 3.5. Glucose. 3.4. o. 25. 100 75 Ratio of each carotenoids (%) 50. Fig.3-3 Effect of carbon sources on carotenoids biosynthesis of R. glutinis no.21. Each carbon source expects lactic acid and pyruvic acid was used in the growth medium at 2.0%. The latter two carbon sources were added at 0.5%. Cultivation was carried out until the stationary phase in each medium.. 2) Nitrogen Concentration of nitrogen in the medium was changed, and its influence on biosynthesis of carotenoid was studied. increased the amount of carotenoid.. A decrease in the concentration of nitrogen. However, it did not show a tendency of changing. the amount of a specific carotenoid (Fig. 3-4).. 23.
(27) 4. ••. Y- Carotene • Torularhodin • Torulene o /3 - Carotene. 3. I I. • I I. 2 x. o. o 1/ 1. 1/ 4. 1/ 20. 1/ 200. C/N ratio. Fig.3- 4 Effect of CI N ratio on carotenoids biosynthesis of R. glutinis no.21 . Glucose and ammonium sulfate were used as carbon and nitrogen sources, respectively. Cultivation was carried out for 6 days in each medium .. 3.3 Dissolved oxygen. Change in carotenoids from 24 hours before start of aeration was shown in Fig. 35.. As a result of slight stirring up to the start of aeration (72 hours after start of. culture), dissolved oxygen nearly disappeared .. Thereafter, the oxygen concentration. increased with aeration, and it reached 8 ppm 48 hours after start of aeration.. As. regards carotenoid content, scarcely any torulene or torularhodin was observed under the condition of no aeration, and a larger part of the main carotenoids consisted of ycarotene and. ~-carotene .. However, the amount of torulene and torularhodin. increased with start of aeration, and at 48 hours after start of aeration, most of the main carotenoids consisted of the said two types.. 24.
(28) •. 2 ....... t:,-.... --. -.. 10 Growth. Ec.. - - DO. 7.53-. t:(). ,-.... ....... Q). t:. bO. go. -0. 8. t:. Q)3:. ::J. >X. bO. 5. .- 0 0 0 t:.,-. 0. -0 Q). >. B't;n eE 11:1-----. 0. (). .!!2. 4. (5. 0. ..J. 2.5 i:S. ..c: ----~0. fj-Carotene. ~. 0. 0. -24. a. 12. 24. (). bO. C/l C/l. (). Q). <.:). 0. 48. Time after aeration(h) Fig.3-5 Effect of aeration on carotenoids biosynthesis of R. glutinis no.21 . A jar fermenter was used for cultivation. Aeration was started at 0 time by ventilation with 5 I/ min and mixing with 500 rpm . Mixing at a low speed (300 rpm) was carried out for 72 h. until starting the aeration. After mixing for 48 h under low-speed rotation. dissolved oxygen could not be detected in the medium.. The dissolve oxygen and the amount of each carotenoid were compared in detail , and the following were found:. ~-carotene. was produced at a nearly constant rate. regardless of the oxygen concentration, while the tolulene and torularhodin increased oxygen concentration-dependently.. This tendency was remarkable with torulene.. The total of torulene and torularhodin clearly showed a proportional relationship to increased dissolved oxygen (Table 3-1).. Table 3-1. Relationship between carotenoid contents and dissolved oxygen Carotenoid contents (mg/lOOg cell dry weight). DO (ppm ) 0 4.080 8.304. 'Y-Carotene. ~ -Carotene. Torulene. Torularhodin. 0.47 0 .10 0.06. 0.34 0.85 0.17. 0 .06 0.92 1.39. 0 .05 0.48 0.68. 25. Torulene+ Torularhodin 0 .11 1.39 2.07.
(29) 3.4 Discussion There have been many studies about carotenoids produced by yeast of genus. Rhodotoruia (10,11). already been clarified.. As regards R. giutinis, the routes of its biosynthesis have One route is to produce ~-carotene from 'Y-carotene.. Another route is to biologically synthesize torularhodin from 'Y-carotene with torulene as precursor (1).. From various studies, it has been known that. ~-carotene. works not. only as provitamin A, but also, it has various actions, thereby contributing to activities of cells (5).. This specie biologically synthesizes. ~-carotene. and torularhodin.. suggests that torularhodin itself has actions different from those of. This. ~-carotene.. In. this respect, one is tempted to have interest in basic study of torularhodin.. R. giutinis no. 21 was isolated as a yeast viable with lactic acid as carbon source and the yeast cells obtained after culture with lactic acid is slightly reddish as compared with one obtained after culture with glucose as carbon source. carotenoid in each cultured yeast cells were examined.. As a result, the following. were found: When cultured with lactic acid, a decrease was observed in content.. Thus,. ~-carotene. The difference in color tone was attributable to quantitative difference. between two finally obtained carotenoids.. Even when pH was adjusted at the start of. culture, carotenoid composition showed a similar tendency.. At completion of culture,. the pH was found to have decreased to around 3 in each medium regardless of whether pH was adjusted or not at the start of culture.. Therefore, it was suggested that the. change in carotenoid content was brought about by the difference in carbon source. Nitrogen content had an influence on carotenoid content, and a decrease in nitrogen led to an increase in carotenoid content (3). in oil and fat.. Carotenoids are highly soluble. When nitrogen decreases, lipid increases, resulting in a state of easy. accumulation, which is considered to be one of the causes of increased carotenoid content.. In present study, an increase in carotenoid content was observed as reported. previously.. However, no tendency of increase in the amount of a specific carotenoid. was observed.. This suggests that although nitrogen concentration is involved in. carotenoid accumulation, it is not a factor that affects biosynthesis. A study was conducted of the influence of oxygen concentration.. As a result, it. was found that the production of torulene and torularhodin tended to increase with aeration.. A study of relationship to oxygen concentration in the medium revealed. that the. production. of torulene. and. torularhodin 26. clearly. increased. oxygen.
(30) concentration-dependently.. On the other hand, the production of 13-carotene. remained the same regardless of whether there was aeration or not.. This shows that. the production is not induced by the oxygen concentration in the medium.. It is known. that the load of oxidative stress activates removal of active oxygen in the yeast.. With. Saccharomyces cerevisiae, an increase in intracellular glutathione level has been confirmed (4).. On the other hand, carotenoids, such as 13-carotene and astaxanthin,. have capability of removing active oxygen species (5,6), and these protect the living body from disturbances caused by free radicals present in an excessive state.. When. oxygen concentration in R. giutinis no. 21 was increased by aeration, no change was observed in 13-carotene content; however, increases were noted in the production of torulene and torularhodin, and this tendency was remarkable with torulene.. In the. carotenoid biosynthetic system of this yeast, there are two routes as previously mentioned.. One is synthesis of 13-carotene from y-carotene, and the other is. synthesis of torularhodin via torulene.. This is considered to result from activation of. the torularhodin production system due to oxygen concentration. produces active oxygen by various intracellular reactions (7).. Molecular oxygen. In other words, a state. of excessive oxygen brought about by aeration increases intracellular level of active oxygen, and as a result, R. giutinis no. 21 increases intracellular production of torularhodin and not 13-carotene.. From these, a possibility is suggested that in. protection of cells from active oxygen, namely, in action of quenching active oxygen, torularhodin have a stronger action than 13-carotene. will be further studies of torularhodin.. Therefore, it is hoped that there. It already became clear that the change in. color tone observed previously when lactic acid was used as carbon source was attributable to increased ratio of torularhodin in carotenoid biosynthesis. change was observed with some carbon sources.. Similar. In case of lactic acid, it is. considered that in assimilating it, it is converted to pyruvic acid by lactic acid dehydrogenase, and then, metabolism occurs.. However, one of the causes of the. aforesaid change is considered to be that the system of carotenoid biosynthesis responds to the active oxygen (8) produced by the oxidation-reduction reaction of the NADH produced in the dehydrogenation process.. 27.
(31) MATERIALS AND METHODS in Chapter 3 1) Culture condition Cultivation medium was used in the mixture composed of 20.0 g of glucose, 2.0 g of (NH4)2S04' 1.0 g of K 2HP0 4, 0.5 g of MgS0 4·7H 20, 0.01 g of FeS04·7H20 and O.Olg of Peptone in 1,000mL of distilled water (pH 6.8). When adjusting pH, 1 N phosphoric acid or 1 N dipotassium hydrogen phosphate was used.. Culture. conditions were same as mentioned in chapter 2. In studying oxygen concentration, culture was performed (28°C, 500 rpm) in a medium of 5 liters (jar fermentor, 8-liter capacity, MBF-800, EYE LA Co.). Dissolved oxygen (DO) in the medium was measured by a DO measuring equipment (Integral Unit FM-110, EYELA Co.). by aeration and stirring.. Dissolved oxygen in the medium was adjusted. In order to remove the influence of existing oxygen at the. time of adjusting the medium, culture was performed with only slow speed stirring (300 rpm) up to 72 hours after start of the culture, and thereafter, aeration (5 liters/min, stirring speed, 500 rpm) was given. 2) Isolation and quantitative determination of carotenoid Pellet of cultured cells was lyophilized, after which extraction was performed in a mortar with benzene, chloroform and methanol in order, and then, the resultant product was dissolved again in benzene after evaporation of previously used solvents.. After. that, the product was dehydrated with addition of anhydrous sodium sulfate. The substance obtained was used as sample of carotenoid.. Torularhodin was isolated by. column chromatography (Wakogel C-100, Wako Pure Chemical Industries, Ltd.) with benzene:acetone = 4: 1 (TLC on silica gel 60 was shown in Fig.3-6).. ~-Carotene, y-. carotene and torulene were isolated by high performance liquid chromatography (Waters 486, Column. Cosmosil 5SL, flow rate 1.0 ml/min; elute, A=100% hexane, B=20% ethyl acetate in hexane, A. ~. chromatogram was shown in Fig. 3-7).. B 30min linear graduate; detection, 460 nm, Quantitative determination (mgllOOg dry cell. weight) was made using Mcbeth (9-11) formula.. 28.
(32) Carotenoid (mg/lOOg cell dry weight) =. OD. ~DXVXl.03. e x weight of tIssue(g). (McBeth formula). : ""max. V : solvent volume E. :. Absorption coefficient (Table 3-2). Table 3-4. Absorption coefficient of McBeth formula Carotenoid E 2337 p-Carotene 3100 'Y-Carotene 3240 Torulene 1932 Torularhodin. 29. Solvent Benzene Light petroleum Light petroleum Chloroform.
(33) .........._._-f---. .............- - - t - -. Fig.3-6. R f A. 1. B. a. 4. c. o. TLC on siligagel 60 of carotenoids from R. glutinis no.21. Elute: 20% ethyl acetate in benzene,. 30. Fraction B: torularhodin.
(34) @ @. 5. 10 Fig.3-7. 20 min. 15. HPLC pattern of fraction A. Conditions: Waters 486, Column. Cosmosil 5SL, flow rate 1.0 mllmin; elute, A=100% hexane , B=20% ethyl acetate in hexane, ADB 30min linear graduate; detection, 460 nm, Fraction 1, f3-carotene; Fraction 2, f3-zeacarotene; Fraction 3, y-carotene; Fraction 4, torulene.. 31.
(35) REFERENCES 1) Simpson, K.L., Nakayama, T.O.M., and Chichester,. c.o.: Biosynthesis of yeast. carotenoids. J . Bacteriol. ,88, 1688-1694(1964). 2) Fell, J.W., Tallman, A.S., and Ahearn, D.G.: The Yeast,(Kreger-van Rij,N.J.W), 3rd ed., 893-894,Elsevier Science Publishers B.W.,Amsterdam(1984). 3) Yamaoka, T., Takimura, 0., Fuse, H., Uemura, K., Manabe, E., Takano, H., and Hirano, M., J. Fermen. Tech., 70,25-28(1992). 4) Inoue, Y., Kyoto Univ. Food Chern. Rep.,59,1-3(1996). 5) Ito, T., Vitamin,66,137-146(1992). in Japanese 6) Miki, W.: Biological functions and activities of animal carotenoids. Pure Appl. Chern., 63, 141-146 (1991). 7) Asada, K., Biochem., 48,226-257(1976). in Japanese 8) Asada, K., Metabolism, 17,1705-1718(1980). 9) McBeth, J.W.: Carotenoids from nudibranchs. Comp.Biochem.Physiol., 41,5568(1972). 10)Bonner, J., Sandroval, A., Tung, T.W., and Zechmister, L.,Arch.Biochem, 10,113123(1946). l1)Villoutrein, L., J.Biochem.Biophys.Acta., 40,442-457(1960).. 32.
(36) ACTIVATION OF TORULARHODIN PRODUCTIVITY AND ENHANCEMENT OF TOLERANCE. 4.1 Introduction An yeast R. glutinis no.21 varies the production rates of the two carotenoids depending on the incubation conditions, and production of torularhodin is induced by an oxidative stress.. These findings arose the assumption that R. glutinis no. 21. increases its production of torularhodin for protection from membrane impairment by activated oxygen molecules, suggesting that torularhodin may possibly have a potent anti-oxidation property.. /3-Carotene was subjected to epidemic researches in yeast (1).. In addition, physiological activity of carotenoids including /3-carotene became attractively valuable (2-4), causing a further demand of the studies on torularhodin. To know the characteristics of torularhodin, a mutant that accumulates torularhodin was made. In this chapter, a mutant which produces increased amount of torularhodin from stock strain R. glutinis no. 21 was characterized the carotenoid biosynthesis by a resultant mutant together with the incubation characteristics based on the increase in the production of torularhodin.. In the study of characterization of mutant incubated. with the oxidative stress, methylene blue was used for generator of active oxygen species, and cell damage was evaluated with the guidance of the leakage of LDH into the cultivation medium.. 4.2 Effect of active oxygen species generators Effect of the MB, MV and AAPH on the carotenoid biosynthesis was shown in Fig. 4-1.. All the generators enhanced the carotenoid biosynthesis, above all, MV. indicated the remarkable effectiveness.. However, each generator did not enhance the. productivity of /3-carotene, whereas each active oxygen species and radical enhanced that of torularhodin and torulene, precursor of torularhodin.. Torulene production was. increased up to about two-fold, and 2- to 2.5-fold increase was observed for torularhodin production, irrespective of the types of generators. 33. Torulene and.
(37) torularhodin were produced in the quantity of 1.5 mg/l00 g cell dry weight in total, which was approximately double that produced in the non-added group (0.75 mg/lOO g cell dry weight).. The result showed that oxidative loading activates the pathway for. torularhodin production.. AAPH. Methyl viologen. Methylene blue. Control. o. 2.5 0.5 1 1.5 2 Amount of carotenoids (mg/100g cell dry weight). Fig. 4-2 Effect of active oxygen species on the carotenoids biosynthesis in R. glutinis no.21. MB (1.0 x 10-7 M), MV (1.0 x 1Q-l0 M) and AAPH (3.0 x 10-6 M) were used as generators of singlet oxygen, superoxide anion radical and peroxy radical, respectively. Yeast was harvested after growth was reached stationary phase. 0: /3-Carotene, . : Torularhodin, [i): Tolurene. Investigation of the effect of continuous MB supply on carotenoid biosynthesis (Fig. 4-2) revealed the increased production of torularhodin.. This increase is. apparently greater than that produced by single addition at the onset of culture, indicating that oxidative load gives rise to increased production of torularhodin.. 34.
(38) 24. 48. 72. 96. Control. Fig.. 4-3 Change of carotenoids component by the continuous addition of methylene blue. MB, prepared at concentration of 1 X 10-5 M, was continuously added by rate of 10ml per 1DOOml medium once or more a day. 0: p-Carotene, . : Torularhodin, LillI: Tolurene. 4.3 Tolerance against oxygen stress of torularhodin higher production mutant After irradiation of an ultraviolet (254nm, 68 f.1W/cm2 and 65cm distance), a more reddish colony, which exhibited a color tone different markedly from that of a parent strain, was obtained.. Microbiological identity of this mutant was investigated based. on the morphological, physiological and fermentation characteristics, and no difference from those of the parent strain (see chapter 1) was observed so far as examined.. This mutant was designated as mutant TL121 and subjected to the. following experiments.. To compare behavior on oxidative stress with TLI21, a. mutant that increases in the production of p-carotene was concomitantly obtained and named mutant C. The compositions of the carotenoids produced by the mutants obtained are shown in Fig.4-3.. When compared with the parent strain, a marked increase in the. production of torularhodin was observed in the mutant TLl21.. While torularhodin. was produced in an amount larger by about 4 times than that produced by the parent 35.
(39) strain, no substantial difference in the production of ~-carotene was observed.. With. regard to the variation by the wavelengths of the lights irradiated, no increase in the production of torularhodin was observed with an red or green light, but the incubation under a blue light caused the increase in the production of torularhodin.. The parent. strain exhibited no change in carotenoid composition in response to the irradiation with the blue light.. Thus, the mutant has a capability of varying the production of. torularhodin depending on the lights and that this capability is induced by the irradiation with a blue light.. A mutant C produced. about 7 times than parent strain.. ~-carotene. in an amount larger by. Total amount of carotenoids in this mutant was less. larger than mutant TLl21 (ratio of 0.7).. However this mutant showed carotenoid. content significantly different from that of parent strain (R. giutinis no. 21).. The responses of mutant TLl21 and other strains to different MB concentrations. m1Torulene • Torularhodin o (3 -Carotene. I. I I. I. "'0 Q). c:::. Mutant TL/21. Parent strain. Mut C. Fig.4-4 Biosynthesis of carotenoids under different wavelengths of light irradiation. Wavelengths supplied from color films were as follows. Blue: 360-495nm, Green: 500-583nm, Red: 61 Onm infrared light. Mut C is a high-p-Carotene-producing mutant. In this mutant, no torulene production was detected.. are shown in Table 4-1.. Incubation with MB at 0.1 mM under the dark condition. depressed the yeast growth in the all strains, but more depression was noted with irradiation of white light in the parent strain.. Singlet oxygen generated by methylene. blue with white light illumination was indicated results from these control data. 36. In.
(40) the parent strain, the proliferation in the presence of MB at 0.01 mM or higher was inhibited, and the cell number at 0.1 mM was 23 % based on the growth in the absence of MB.. On the other hand, mutant TL121 exhibited a satisfactory proliferation. equivalent to the growth in the absence of MB, although addition of MB at O.lmM prolonged the log phase of yeast growth.. In the mutant TLI21, torularhodin was. observed to increase with irradiation of white light.. Mutant C indicated tolerance to. MB, but its degree was not so strong as mutant TLl21. To measure a LDH activity, MB was added to a concentration up to 1.0 mM after incubation until the stationary phase to obtain sufficient amount of LDH from injured yeast.. LDH activity after 24 hours from addition of MB was compared with initial. value.. The parent exhibited an 84 % increase in the activity at 0.5 mM and an almost. similar increase in the activity even at 1.0 mM.. In mutant TLl21, the activity was. increased at 0.5 mM only by 35 %, and only by 54 % even at 1.0 mM (Fig.4-4).. LDH. in the medium incubated with mutant C was much than it from mutant TL121 at 0.5 and 1.0 mM of MB.. It is well known that LDH leak out of damaged cells.. released from microbiological organism injured by oxidative stress (5).. It was also. Accordingly,. it was suggested that the torularhodin-producing mutant had the resistance to an oxidative stress by the addition of MB.. The low LDH activity also suggested that the. resistance be obtained as a result of prevention of a membrane injury.. A parent yeast,. R. glutinis no. 21, was proven to increase the production of torularhodin when incubated under an oxidative load by aeration.. These findings are considered to. reflect a priority of torularhodin synthesis under an oxidative load in a test strain undergoing biosynthesis of J3-carotene and torularhodin.. A torularhodin production-. increasing mutant obtained in this study acquired the resistance to MB-derived singlet oxygen evidently when compared with the incubation characteristics of the parent strain.. Based on the results obtained here, torularhodin may contribute, at least to. some extent, to the protection from an oxidative impairment. known to be associated with J3-carotene (3).. The similar effects are. R. glutinis no. 21 produces not only J3-. carotene but also torularhodin, and this fact is considered to be important in understanding the significance of the production of torularhodin. obtained in this study will serve greatly in investigating torularhodin.. 37. The mutant.
(41) 100 90 80 ,.-.. ~ -....; 70 I 60 0 ...I 50 40 40 Q) be 30 III .:t:. III 20 Q) ...I 10 0 -10. O.1mM. O.5mM. 1.0mM. Methylen blue concentration Fig.4-5 Changes in LDH activity under different concentration of methylene blue. Methylene blue was added to the GP medium diluted to one tenth after the cells reached the stationary phase. The cells were irradiated with white light during the incubation. . : Parent strain; 0 : Mutant TL/21; ~ : Mut C, high-~ Carotene-producing mutant.. Table 3-1. Influence of Methylene blue (MB) on the growth of mutant TLl21. Torularhodin Cmg/lOOg cell dry weight) Control 1.5 4.02±0.07 2.0 O.OlmM 3.SS±0.lS 0.S9 O.OSmM 2.3 Parent Strain 1.09±0.11 0.27 O.lmM 2.1 0.91±0.20 0.23 O.lmMCDark) 0.7 0.44 1.76±0.30 Control 4.3 3.29±0.16 O.OlmM 4.9 3.S4±0.26 LOS O.OSmM 4.S TL121 3.1S±0.14 0.96 4.4 O.lmM 3.44±0.24 LOS O.lmMCDark) 1.9 0.S6 1.S4±0.02 Control O.S 2.84±0.11 O.OlmM 1.1 2.70±0.06 0.9S O.OSmM 1.3 Mutant C 2.01±0.26 0.71 O.lmM 1.5 2.02±0.19 0.71 O.lmMCDark) 0.6 0.46 1.32±0.10 Each stram was mcubated under lfradlatlOn wIth whIte lIght m GP medIUm dIluted to one tenth. The growth rate is represented as the inhibition rate of growth with respect to the number of cells grown in methylene-bluefree medium. Yeast growth was monitored by measuring OD at 610nm, and after reaching the stationary phase, the number of cells was counted using Thoma's plate. The number of cells is represented as the mean ± S.D. of three data. Strain. Concentration of MB. Number of cells. 38. Rate.
(42) MATERIALS AND METHODS in Chapter 4. 1) Generators of active oxygen species and radicals Methelene blue (MB), methylviologen (MV) and 2,2-azo-bis (2- amidinopropane) -dehyrochloride (AAPH) were used as generators of singlet oxygen, superoxide anion and peroxy radical, respectively.. For 5 days culture broth was incubated with 1.0 X. 10-7 M of MB, 1.0 X 10- 10 M of MV and 3.0 X 10-6 of AAPH and without any generators.. From the result of a preliminary examination, the highest concentration. from which growth was not inhibited was decided as a concentration of addition to a culture medium.. When the influence of intermittent exposure was evaluated,. generator was continuously added to the culture medium once or more per day. 2) Preparation of mutants and its microbiological properties Mutant was produced using a procedure as described previously (6,7).. The. conditions of ultraviolet rays irradiated were as follows, 254nm, 68 JlW /cm 2 and 65cm distance. For characterization of mutant, glucose pepton broth (NIHONSEIYAKU Co. ,Ltd., GP medium) were used.. A 500-ml flask containing 100 ml of GP medium was. inoculated with the yeast cells washed with phosphate buffer saline(pH 5.6) and incubated for 3 to 5 days at 30 e with reciprocal shaking at 120 rpm with 5 cm span. 0. Fluorescent lamp was used for illumination and wavelength was controlled with spectrum color filters.. Intensity of penetrable illumination was adjusted to 150 lxs.. In the examination to tolerance for active oxygen species, methylene blue (MB, color index number: 52015, Wako Co., Ltd.), as the origin of singlet oxygen, was added from O.OlmM to O.lmM in GP medium diluted to one tenth.. Cell number was counted by. Thoma plate after reaching stationary phase, which was confirmed by monitoring optical density at 610nm.. A tolerance in each strain was represented as inhibition. rate (%) based on the cell number in the absence of MB.. 3) Assay of lactate dehydrogenase When lactate dehydrogenase (LDH) activity was measured, MB was added to diluted medium from O.lmM to 1.0mM.. LDH activity in the cultivation medium was. determined before and after addition of MB according to the procedure described 39.
(43) previously (8).. A concrete procedure is as follows.. A portion of the cultures. collected at a predetermined time was centrifuged at 3,000 rpm for 10 min. Supernatant broth (0.05 ml) was incubated in a water bath at 37°C for 10 min with 0.5 ml of buffer solution including. ~-nicotinamide. adenine dinucleotide (NAD+: 0.65. !lmol) and nitroterazolium blue (N0 2 _TB: 0.125 !lmol) as color and lithium lactate as reaction substrate. nm. of reaction. spectrophotometer.. After mixture with 0.1 N hydrochloride acid, absorbance at 560 mixture. (attributed. with. diformazan). was. measured. using. The activity of LDH present in culture broth was determined by. reference to a calibration curve prepared in advance.. Microbiological properties of. mutants, examined according to the procedure described previously (9), were investigated by comparing morphological, biological and cultivating properties with R. giutinis no. 21.. 40.
(44) REFERENCES 1) Moore, M.M., Breedveld, W., and Autor, A.P.: The role of carotenoids in preventing oxidative damage in the pigmented yeast, Rhodotorula mucilaginosa. Arch. Biochem. Biophys., 270,419-431 (1989). 2) Miki, W.: Biological functions and activities of animal carotenoids. Pure Appi. Chern., 63, 141-146 (1991). 3) Terao, J.: Antioxidant activity of /3-carotene-related carotenoids in solution. Lipids, 24, 659-661 (1989). 4) Murakoshi, M., Nishino, H., Satomi, Y., Takayasu, J., Hasegawa, T., Tokuda, H., Iwashima, A., Okuzumi, J., Okabe, H., Kitano. H., and Iwasaki, R.: Potent preventing action of a-carotene. against carsinogenetis:. Spontaneous liver. carcinogenesis and promoting stage of lung and skin carcinogenesis in mice are suppressed more effectively by a-carotene than by. ~-carotene.. Cancer Res., 52,. 6583-6587 (1992). 5). Pizarro R.A.: UV-A oxidative damage modified by environmental conditions in Escherichia coli. Int. J. Radiat. BioI., 68, 293-299(1995).. 6) Ouchi, K., Shimada, M., Nakamura, Y., Kojima, Y., and Nishiya, T.: Isolation of auxotrophic mutants from industrial yeasts. Hakkokogaku., 61, 349-352 (1983). 7) Sato, S., Otani, M., Shimoi, H., Saito, K., and Tadenuma, M.: Selection of flocculent mutants of yeasts for wastewater treatment. J. Brew. Soc. Japan, 82, 515-519(1986). 8) Babson, A.L., and Pillips, G.E.: A rapid colorimetric assay for serum lactic dehydrogenase. Clin. Chim. Acta., 12, 210-215 (1965). 9) Fell, J.W., Tallman, A.S., and Ahearn, D.G.: The Yeast,(Kreger-van Rij,N.J.W), 3rd ed., 893-894,Elsevier Science Publishers B.W.,Amsterdam(1984).. 41.
(45) SCAVENGING OF PEROXYL RADICALS BY TORULARHODIN. 5.1 Introduction It has been suggested that R. glutinis, a soil yeast that accumulates torularhodin. and ~-carotene as final products, regulates biosynthesis of carotenoids by increasing production of torularhodin based on increase in dissolved oxygen of cultivative medium.. In addition, a mutant strain with increased torularhodin productivity. exhibited resistance against oxygen loading caused by addition of methylene blue, a singlet oxygen generator, to culture medium and proliferated better than the mother strain or another mutant with increased. ~-carotene. productivity.. These results. suggested that R. glutinis produced more torularhodin against oxygen loading than carotene.. ~. It can therefore be presumed that torularhodin defenses against active. oxygen species in yeast cells. In this chapter, carotenoid composition after the addition of peroxyl radicals generator to culture medium was analyzed.. In addition, peroxyl radicals, main chain. carrying species in lipid peroxidation, scavenging activity of torularhodin was examined.. Inhibitory effect on lipid peroxidation was also compared with that of. ~. carotene, which was reported to be a strong peroxyl radical scavenger (1-3).. 5.2 Relation of growth inhibition and carotenoid content The growth of R. glutinis in AAPH was similar to that in the control group up to a concentration of 0.1 mM, but low cell density at 48 hours after inoculation was observed at 0.5 mM or higher (Fig. 5-1). noted at 1 mM.. And remarkable growth inhibition was. When AAPH was added to the culture medium at the concentration of. 0.01 mM, the amount of carotenoids in yeast was increased. AAPH addition,. torularhodin was. With increase in load by. consumed faster than. ~-carotene.. After. torularhodin decreased to the same concentration as AAPH free condition, f3-carotene began to decrease.. 42.
(46) 7.0 6.0 ':;). .... :§ 5.0 5i ~. 1:: ~. 8. 4.0. "0. "'C~. ·0. 5i. 8 3.0. e8. bO. "'~. 0. 1. 2 .0 1.0 0.0. o. 0.01. 0.05. 0.1. 0.5. Concentration(mM). Fig.5-1 Growth of R. g/utinis no.21 in medium including MPH as a peroxyl radical generator and change in content of major carotenoids. MPH was added to glucose pepton medium diluted to onetenth. The yeast cells were harvested at 72 hours after inoculation, washed and then disruped in a supersonic generator. 0: b-Carotene, . : Torularhodin, III :Tolurene. 5.3 Peroxyl radicals scavenging The radical scavenging activities of each compound obtained at different concentrations in expo 1 are shown in Fig. 5-2.. Torularhodin in the range of 0.0025 to. 2.5 mM exhibited concentration-dependent scavenging activity and it reached to 60 % ~-carotene,. at 2.5 mM.. In respect of. at 2.5 mM.. The activity of torularhodin was stronger than that of. concentrations tested.. the elevation of the activity was observed only ~-carotene. at all. On the other hand, a-tocopherol exhibited concentration-. dependent scavenging activity up to 0.25 mM, and its activity was clearly stronger than that of. ~-carotene. or torularhodin.. However, at concentration above 0.25 mM, the. activity of a-tocopherol was decreased, and it was weaker than that of torularhodin at 2.5mM. 43.
(47) In expo 2 (Fig. 5-3), a-tocopherol exhibited concentration-dependent scavenging activity.. At concentrations above 0.25 mM, scavenging activity of torularhodin was. clearly elevated.. However, a-tocopherol exhibited stronger scavenging activity than Scavenging by ~-carotene against peroxyl. did torularhodin at the same concentration.. radicals was not found in the tested concentration range in expo 2. The results of these two experiments indicated that torularhodin exhibited peroxyl radicals scavenging activity regardless of the generating system used, and its activity was stronger than that of a-tocopherol at high concentration particularly in expo 1.. 70. ~ 50 ....... .~ .~. (). '" 30. !. '6iJ. c ~. ~. 10. o 0.0025. 0.025. 0.25. Concentration. (mM). 2.5. Fig.5-2 Scavenging activity against peroxyl radical generated from CHP-TPP' Fe(m) reaction in DMSO. O. torularhodin; D. /3 -carotene; •. vitamin E.. 44.
(48) 70. ....... ~ ........ .~>. '13 I'll l!!' '611 ii. 50. 30. >. ~. 10 0 0.0025. 0.025. 0.25. 2.5. Concentration (rrM). Fig.5-3 Scavenging activity aginst peroxyl radical generated from AIBN pyrolysis in DMSO. 0. torularhodin; D. /3 -carotene; •. vitamin E.. 5.4 Inhibitory effect on lipid peroxide formation Each. compound. exhibited. concentration-dependent. peroxidation in the range of 0.1-10 J.lM (Table 5-1).. inhibition. of. lipid. Of the compounds tested,. torularhodin exhibited the strongest inhibition, and its inhibitory activity was stronger than those of a-tocopherol and /3-carotene at concentrations above 1 J.lM. no significant difference was observed among three compounds.. At 0.1 J.lM,. At 1 J.lM or lower,. /3-carotene had almost the same inhibitory activity as a-tocopherol, whereas at 10 J.lM its activity was weaker than that of a-tocopherol.. Of the three compounds tested in. this study, the inhibitory activity of /3-carotene was weakest.. 45.
(49) Table 5-1 Effect of torularhodin on formation of MDA caused by lipid peroxidation of rat brain homogenate MDA (nmol/ml of homogenate) Inhibition Concentration Compound (% of control) Mean Standard Deviation M (Il ) 0.1 28.0 3.9 36.8 55.3 Torularhodin 19.8 6.2 1.0 10.8 75.6 10 3.3 32.1 27.5 0.1 8.6 36.1 28.3 0,- Tocopherol 1.0 5.4 63.7 10 16.4 3.7 32.7 26.2 0.1 5.1 30.0 32.3 1.0 6.1 ~-Carotene 49.0 10 22.6 4.1 Control (DMSO) 44.3 10.9 Mean and standard deviation was calculated from 3 data. MDA: Malonyldialdehyde. 5.5 Discussion R. glutinis, a yeast accumulates torularhodin abundantly, varied system of carotenoid biosynthesis and increased torularhodin productivity by oxygen stress. These findings suggested that torularhodin plays a more important role in defensive against oxygen stress than other carotenoids in yeast. properties of torularhodin were compared with. Hence, the anti oxidative. ~-carotene.. And author sought to. estimate the biological function in yeast of torularhodin. Peroxyl radical is most dominants radical in the process of lipid peroxide formation.. And the chain reaction ends with formation of non-radicals owing to. collision of peroxyl radicals with one another or scavenging by anti-oxidants. Therefore, evaluation of the potency of scavenging of peroxyl radicals is very important in evaluation of lipid oxidation suppression. The total carotenoids increased at the concentration of the AAPH, peroxyl radical generator, with which yeast growth is not inhibited. torularhodin than. ~-carotene. R. glutinis consumed. with increase in loading by AAPH.. declared that peroxyl radicals are main factor.. But it cannot be. Therefore, author examine scavenging. of peroxyl radicals by carotenoids biosynthesized by R. glutinis.. Although. scavenging activity was not measured directly so far, Namikawa et al developed a novel procedure using ESR for detection of peroxyl radicals generated by the CHPTPP-Fe(ill) reaction and pyrolysis of AIBN (13). 46. This has enabled determination of.
(50) the peroxyl radicals scavenging activity of highly lipophilic antioxidants.. In this. method of direct measurement, torularhodin exhibited scavenging activity equivalent or superior to that of a-tocopherol, although the activity measured differed depending on the peroxyl radicals generator system used.. The activity of torularhodin never. surpassed that of a-tocopherol at any concentration in the system with pyrolysis of AIBN.. This may be attributed to the heat stability of carotenoids.. In the reaction of. CHP and the TTP-Fe (III) system, activity of a-tocopherol decreases at high concentration.. It was reported that a-tocopherol became proxidant at high. concentration (4), and result obtained may support that. scavenging of peroxyl radicals by affected this (5).. ~-carotene,. Scavenging activity of. There are a few reports on. and partial oxygen pressure strongly. ~-carotene. was not detected under the. present experimental conditions. When torularhodin was added to rat brain homogenate, concentration-dependent suppression of MDA formation was observed, and the effect of torularhodin was stronger than those of a-tocopherol and peroxyl radicals by. ~-carotene. ~-carotene.. It is thought that scavenging of. is the main cause of suppression of the lipid. peroxidation (5), and strong activity is related to stabilizing of hydroperoxide in atocopherol (6).. Torularhodin showed strong activity compared with these compounds. against lipid peroxide formation.. The fact that torularhodin inhibited peroxide ~-carotene. formation more effectively than a-tocopherol or. is very interesting,. considering the deep involvement of peroxyl radicals in the chain reaction of lipid peroxidation (7). Although torularhodin exhibited potency in scavenging peroxyl radicals similar to that of a-tocopherol, it exhibited marked difference from it in suppression of lipid peroxide formation in rat brain homogenate.. This finding suggested that torularhodin. may contribute to the suppression of peroxide formation by an activity in addition to scavenging of peroxyl radicals.. To explain to above results, more examination is. needed. It has been demonstrated that antioxidative activities are due to capturing or. scavenging of active oxygen species by carotenoids, and their relation to structure has been discussed (8-10).. There are two pathways of biosynthesis of carotenoids by. yeast in Rhodotorula sp, which are known to produce carotenoids. carotene to. ~-carotene. torularhodin. accompanying cyclization of. formation. through. desaturation. ~-ionone. and. One is from y-. ring, and the other is to. carboxylation. (11).. Since. torularhodin is not cyclized in the process of biosynthesis from y-carotene, the carbon 47.
(51) chain that contributes to the stability of radicals appears to be long. be presumed that the difference in activity between torularhodin and. It can therefore ~-carotene. was. related to the difference between them in length of carbon chain. R. glutinis increased production of torularhodin upon excessive aeration of culture. medium.. In addition, a high-torularhodin-producing mutant (TL12l). resistance to oxygen stress higher than another strain.. showed. Lactate dehydrogenase activity. in medium of low resistance strain was high as compared with that of medium cultured with TL121.. This indicated that the yeast membrane has received the damage by. oxygen stress.. Aeration and singlet oxygen generated by methylene blue addition are. considered to be an initiator on the process of damage of yeast cells.. It is reasonable. to assume that a dominant factor is peroxyl radicals because cell membrane is rich in lipid.. Although it is important to remove initiator to prevent growth inhibition of. yeast, the deletion of peroxyl radicals becomes important in the stop of the chain reaction begun.. As a result of examination which used ESR, torularhodin was found. to be a strong scavenger of peroxyl radicals than ~-carotene at all concentrations tested. It thus appears that torularhodin was biosynthesized to preserve yeast from oxygen. stress.. Further, the results that cannot be explained only by difference of the. scavenging activity of peroxyl radicals was obtained from experiment of lipid peroxide formation.. This result suggested that torularhodin have another effective activity or. biological property such as affinity for the membrane. In conclusion, torularhodin is an effective scavenger against peroxyl radicals, and appears to play an important role in protecting against injury by oxygen stress in R. glutinis.. Further investigation of this carotenoid is expected.. 48.
(52) MATERIALS AND METHODS in Chapter 5 1) Culture condition A glucose peptone broth (Nihonseiyaku Co. Ltd., Tokyo, GP medium) was used for yeast cultivation.. A 500-ml flask containing 100 ml of GP medium was. inoculated with yeast cells washed with phosphate-buffered saline (pH 5.6) and then incubated for 3 to 5 days at 30°C with reciprocal shaking at 120 rpm with a 5 cm span. 2,2'-Azobis(2-amidinopropane)-dihyrochloride (AAPH, Wako Co. Ltd., Osaka) was added as a generator of peroxyl radicals at various concentrations from O.OlmM to 1mM to the GP medium.. Yeast growth was monitored by optical density at 610nm.. Carotenoids were isolated and analyzed quantitatively as described previously (9). 2) Electron spin resonance (ESR) spectrometry Two reaction systems were used for generation of peroxyl radicals (15).. The. first system used cumene hydroperoxide (CHP) with 5,10,15,20-tetraphenyl-21 H,23 H-prophine iron(III) chloride (TPP-Fe(III)) (exp. 1).. Fifty III of 0.1 mM TPP-Fe(III),. 50111 of 0.2 mM 5,5'-dimethyl-1-pyrroline N-oxide (DMPO) as a spin trap reagent, 50. III of test substances dissolved in dimethyl sulfoxide (DMSO) and O.4mM CHP were placed in a test tube and mixed using an automatic mixer.. Exactly 7 minute after, the. spin adducts formed by the reaction of peroxyl radicals were recorded by ESR spectrometry (JES FR-30, JOEL, Tokyo, Japan). 2,2'-azobisisobutyronnitrile (AIBN) (exp. 2).. The second system used pyrolysis of Fifty III of 0.2 M AIBN, 50111 of 0.2 M. N-tert-butyl-a-phenylnitrone (PBN) as a spin trap reagent, and 50 III of teat substances dissolved in DMSO were placed in a test tube and mixed using an automatic mixer. After heating at 50°C for 10 min, recording of spin adducts was started using ESR spectrometry.. Conditions for ESR spectrometry were as follows: magnetic field,. 335.5 mT; power, 8.0 mW; response time, 0.10 sec; modulation, 0.2 mT; temperature, 298 K; sweep time, 1.0 min.. Manganese oxide was used as an internal standard.. Scavenging activity was calculated as the ratio to blank sample that did not contain test compound. 3) Lipid peroxidation of rat brain homogenate The brain was isolated from euthanized Sprague Dawley rats at 4 weeks age and stripped of its meninges.. Five grams of brain was homogenized in 30ml phosphate 49.
(53) buffered saline (pH 7.4) and supernatant was obtained by centrifugation (750 X g, 10 min).. Supernatant was diluted to one-fifth with the same buffer.. Ten microliters of. each test substance such as carotenoid and a-tocopherol in DMSO were added to 990 JlI of diluted supernatant in container.. Containers were transferred to a 37°C water. bath and incubated continuously for 1 h after addition of 0.02 mM FeCl 2 (final concentration).. Reaction was stopped in ice-cold water.. Malonyldialdehyde (MDA). was quantified using the calibration curve from absorbance at 532 nm after thiobarbituric acid reaction (12). The torularhodin used in these studies was isolated and purified from R. glutinis no. 21.. d,l-a-Tocopherol and. ~-carotene. Industries as comparative controls.. were purchased from Wako Pure Chemical. Addition of DMSO without test substances was. performed as a negative control.. 50.
図


+7



Outline
関連したドキュメント
Keywords: continuous time random walk, Brownian motion, collision time, skew Young tableaux, tandem queue.. AMS 2000 Subject Classification: Primary:
This paper develops a recursion formula for the conditional moments of the area under the absolute value of Brownian bridge given the local time at 0.. The method of power series
[Tetali 1991], and characterises the weights (and therefore the Dirichlet form) uniquely [Kigami 1995]... RESISTANCE METRIC, e.g. equipped with conductances) graph with vertex set V
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
Applications of msets in Logic Programming languages is found to over- come “computational inefficiency” inherent in otherwise situation, especially in solving a sweep of
Shi, “The essential norm of a composition operator on the Bloch space in polydiscs,” Chinese Journal of Contemporary Mathematics, vol. Chen, “Weighted composition operators from Fp,
[2])) and will not be repeated here. As had been mentioned there, the only feasible way in which the problem of a system of charged particles and, in particular, of ionic solutions
This paper presents an investigation into the mechanics of this specific problem and develops an analytical approach that accounts for the effects of geometrical and material data on
