学位論文 博士(医科学)甲
Mechanism of Action Analysis of Allergy
Immunotherapy for Japanese Cedar Pollinosis
Patients: Evaluation of Blocking Activity for
the Allergen-IgE Binding using Cell Free
Biomarker Assay
スギ花粉症患者に対するアレルゲン免疫療法の作用機序解
析:バイオマーカーアッセイを用いたアレルゲン-IgE 結合
に対するブロッキング能の評価
深野 千陽
山梨大学
Abbreviations
AIT Allery specific immunotherapy
ELIFAB Enzyme-Linked Immunosorbent Facilitated Antigen Binding ELISA Enzyme-Linked Immunosorbent Assay
FAB Facilitated antigen binding FAP Facilitated antigen presenting JAU Japanese allergy unit
JC Japanese cedar
JSA Japanese Society of Allergology SCIT Subcutaneous immunotherapy SLIT Sublingual immunotherapy
Contents
1. Introduction ... 5
Japanese Cedar Pollinosis ... 5
Allergy Immunotherapy ... 5
Mechanism of Action of Allergy Immunotherapy ... 6
2. Materials and Methods ... 10
2.1 Study subjects and sera ... 10
2.2 Antibodies and reagents ... 10
2.3 Enzyme-linked immunosorbent-facilitated antigen binding (ELIFAB) assay for Japanese cedar pollen allergen ... 11
2.4 Assessment of CD23 and IgE dependency ... 11
2.5 Detection of treatment-induced IgE-blocking activity in sera from JC pollen SLIT/ SCIT patients ... 12
2.6 Measurement of allergen specific IgG4 ... 12
2.7 Data analysis and statistics ... 12
3. Results ... 14
3.1 Immobilized concentration of CD23 ... 14
3.2 Effect of allergen concentration ... 14
3.3 Dependency of CD23 and IgE ... 14
3.4 ELIFAB for SCIT patients ... 14
3.5 Correlation between ELIFAB and allergen specific IgG4 ... 15
3.6 ELIFAB for SLIT patients ... 15
5. Conclusion ... 27 ACKNOWLEDGMENTS ... 28 REFERENCES ... 29
5
1. Introduction
Japanese Cedar Pollinosis
Japanese Cedar (JC; Cryptomeria japonica or ‘sugi’) pollinosis is a serious type 1 allergic rhinitis caused by airway exposure of JC pollen. Sneezing, rhinorrhea, nasal blockage and itchy eyes are the main symptoms of JC pollinosis, which seasonally occurs from February to April [1, 2]. The annual numbers of JC pollinosis patients have been increasing since the disease was recognized as hay fever in 1964 [2]; the disease become a health problem in Japan. As treatment for JC pollinosis, pharmacotherapy has been the major choice of treatment, however the ratio of JC pollinosis patients who are satisfied with anti-histamine treatment was estimated as 35% [3].
Allergy Immunotherapy
Allergy immunotherapy (AIT) was developed in 1911 by Noon as safe and effective treatment for allergic diseases [4–8]. Repeated subcutaneous or sublingual administration of allergens to allergic patients gradually establishes immunological tolerance for sensitizing allergens. Unlike the pharmacotherapy, AIT has properties for disease-modification and confers long-term clinical benefit after cessation of treatment [9]. In Japan, subcutaneous immunotherapy (SCIT) was introduced in the 1960s to treat rhinitis and/or asthma caused by allergic sensitization. The use of non-standardized JC pollen extract for SCIT began in 1969 and continued until 1996. In 1996, standardized JC pollen allergen extract was developed by the Japanese Allergology Society (JSA), according to the concept published by the Center for Biologics Evaluation and Research (CBER) in the USA. For biological standardization, allergen extracts was maintained the quality of allergens by determining the overall IgE-binding potency.
6
alternative to SCIT [10, 11]. It is considered that Scadding first reported the efficacy of SLIT for house dust mite allergy in 1986 [12]. Regarding to JC pollen extract, a modified formulation of SCIT product of standardized JC pollen extract named CEDARTOLEN® was developed by Torii Pharmaceutical Co. Ltd., for application of SLIT and registered in 2014 with an induction for adults and adolescent (>12 years) patients. Some clinical trials revealed the efficacy of SLIT [10, 13]. In these trials, total nasal symptom and medication scores (TNSMS) were significantly reduced compared with the placebo control. AIT is a unique treatment that can change the natural course of allergic diseases. It is expected to improve the symptom of JC pollinosis patients, and to reduce the amount of drugs used for pharmacotherapy. In contrast, there are invalid subjects for AIT. The mechanism of AIT is not fully understood; many studies are being conducted to reveal the mechanism of AIT, and to find useful biomarkers as indicators of treatment completion or to predict treatment effects before AIT.
Mechanism of Action of Allergy Immunotherapy
The immunologic basis of allergic diseases is observed in two phases; the early phase and late phases. The events in the early phase are sensitization and development of memory T cell and B cell responses and IgE. CD4+ Th2 cells producing IL-4 and IL-13 induce class switching in B cells, which produce allergen specific IgE (s-IgE). Allergen-specific IgE leads to the sensitization by binding to the high-affinity transmembrane IgE receptor FcRI on the surface of mast cells and basophils [14]. The events in the late phase are effector functions related to tissue inflammation and injury. Allergen mediated cross-linking of the IgE FcRI complexes on sensitized mast cells and basophils will lead to cellular degranulation and immediate release of histamine, leukotorienes and other chemical mediators (type 1 hypersensitivity). CD4+ naïve T
7
cells can differentiate into type memory and effector cells like Th1 and Th2, depending on the innate immune response activating capacity with allergens and cytokines. Th2 cells produce IL-4, IL-5, IL-9 and IL-13 during the development of allergic diseases [15-17]. These cytokines play a role in the production of allergen s-IgE, eosinophil and the other inflammations.
The mechanism of successful AIT still remains to be established in full, but it is considered that it involves cellular reactions and antibody response. For cellular response, a recurring observation in AIT is the immune-deviation induced by treatment where the ratio of allergen-specific Th1: Th2 cells increases, driven primarily by a reduction in Th2 cells rather than an increase in Th1 response [18]. Cellular changes include the generation of allergen-specific T regulatory cells (Tregs) which produce IL-10 or TGF- and B regulatory cells (Bregs) which produce IL-10 and have inhibition capacity for inflammatory response, and suppression of allergen s-Th2 cells [19-21].
For antibody response, several pollen AIT studies reported that both SCIT and SLIT have been associated with transient early increases in serum allergen s-IgE levels that are followed by blunting of the usual seasonal increases in IgE levels during natural allergen exposure [22]. In addition, serum level of allergen s-IgG, in particular s-IgG4 is currently considered a candidate for biomarkers not being associated with clinical efficacy [23, 24]. Several studies on AIT have reported 10-100 fold increases in IgG (s-IgG4) concentration [25-28]. It has also been revealed that SLIT induces allergen s-IgG1, s-IgG4 and IgA [4, 29, 30]. These non-IgE antibodies induced by AIT are thought to compete with s-IgE for binding to allergens, inhibiting allergen s-IgE complex formation on mast cells, basophils and the other s-IgE receptor-expressing cells, and thereby reducing the activation of effector cells. To investigate the concept of
8
“blocking” antibodies, several assays, such as IgE-facilitated allergen presentation or binding assay and IgE-blocking factors, have been developed to measure serum inhibitory activity [4, 25, 31, 32].
In facilitated antigen presentation (FAP), allergen molecules bound to two or more allergen-specific IgEs are captured by the low-affinity IgE receptor CD23 (FcRII) expressed on the surface of antigen presenting cells. After internalization,
allergen-derived peptides are presented to allergen-specific T-cells, and the level of T-cell activation correlates with the number of IgE-allergen-CD23 complexes formed. In the absence of allergen, binding of monomeric IgE and CD23 is weak and transient due to low affinity, but when two or more allergen-specific IgE antibodies combine by binding to the same allergen molecule, the avidity of the IgE-CD23 association
increases and stable IgE-allergen-CD23 complexes can form [25].
A variant of the FAP assay is the facilitated allergen binding (FAB) assay. Both FAP and FAB measure the formation of IgE-allergen-CD23 complexes but in FAB, the relative amounts of cell surface-bound allergen are determined by
fluorescence-activated cell sorting (FACS). Inhibition of IgE-allergen-CD23 complexes by AIT-induced IgE-blocking antibodies can be measured in vitro by both assays.
A major obstacle for routine use and adaptation for high-throughput protocols is that these assays require whole blood from allergic patients or freshly cultured Epstein-Barr virus (EBV)-transformed B-cells that maintain a high level of CD23 expression. As an alternative to the cell-based assays, the aim of this study was to establish a cell-free, Enzyme-linked Immunosorbent assay (ELISA)-based FAB (ELIFAB) assay with JC pollen allergens with the purpose to examine the effects of JC AIT-induced IgE-blocking antibodies on FAB and provide further insight into the immunological mechanism of action of successful AIT with JC pollen extract.
9
To find the surrogate biomarkers predictive or indicative of the effects of AIT, treatment could be more patient-friendly to maintain patient’s motivation, because AIT requires a long time more than 3 years.
10
2. Materials and Methods
2.1 Study subjects and seraThe JC pollen-specific IgE titer of serum samples was measured using the ImmnoCAP assay. From in-house volunteers, patients sensitized to JC pollen were selected and comprised the untreated “allergic group”. Patients allergic to JC pollen who have been treated with sublingual or subcutaneous immunotherapy with standardized JC pollen extract comprised the “SLIT group” or “SCIT group”. In the SLIT group, patients treated with JC pollen extract droplet of 2,000 JAU or placebo for 1 year (visit 12), 2 year (visit 24) and pretreatment (visit 1) in clinical trial were applied (n=40, male or female, 12-64 years old, and 2 years history of JC pollinosis). Patients received SCIT with JC pollen extract (Torii) at University of Yamanashi as a maintenance dose of 2,000 JAU for at least 48 months (n=11, male and female, 22–65 years old). Patients which symptom was improved by AIT were applied in this study. All procedures were approved by the Ethics Committees of University of Yamanashi and Torii Pharmaceutical Co. Ltd. according to the Clinical Research Ethical Guidelines. Written informed consent was obtained from all subjects before the study.
2.2 Antibodies and reagents
The following antibodies and reagents were used: monomeric human recombinant soluble CD23 (sCD23, 123-FE-050; R&D Systems, MN); biotin-conjugated anti-human IgE antibody (555858, BD Biosciences, NJ); ExtrAvidin-Peroxidase (E2886-1ML, Sigma, MO); tetramethyl benzidine (T2885, Sigma, MO); Anti-CD23 monoclonal antibody (Ab16702, Abcam plc, Cambridge, UK); JC pollen allergen extract (200 µg of protein/mg of dry extract, Torii, Japan). Microtiter plates (MediSorp 96-well plate, Thermo, MA) and round-bottom plates (3799, Corning, NY) were also used. A
11
high-titered serum (JC pollen-specific IgE; 400 kU/L) from an in-house volunteer was used as ‘indicator serum’ [4, 22].
2.3 Enzyme-linked immunosorbent-facilitated antigen binding (ELIFAB) assay for Japanese cedar pollen allergen
The procedure for ELIFAB used for Phleum pretense [22] was adapted and optimized for JC pollen allergen [33]. Briefly, microtiter plates were coated with CD23 overnight at 25°C. The plates were then washed and blocked with 1% bovine serum albumin/phosphate-buffered saline for 1 h at 37°C. The indicator serum was mixed with an equal volume of medium (RPMI1640, 11875093, Thermo, MA) or serum from untreated or JC pollen SLIT/SCIT treated patients, and incubated with JC pollen allergen extract for 1 h at 37°C with in round-bottom plates before transfer to a CD23-coated microtiter plate for IgE-allergen CD23 complex formation. Allergen-IgE CD23 complexes were detected using a biotin-conjugated anti-human IgE antibody with a standard streptavidin peroxidase/tetramethyl benzidine detection method. The level of allergen-IgE complexes binding to CD23 was expressed as absorbance at 450 nm (SectraMax Plus 384, Molecular Devices). To investigate the assay dependency, anti-CD23 antibody and the denatured indicator serum were used.
2.4 Assessment of CD23 and IgE dependency
The ELIFAB assay procedure described above was used with modification [33]. Briefly, an anti-human CD23 monoclonal antibody was added to the CD23-coated plates and incubated for 30 min followed by washing before the transfer of the serum-allergen mixture. The indicator serum was mixed with an equal volume of the medium before incubation with the allergen extract.
12
Structurally intact IgE was selectively eliminated from the indicator serum by heating for 30min at 56°C [34].
2.5 Detection of treatment-induced IgE-blocking activity in sera from JC pollen SLIT/ SCIT patients
The effect of JC pollen SLIT/SCIT induced IgE-blocking antibodies on IgE-allergen-CD23 complex formation was measured by mixing the indicator serum in 1:1 ratio with serum samples from JC pollen sensitized patients who were either untreated (the “allergic group” or “placebo group”) or treated (the “SLIT group” and “SCIT group”) before incubation of the sera with JC pollen extract.
2.6 Measurement of allergen specific IgG4
Allergen-specific IgG4 titers were determined using Phadia ImmunoCAP system [35]. Immobilized allergen CAP of JC pollen (Uppsala, Thermo Fisher Scientific) was used. All reagents (wash buffer, dilution buffer, enzyme-labeled antibody, development solution, and stop solution) were obtained from Thermo Fisher Scientific, Sweden. This test method was validated at Torii pharmaceuticals. Data were expressed as mgA/L, with a cut off value of 0.07 mgA/L (lower) and 30 mgA/L (upper).
2.7 Data analysis and statistics
For the ELLIFAB assay, data are expressed as the relative binding to allergen-IgE complexes where binding with indicator serum alone was normalized to 100%. The following formula was used:
% relative allergen-IgE complex binding to CD23 = (Abs of indicator serum and test samples / Abs of indicator serum only) x 100
13
Statistical analysis was performed using GraphPad Prism Software (version 6.07, GraphPad Inc. CA). Data for the binding ability to CD23 in SCIT patients are presented as the median values. The statistical significance of differences between the two groups was evaluated using the Kruskal-Wallis test followed by the Dunn’s post hoc test. Linear regression was employed to analyze between the binding ability to CD23 and JC pollen specific IgG4.
Data in the SLIT patients, paired t test was used to analyze between differences in pretreatment (visit 1) and posttreatment (visit 12). The Mann-Whitney U test was used to analyze between group differences in the binding ability to CD23. When p value was <0.05, the difference was considered significant.
14
3. Results
3.1 Immobilized concentration of CD23
The optimal concentration of CD23 for ELISA plate coating was determined by a wide range of CD23 concentrations (5–50 g/mL) with the indicator serum mixed with JC pollen extract (1 g/mL). The complex formation of allergen-IgE-CD23 reached a plateau at 20 g/mL of CD23 (Figure 1).
3.2 Effect of allergen concentration
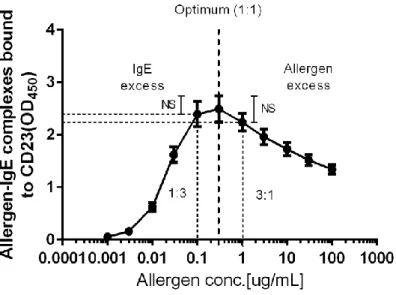
The formation of allergen-IgE-CD23 complexes at various concentrations was examined following serial dilutions of JC pollen extract (0.001–100 g/mL) with the indicator serum and ELISA plates coated with 20 g/mL of CD23. The peak response was observed at 0.3 g/mL of JC pollen allergen (Figure 2), which was set as the standard condition for the assay.
3.3 Dependency of CD23 and IgE
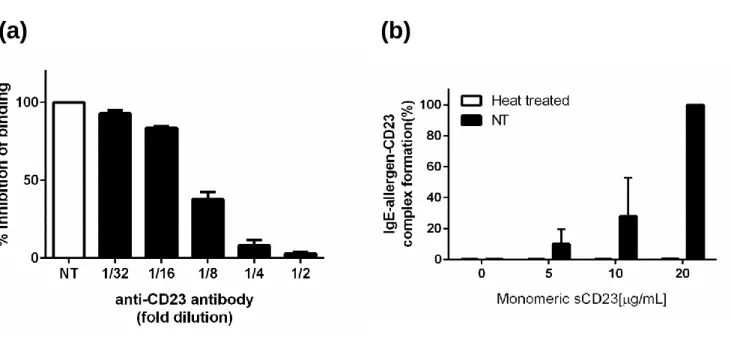
The dependency of the assay on CD23 was investigated using an anti-CD23 antibody. With the anti-CD23 antibody on the CD23 coated plate, the formation of allergen-IgE-CD23 complexes was abolished in a dose-dependent manner (Figure 3(a)). The dependency of the assay on IgE was assessed with the indicator serum, which was heat-treated prior to mixing with the allergen for removing structurally intact IgE (Figure 3(b)). These data indicate that both CD23 and IgE are essential components of the complex formation detected in this assay.
3.4 ELIFAB for SCIT patients
15
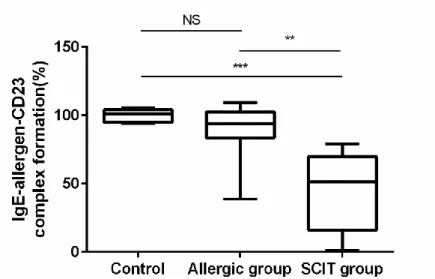
allergic group and SCIT group, n = 11) was used. When sera from the SCIT group were combined with the indicator serum, the magnitude of the allergen-IgE complex binding to CD23 reduced in a statistically (p < 0.01) to nearly half (51.5%) compared with the level of complex formation obtained with the indicator serum diluted with medium. A non-statistically significant difference (93.9%) was observed when the indicator serum was mixed with sera from the allergic group (Figure 4).
3.5 Correlation between ELIFAB and allergen specific IgG4
To understand the contribution of the levels of IgG4 to the ELIFAB assay, the
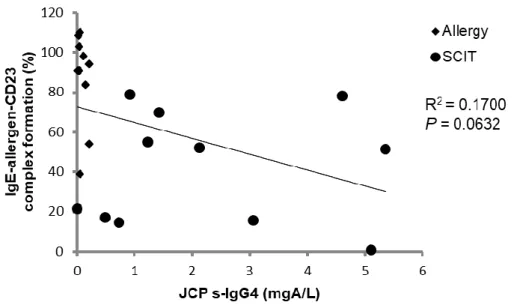
correlation between the levels of JC pollen-specific IgG4 and the IgE-allergen-CD23 complex formation (%) was investigated (Figure 5). In the SCIT group, IgE-CD23 complex formation was deemed to decrease depend on the level of JC pollen-specific IgG4; however, no correlation was observed between these two factors, while low JC pollen-specific IgG4 level was detected in the non-treated allergic group. This data consists of 11 subjects in each group. To investigate the correlation between the IgG4 levels in sera and the IgE-allergen complex formation (%), it was considered that a larger number of subjects were required. Additionally, patients have some extent of s-IgG4 originally. Therefore, it was considered that the change ratio of s-IgG4 from the baseline might affect the results of this assay.
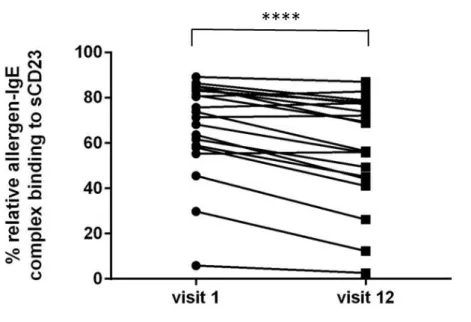
3.6 ELIFAB for SLIT patients
The ability of inhibition on allergen-IgE-CD23 complex formation was measured using the indicator serum and the established standard assay conditions. Serum samples treated with sublingual immunotherapy (pretreatment (visit 1) and posttreatment (visit 12)) were applied. Comparing the allergen-IgE complex binding to CD23 between pre-
16
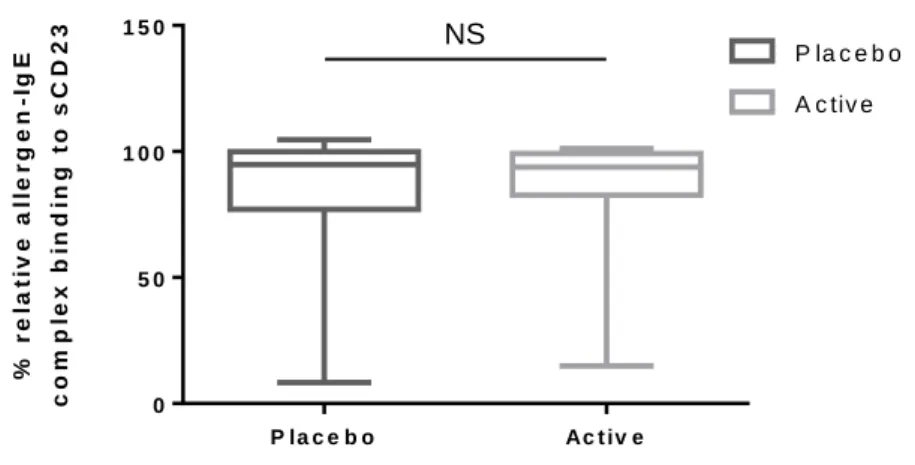
and post-treatment, the binding decreased significantly after SLIT treatment (Figure 6). However, no difference of the allergen-IgE complex binding to CD23 was observed between the placebo and the active-treated group (Figure 7). Recently, many clinical data have indicated that SLIT needs treatment periods that are more than 3 years and without relapse of allergic symptoms. The sublingual treatment with JC pollen for 2 years is not enough duration to change immunological response, which is detectable by functional assays completely.
17
Figure 1. Optimization of immobilized CD23 concentration
Binding ability of IgE-allergen complexes depending on concentration of immobilized CD23. Indicator serum
from a subject with JC pollen allergy was incubated with JC pollen (1 μg/mL) before immune complex capture.
18
Figure 2. Optimization of allergen concentration
Effect of JC pollen allergen concentration on allergen–IgE binding to immobilized CD23. The indicator serum was
applied to this experiment. Three independent experiments were performed. Data are expressed as mean ± SE
(absorbance at 450 nm). Heavy dashed line: Optimal allergen: IgE ratio. Vertical dotted lines: IgE excess (1:3) or
19
Figure 3. Specificity(CD23 and IgE dependency)
Assessment of assay dependency. Three experiments were performed for each test. Values represent mean ± SE.
(a) CD23 dependency was investigated using the anti-CD23 antibody. Data are shown as percentage of inhibition
binding which indicator serum without adding of anti-CD23 was normalized 100%. (b) IgE dependency was assessed
using denatured serum. Data was expressed as % relative allergen-IgE complex binding to CD23. NT; Non treated.
(b)
20
Figure 4. Inhibitory activity of allergen-IgE binding to CD23 in patients with JC pollen SCIT
Data are expressed as median values with 1st and 3rd quartiles (n=11). Statistical significance was detected by using
the Kruskal-Wallis test followed by the Dunn post hoc test. ** p <0.01 and ***p<0.001, NS: Non-significant.
Control: indicator serum diluted with medium. Allergic group: Sera from JC pollen-sensitized subjects diluted with
21
Figure 5. Correlation between JC pollen specific IgG4 and IgE-allergen-CD23 complex formation in SCIT group
The relationship between IgE-allergen-CD23 complex formation by ELIFAB assay and JC pollen specific IgG4,
22
Figure 6. Change of allergen-IgE-CD23 complex formation (%) with pre and post SLIT in same patient
Serum samples of patients (n=20) were used. Statistical significance was detected by using the paired t test between
23 P la c e b o Ac t iv e 0 5 0 1 0 0 1 5 0 % r e la ti v e a ll e r g e n -I g E c o m p le x b in d in g t o s C D 2 3 P la c e b o A c tiv e
Figure 7. Allergen-IgE-CD23 complex formation (%) with placebo and SLIT group at visit24
Data are expressed as median values with 1st and 3rd quartiles (n=20 in each group). No statistical significance was
detected by using the Mann-Whitney U test. NS: Non-significant.
24
4. Discussion
AIT-treatment-induced increases in allergen-specific antibodies block the binding of the allergen to IgE. This mechanism is a consistent biomarker of AIT (e.g., tree- and grass-pollens and JC pollen [4, 36-40].
FAP occurs as complexes between allergen, IgE and CD23 expressed on the surface of B cells, which are formed and subsequently internalized and processed for MHC II presentation; this may occur at extremely low allergen concentrations because of the selective capture of allergen by IgE. FAP strongly enhances the activation of allergen-specific T cells and the subsequent secretion of pro-inflammatory Th2 cytokines. Inhibition of this process by IgE-blocking antibodies may result in the down regulation of Th2 T cell responses and result in a reduction in allergic responses in allergic patients.
To measure the formation of allergen-IgE-CD23 complexes in vitro, the cell-based IgE-facilitated allergen binding (IgE-FAB) assay was developed [25, 41]. Previous studies using IgE-FAB assay have shown that AIT with birch pollen extract inhibits the formation of allergen-IgE-CD23 complexes compared to the baseline and placebo and that the observed reduction is associated with the development of clinical tolerance [31]. Although IgE-FAB is well validated, the disadvantage of the assay are prolonged culturing and the difficulty in maintaining viable EBV-transformed B cells expressing FcRII/CD23; these limitations are a barrier to the routine use of the assay. As an alternative, the more simple method ELIFAB which needs no cell lines was developed using Phleum pratense allergen [22]. There is no difference between IgE-FAB and ELIFAB assays with regards to specificity and sensitivity [22], but the more simple method was required for routine use. Both IgE-FAB and ELIFAB are allergen-specific assays. The basic protocol of ELIFAB assay was developed by Shamji and colleagues
25
[22], however, careful selection of reagents and optimization of the experimental conditions are required for each individual allergen. The ELIFAB assay depends on multiple factors (s-IgE concentration, composition and affinity of patient sera or allergen species) that determine the effectiveness of complex formation [42-44].
Having established the optimal assay conditions [33], the effect of JC pollen AIT on the formation of allergen-IgE-CD23 complexes could be investigated. When sera from SCIT patients and allergic patients were applied in the ELIFAB assay, the formation of allergen-IgE-CD23 complexes was significantly different between these two groups [33]. When sera from the SLIT patients was applied in the ELIFAB assay, the allergen-IgE-CD23 complex formation reduced after one-year SLIT treatment compared with the baseline in individual SLIT samples (visit 1 vs. visit 12). To obtain more information about the blocking activity associated with clinical tolerance, allergen-IgE-CD23 complex formation was compared between the placebo and SLIT groups. However no significant difference was observed between the two groups. As Durham mentioned in his report, there was no difference between SLIT and SCIT in rhinitis symptoms or medication scores. Additionally, a large DBPC clinical trial (TO-194SL-3-1, CEDARTOLEN®, 2,000 JAU) demonstrated clinical efficacy following a significant increase in serum allergen-specific IgG4 in the SLIT group. However, different results of ELIFAB assay were obtained in SCIT and SLIT in this study. Sera from 2 years SLIT and SCIT, which continued at least 4 years, were used as serum samples, and this difference in the treatment period induced different magnitudes in the ELIFAB assay.
Before AIT, patients had detectable allergen-specific IgG4 (data not shown), probably originating from natural exposure; however, there was no measurable blocking activity. The lack of IgE-blocking activity despite the presence of IgG4 could be due to low
26
IgG4 concentrations or low IgG4 affinity compared to allergen-specific IgE. The difference between the degree of increase in IgG4 and IgE blocking factors during the AIT up-dosing and maintenance phases indicates that, in addition to IgG4 concentrations, other factors such as clonality and affinity maturation also influenced blocking activity [45].
It was reported that the change in immunoreactive IgG4 levels showed a poor correlation with the clinical response as measured by symptom and medication scores [23, 24]. One possible explanation for this discrepancy is that AIT may have induced the production of new s-IgG4 with augmented binding specificity and/or affinity. To investigate the function of allergen s-IgG4 induced by AIT, not only the blocking activity but also the structure or the character of the antibody itself should be evaluated. Regarding the establishment of ELIFAB in this study, some data is required to increase reliability. To investigate the contribution of allergen s-IgG4 as blocking factor, purified IgG4 antibody should be applied in the ELIFAB assay. Additionally, the expression rate of CD23 on B cells should be confirmed to understand how ELIFAB assay could reflect the in vivo environment. Furthermore, to confirm the concept of the mechanism of action of AIT, further studies with a large number of samples from adequate JC pollen AIT DBPC clinical trials are required to establish the correlation between clinical improvement and IgE-inhibitory activity detected by ELIFAB assay, and some other factors such as cytokines, chemokines and cellular biomarkers like Treg cells need to be simultaneously investigated with JC pollen.
27
5. Conclusion
ELIFAB for JC pollen was newly established for confirming the concept of blocking factor. By using the ELIFAB assay, it was revealed that the formation of the
allergen-IgE CD23 complex was significantly reduced by one year SLIT in identical subjects and by SCIT compared with the allergic patients group. This study provides proof of principle for the JC pollen ELIFAB assay as a suitable biomarker assay for monitoring the immunological response of groups and individual patients to JC pollen AIT.
28
ACKNOWLEDGMENTS
I would like to express my deep gratitude to all those who provided guidance, support and encouragement during the preparation of this dissertation.
Most of all, I would like to express my sincere appreciation to Professor Atsuhito Nakao (Department of Immunology, Faculty of Medicine, University of Yamanashi) for valuable guidance and critical reading of this manuscript.
I would like to express my deep gratitude to Dr. Atsuyuki Kakee and Mr. Hideki Ozaki (Torii Pharmaceutical Co. Ltd.) for providing the opportunity of the research and encouragement in preparation of manuscript.
I would like express my special gratitude to Dr. Katsuyo Ohashi-Doi and Dr. Yuko Mitobe (Torii Pharmaceutical Co. Ltd.) for invaluable guidance and constructive discussions.
I am deeply indebted to Dr. Tomokazu Matsuoka (University of Yamanashi) Mr. Keisuke Suzuki and Ms. Satoko Kobayashi (Torii Pharmaceutical Co. Ltd.) for their excellent technical supports.
29
REFERENCES
1) Isizaki T, Koizumi K, Ikemori R, Ishiyama Y, and Kushibiki E., Studies of
prevalence of Japanese cedar pollinosis among the residents in a densely cultivated area. Ann Allergy, 1987. 58: 265-270.
2) Horiguchi S and Saito Y., Japanese Cedar Pollinosis in Nikko, Japan. Japanese Journal of Allergology, 1964. 13(1-2): 16-18, 74-75.
3) Konno A, Kubo N., Patient satisfaction with the second generation antihistamines
in the treatment of pollinosis and improved awareness on medical consultation – choice of drugs based on a large-scale questionnaire survey in patients with pollinosis. Prog Med, 2008. 28: 2285–96.
4) Shamji MH, Ljorring C, Francis JN, Calderon MA, Larche M, Kimber I, et al.,
Functional rather than immunoreactive levels of IgG4 correlate closely with clinical response to grass pollen immunotherapy. Allergy, 2012. 67: 217-226.
5) Durham SR, Walker SM, Varga EM, Jacobson MR, O’Brien F, Noble W, et al.,
Long-term clinical efficacy of grass-pollen immunotherapy. N Engl J Med, 1999.
341: 468-475.
6) Matsuoka T, Shamji MH, Durham SR., Allergen immunotherapy and tolerance. Allergol Int, 2013. 62: 403-413.
7) Akdis M, Akdis CA., Mechanisms of allergen- specific immunotherapy: multiple
suppressor factors at work in immune tolerance to allergens. J Allergy Clin
Immunol, 2014. 133: 621-631.
8) Calderon MA, Penagos M, Sheikh A, Canonica GW, Durham SR., Sublingual
immunotherapy for allergic conjunctivitis: Cochrane systematic review and meta-analysis. Clin Exp Allergy, 2011. 41: 1263-1272.
30
SQ-standardized sublingual grass immunotherapy: confirmation of disease modification 2 years after 3 years of treatment in a randomized trial. J Allergy Clin
Immunol, 2012. 129: 717-725 e715.
10) Gotoh M, Yonekura S, Imai T, Kaneko S, Horikawa E, Konno A, et al., Long-Term
Efficacy and Dose-Finding Trial of Japanese Cedar Pollen Sublingual Immunotherapy Tablet. J Allergy Clin Immunol Pract, 2019. 7(4): 1287-1297. e8
11) Durham SR, Creticos PS, Nelson HS, Li Z, Kaur A, Meltzer EO, et al., Treatment
effect of sublingual immunotherapy tablets and pharmacotherapies for seasonal and perennial allergic rhinitis: Pooled analyses. J Allergy Clin Immunol, 2016.
138(4): 1081-1088 e4.
12) Scadding GK, Brostoff J., Low dose sublingual therapy in patients with allergic
rhinitis due to house dust mite. Clin Allergy, 1986. 16(5): 483‒491.
13) Okubo K, Gotoh M, Fujieda S, Okano M, Yoshida H, Morikawa H, et al., A
randomised double-blind comparative study of sublingual immunotherapy for cedar pollinosis. Allergol. Int, 2008. 57: 265–275.
14) Wang M, Takeda K, Shiraishi Y, Okamoto M, Dakhama A, Joetham A, et al.,
Peanut-induced intestinal allergy mediated through a mast cell-IgE FcRI IL-13
pathway. J Allergy Clin Immunol, 2010. 126: 306-16.
15) Akdis M., Healthy immune response to allergens: T regulatory cells and more. Curr Opin Immunol, 2006. 18: 738–44.
16) Akdis M, Akdis CA., Mechanisms of allergen-specific immunotherapy. J Allergy Clin Immunol, 2007. 119: 780–91.
17) Larche M, Akdis CA, Valenta R., Immunological mechanisms of allergen-specific
immunotherapy. Nat Rev Immunol, 2006. 6: 761–71.
31
allergens and allergen immunotherapy. J Allergy Clin Immunol, 2018. 142:
1697-1709.
19) Wachholz PA, Nouri-Aria KT, Wilson DR, Walker SM, Verhoef A, Till SJ, et al.,
Grass pollen immunotherapy for hayfever is associated with increases in local nasal but not peripheral Th1:Th2 cytokineratios. Immunology, 2002. 105: 56–62.
20) Bohle B, Kinaciyan T, Gerstmayr M, Radakovics A, Jahn-Schmid B, Ebner C.,
Sublingual immunotherapy induces IL-10-producing T regulatory cells, allergen-specific T-cell tolerance, and immune deviation. J Allergy Clin Immunol,
2007. 120: 707-13.
21) Rosser EC, Mauri C., Regulatory B cells: origin, phenotype, and function. Immunity, 2015. 42: 607-12.
22) Shamji MH, Francis JN, Würtzen PA, Lund K, Durham SR, Till SJ., Cell-free
detection of allergen-IgE cross-linking with immobilized phase CD23: inhibition by blocking antibody responses after immunotherapy. J Allergy Clin Immunol, 2013.
132: 1003-5, e1-4.
23) Flicker S, and R Valenta., Renaissance of the blocking antibody concept in type I
allergy. Int. Arch. Allergy Immunol, 2003. 132: 13–24.
24) van Neerven, RJ, Knol EF, Ejrnaes A, and Würtzen PA., IgE mediated allergen
presentation and blocking antibodies: regulation of T-cell activation in allergy. Int
Arch Allergy, 2006. 141(2): 119-29.
25) Wachholz PA, Soni NK, Till SJ, Durham SR., Inhibition of allergen-IgE binding to
B cells by IgG antibodies after grass pollen immunotherapy. J Allergy Clin
Immunol, 2003. 112: 915-22.
26) Reisinger J, Horak F, Pauli G, van Hage M, Cromwell O, Konig F, et al.,
32
modified allergens are associated with reduced nasal allergen sensitivity. J Allergy
Clin Immunol, 2005. 116: 347-54.
27) Oefner CM, Winkler A, Hess C, Lorenz AK, Holecska V, Huxdorf M, et al.,
Tolerance induction with T cell-dependent protein antigens induces regulatory sialylated IgGs. J Allergy Clin Immunol, 2012. 129: 1647-55.e13.
28) Gleich GJ., The late phase of the immunoglobulin E-mediated reaction: a link
between anaphylaxis and common allergic disease? J Allergy Clin Immunol, 1982.
70: 160-9.
29) Scadding GW, Calderon MA, Bellido V, Koed GK, Nielsen NC, Lund K, et al.,
Optimization of grass pollen nasal allergen challenge for assessment of clinical and immunological outcomes. J Immunol Methods, 2012. 384: 25-32.
30) Francis JN, James LK, Paraskevopoulos G, Wong C, Calderon MA, Durham SR, et al., Grass pollen immunotherapy: IL-10 induction and suppression of late responses
precedes IgG4 inhibitory antibody activity. J Allergy Clin Immunol, 2008. 121:
1120-5.e2.
31) Würtzen PA, Lund G, Lund K, Arvidsson M, Rak S, and Ipsen H., A double-blind
placebo-controlled birch allergy vaccination study II: correlation between inhibition of IgE binding, histamine release and facilitated allergen presentation.
Clin. Exp. Allergy, 2008. 38: 1290–1301.
32) Johansen N, Gronager PM, Ipsen H, Stranzl T, Lund K., Characterization of a new
method for quantitative determination of house dust mite allergen specific IgEblocking factor for monitoring allergy immunotherapy with SQ HDM SLIT-tablet. Allergy, 2016. 71: 118-272, ab321.
33) Fukano C, Ohashi-D K, Lund K, Nakao A, Masuyama K, Matsuoka T.,
33
biomarker assay for Japanese cedar pollen allergy immunotherapy. J Pharmacol
Sci, 2019. 140(3): 223-227.
34) Demeulemester C, Weyer A, Peltre G, Laurent M, Marchand F and David B.,
Thermoinactivation of human IgE: antigenic and functional modifications. Immnol,
1986. 57: 617-620.
35) Jarvis D, Zock JP, Heinrich J, Svanes C, Verlato G, Olivieri M, et al., Cat and dust
mite allergen levels, specific IgG and IgG4, and respiratory symptoms in adults. J
Allergy Clin Immunol, 2007. 119(3): 697-704.
36) Shamji MH, J.L.a.D.S., Serum Immunologic Markers for Monitoring
Allergen-Specific Immunotherapy. Immunol Allergy Clin North Am, 2011. 31:
311-323.
37) Scadding GW, Shamji MH, Jacobson MR, Lee DI, Wilson D, Lima MT, et al.,
Sublingual grass pollen immunotherapy is associated with increases in sublingual Foxp3-expressing cells and elevated allergen-specific immunoglobulin G4, immunoglobulin A and serum inhibitory activity for immunoglobulin E-facilitated allergen binding to B cells. Clin Exp Allergy, 2010. 40(4): 598-606.
38) Mothes N, Heinzkill M, Drachenberg KJ, Sperr WR, Krauth MT, Majlesi Y, et al.,
Allergen-specific immunotherapy with a monophospphoryl lipid A-adjuvanted vaccine reduced seasonally boosted IgE production and inhibition of basophil histamine release by therapy-induced blocking antibodies. Clin Exp Allergy, 2003.
33: 1198-1208.
39) Ejrnaes AM, Svenson M, Lund G, Larsen JN and Jacobi H., Inhibition of rBet v
a-induced basophil histamine release with specific immunotherapy-induced serum
IgG no evidence that FcRIIB signaling is important. Clin Exp Allergy, 2006. 36:
34
40) Lambin P, Bouzoumou A, Murrieta M, Debbia M, Rouger P, Leynadier F, et al.,
Purification of human IgG4 subclass with allergen-specific blocking activity. J
Immunol Methods, 1993. 165: 99-111.
41) Nouri-Aria KT, Wachholz PA, Francis JN, Jacobson MR, Walker AM, Wilcock LK, et al., Grass pollen immunotherapy induces mucosal and peripheral IL-10
responses and blocking IgG activity. J. Immunol, 2004. 172: 3252–3259.
42) Willumsen N, Holm J, Christensen LH, Würtzen PA, and Lund K., The complexity
of allergic patients' IgE repertoire correlates with serum concentration of allergen-specific IgE. Clin Exp Allergy, 2012. 42(8): 1227-36.
43) Holm J, Willumsen N, Würtzen PA, Christensen LH, and Lund K., Facilitated
antigen presentation and its inhibition by blocking IgG antibodies depends on IgE repertoire complexity. J Allergy Clin Immunol, 2011. 127(4): 1029-37.
44) Christensen LH, Holm J, Lund G, Riise E, and Lund K., Several distinct properties
of the IgE repertoire determine effector cell degranulation in response to allergen challenge. J Allergy Clin Immunol, 2008. 122(2): 298-304.
45) Zhao D, Lai X, Tian M, Jiang Y, Zheng Y, Gjesing B, et al., The Functional
IgE-Blocking Factor Induced by Allergen-Specific Immunotherapy Correlates with IgG4 Antibodies and a Decrease of Symptoms in House Dust Mite-Allergic Children. Int Arch Allergy Immunol, 2016. 169(2): 113-20.