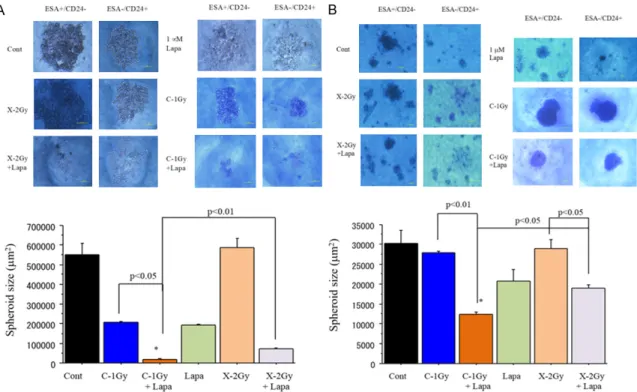
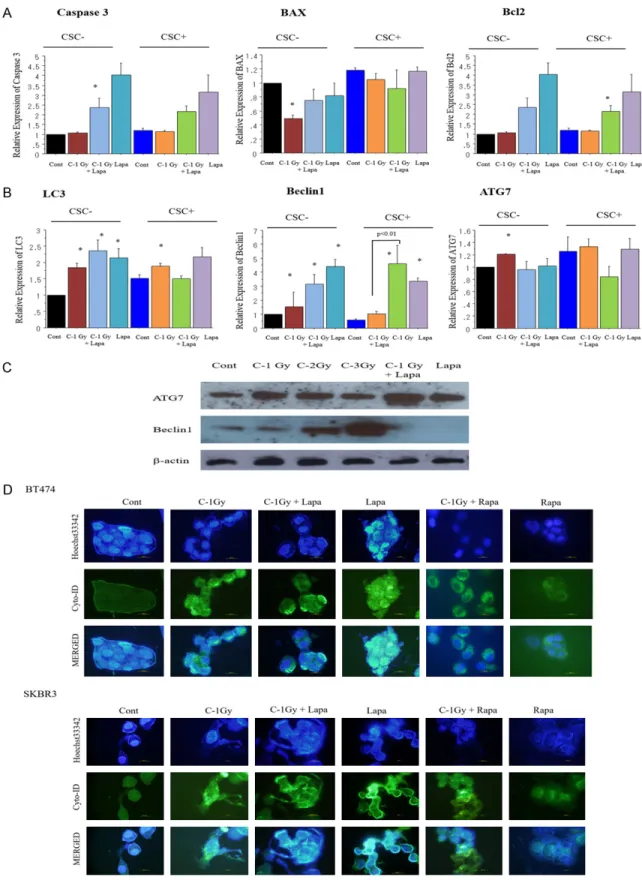
tyrosine kinase inhibitor, lapatinib,
effectively destroys HER2 positive breast cancer stem‑like cells
Author Sai Sai, Eun Ho Kim, Guillaume Vares, Masao Suzuki, Dong Yu, Yoshiya Horimoto, Mitsuhiro Hayashi
journal or
publication title
American Journal of Cancer Research
volume 10
number 8
page range 2371‑2386
year 2020‑08‑01
Publisher e‑Century Publishing
Rights (C) 2020 e‑Century Publishing Author's flag publisher
URL http://id.nii.ac.jp/1394/00001673/
Creative Commons Attribution‑NonCommercial 4.0 International (CC BY‑NC 4.0)
Original Article
Combination of carbon-ion beam and dual tyrosine kinase inhibitor, lapatinib, effectively
destroys HER2 positive breast cancer stem-like cells
Sei Sai
1, Eun Ho Kim
2, Guillaume Vares
3, Masao Suzuki
1, Dong Yu
4, Yoshiya Horimoto
5, Mitsuhiro Hayashi
61