ロジスティック回帰モデルを用いた環境指標によるツキノワグマ(
Ursus thibetanus japonicus
)の
生息確率予測モデル
Predictive Model for the Probability of Occurrences of Asiatic black bear (
Ursus thibetanus
japonicus
) using a Logistic Regression Model with Environmental Indices
土光 智子 *・福井 弘道**・大澤 啓志***・一ノ瀬友博****
Tomoko DOKO, Hiromichi FUKUI, Satoshi OSAWA, Tomohiro ICHINOSE
要旨:害獣でありアンブレラ種でもあるツキノワグマは,有効な生息地管理が期待されており,本種の生息地 推定には意義があると思われる。本研究では,環境指標によるツキノワグマ生息確率予測モデルを開発した。 ロジスティック回帰モデルは,log(p/(1-p))=(-1.486e+01)+(7.335e-04)*
x
1+(9.470e-03)*x
2であった。ここで
x
1 は徒歩道・庭園路等への距離(m),x
2 は標高(m),p は本種の生息確率を示している。本モデルは,生息地管理に応用が可能であり,今後は他の地域個体群生息確率予測への適用可能性の検証が望まれる。
キーワード:生息確率予測モデル,ツキノワグマ,Ursus thibetanus japonicus,ロジスティック回帰モデル,
環境指標
Abstract:Habitat estimation of Asiatic black bear is considered highly significant from the perspective of effective habitat management, because this species is known both as a destructive animal as well as an umbrella species. In this study, we developed a predictive model for the probability of occurrences of Asiatic black bear using environmental indices. Based on this research, the optimal logistic regression model was log(p/(1-p))=(-1.486e+01)+(7.335e-04)*
x
1+(9.470e-03)*x
2. wherex
1is the distance to small paths (m),x
2 is the altitude (m), and p is a probability of occurrence of Asiatic black bear in this study area. Thismodel is applicable for habitat management. Additional research will examine the application of this model to different local Asiatic black bear populations in other regions.
Keywords: predictive model for the probability of occurrences, Asiatic black bear, Ursus thibetanus japonicus, logistic regression model, environmental indices
はじめに
2010 年に,生物多様性条約第 10 回目締約国会議 (COP10)が名古屋で開催されるにあたり,生物多様性保 全への関心が高まっている。我が国でも,2007 年,第 3 次生物多様性国家戦略が提案された。本論で対象とする ツキノワグマ(Ursus thibetanus japonicus)は,本州以南 に生息するアンブレラ種のひとつであり,野生生物の保 全という観点からは,ツキノワグマの生息が地域生態系 の健全性の指標となり得る。一方で,人身被害,林業や 農業への被害を及ぼす点で,害獣としても近年悪名高い。 環境省の報告によれば,2006 年では全国で 4251 頭が捕 殺されている現状があり,このうち 11 県で,年間捕殺頭 数が過去最高を更新した。このように保全生物学的な観 点と社会的な観点から,重要な役割を持つ対象種であり ながら,ツキノワグマの生息地に関する知見は不足して いる。先行研究としては,餌資源としての植生と生息地 の関係性の研究(たとえば,Huygens et al., 2003 など) が主流であり,植生以外の環境情報と生息分布との関係 性に関する知見はほとんど存在しない。これまでのツキ ノワグマ生息環境推定に関する研究は,米田 (2001)によ る植生分類を用いた大まかな生息環境の推定と,坂田・ 三谷 (2002)によるツキノワグマのロジスティック回帰 モデルの紹介があるが,双方において推定結果の検証は 行われていない。坂田・三谷 (2002)は,生物科学の総合 啓蒙誌である「生物の科学 遺伝」にて,ツキノワグマ のロジスティック回帰モデルを新しい試みとして紹介し た。これは,生息環境のパラメータ(以下,環境指標と する)を用いて,ツキノワグマの生息確率を予測すると いう手法である。しかし,坂田・三谷 (2002)の記事では, フィールドデータ収集方法や年次,サンプル数や独立変 数の選択をどのような統計手法を用いて行ったのかなど が不明瞭である。また,気候・標高・植生のみによる予 測であるため,人為的な影響を考慮するモデルではない。 * 慶應義塾大学大学院 政策・メディア研究科 環境デザイン・ガバナンス専攻・(独)日本学術振興会 ** 慶應義塾大学 総合政策学部 *** 日本大学 生物資源科学部 **** 慶應義塾大学 環境情報学部
そこで,本稿では,地形,流域環境,気候,道路,植 生等の環境指標と,生息分布情報(生息域,非生息域) との関係性を,多変量解析を用いて分析し,ロジスティ ック回帰モデルによるツキノワグマの潜在的生息確率 予測モデルを構築することを目標とした。本研究の独創 的な特徴は,構築されたモデルが,自然環境だけではな く,道路からの距離などツキノワグマの生息地選択に対 する人為的な影響も考慮することができ,ツキノワグマ の生息地管理への応用が期待できる点,またそのモデル の精度を科学的根拠のある手法で検証している点であ る。 図 1 ロジスティック回帰モデル構築フロー ツキノワグマは,日本では本州・四国・九州において 分布しており,IUCN のレッドデータブックによれば 「脆弱」な絶滅危惧種と評価されている。本稿では,研 究対象を,富士箱根伊豆国立公園に指定されている富士 地域の個体群と,南アルプス地域個体群とした。富士地 域個体群は4 つの下位個体群が存在(望月ほか, 2005) し,そのうち愛鷹個体群などは,他の下位個体群からの 孤立が懸念されている(望月ほか, 2005)。 1. 研究の方法 本研究の全体の流れを図1 に示す。手順は,以下3 点の流れに大別できる。 (1) 独立変数と従属変数の GIS データ構築 (2) 統計解析とロジスティック回帰モデル構築 (3) モデルの精度検証 これらの手順は,次の項で具体的に説明する。 1.1 独立変数と従属変数のGIS データ構築 1)独立変数のGIS データ構築 野生動物の生息地は,「資源(食物,水,カバー) と環境条件(気温,降水量,捕食者や競争相手の有無 など)が組み合わさって,ある種が生存し繁殖できる ようにする場所」と定義できる(Morrison et al., 1992)。そこで,既存の文献から,ツキノワグマの生 息地選択性に影響を与えると想定される資源と環境条 件を,環境指標として,以下の5 つのカテゴリに分類
して選択した:(a) 地形(Huygens et al., 2003),(b) 水資源(動物にとって水資源へのアクセシビリティは 生命維持に必要であるという仮説から),(c) 気候 (Guisan and Theurillat (2000)),(d) 道路(道路は生 息地や移動に対して負の影響を与えているという仮説 から),(e) 植生(Huygens et al., 2003)。本研究で
使用した環境指標を表1 に示す。
地形に関する独立変数は,数値標高モデル(DEM) として,SRTM, NASA (Zone 5 to 6 in North and Zone 64 to 65 in East,90mメッシュ)を基に算出し, 2 シーンでモザイク画像を作成した。傾斜はArcMap ® 9.1 によりDEMから算出した。 水資源に関する独立変数は,(a) 地球地図日本から 得た全国の主要な河川のシェープファイルと(b) ArcMap® 9.1 の水文学ツールを用いてSRTMのDEM から算出した地域スケールの河川の流れの,2 つを組 み合わせて,調査地内の主要な流水系が抽出されるよ うに河川データを作成し,ArcMap®のSpatial Analyst
ツールを用いて,ユークリッド距離に基づき,30mの
空間分解能で,河川へのユークリッド距離を計算した。
気候に関する独立変数は,WorldClim1)から1kmの
空間分解能でESRI grid form (30 arc~seconds)で入手 した。降水量はひと月ごとのデータから年間平均降水 量を計算し,同様に,月ごとの最低気温と最高気温か ら年間最低気温と年間最高気温を計算した。 道路に関する独立変数は,数値地図25000(空間デ ータ基盤)から取得し,道路への距離は,河川の場合 と同様に計算した。この作業を,全ての道路,高速道 路,一般道,徒歩道・庭園路等,幅13m 以上の広い
表 1 独立変数の分類と単位 分類 独立変数 生息地の種類 単位 Code 標高 環境条件 m DEM 地形 傾斜 環境条件 ° SLOPE 水資源 河川への距離 資源(水) m RIVER 年間平均降水量 環境条件 mm PREP 年間最低気温 環境条件 ℃ TMIN 気候 年間最高気温 環境条件 ℃ TMAX 全て 全ての道路への距離 環境条件 m ALLROAD 高速道路への距離 環境条件 m HIGHWAY 一般道への距離 環境条件 m GENERAL 種類 徒歩道・庭園路等へ の距離 環境条件 m SMALL 幅の広い道路(13m 以 上) 環境条件 m WIDE 道 路 幅 幅の狭い道路(13m 未 満) 環境条件 m NARROW 正規化植生指数 資源(カバー) -- NDVI 植生 植生分類 (再分類し た 5 種類,再分類し た 14 種類) 資源(食物) -- VEG1, VEG2 道路(4 車線以上),幅 13m 未満の狭い道路(2 車線 以下)に対して繰り返し,「全ての道路」のみではなく, 道路の種類や幅に応じても距離を算出した。 植生に関する独立変数は,環境庁自然保護局が編集 している自然環境情報GIS中の数値植生区分図(縮尺 1: 50,000)から,57 種類ある植生群落を,望月ほか (2005)の分類に基づき,2 種類に再区分(5 種類と 14 種類)して解析に用いた。5 種類は,亜高山帯,山地 帯,低地帯,植林,その他である。14 種類は,亜高山 帯(落葉広葉樹林,常緑針葉樹林),山地帯(落葉広 葉樹林,常緑針葉樹林),低地帯(落葉広葉樹林,常 緑広葉樹林,常緑針葉樹林),植林(常緑針葉樹林, 落葉針葉樹林),その他(伐掘地,草地,河川敷ある いは裸地,耕地,造成地)に分類される。さらに,植 生に関する独立変数として正規化植生指数(NDVI) を算出した。まず,GLCFからLandsat-7 ETM+(1999 年10 月 12 日,1999 年 11 月 3 日,2000 年 11 月 8 日,2001 年 9 月 24 日に観測)を入手した。衛星画像 を最大50mの誤差でジオリファレンスし,オルソ幾何 補正したのち,モザイク画像を作成し,NDVI値を計 算した。画像処理にはErdas Imagine®を使用した。 2)ツキノワグマの生息・非生息地点データの抽出 ツキノワグマの生息・非生息地点のデータの抽出,及 び座標系の算出には,ArcMap® 9.1を用いた。ツキノワ グマの「生息」情報については,「静岡県ツキノワグマ 生息調査報告書(1998~2002年度)」(望月ほか, 2005) から,富士地域個体群と南アルプス地域個体群の生息地 点に関する地図を得た。まず,富士地域の4枚の地図と 南アルプス地域の44枚の地図をスキャンし,ジオリファ レンスした (測地系: WGS 84, 投影: UTM Zone 54)。第 一次多項式アフィン変換を用いて,ジオリファレンスし た地図画像の座標変換を行った(誤差は50m未満)。座標 変換後,ツキノワグマの771生息地点をデジタイズした (715地点が南アルプス地域個体群,56地点が富士地域
個体群)。ArcMap® 9.1のVBA機能により, XY座標系
を計算した。一方,ツキノワグマの「非生息」情報に関 しては,以下のような方法を取った。日本の哺乳類に関 しては,環境省が主だった動物のモニタリング調査をし ており,その成果は「第6回自然環境保全基礎調査 種 の多様性調査 哺乳類分布調査報告書」(環境省自然環境 局生物多様性センター, 2004)にまとめられている。こ の報告書から全国一律5 km のグリッドでツキノワグマ の生息分布地図を入手し,1978年と2003年での聞き取 り調査でどちらの年においても生息が確認できなかっ た地域を本研究では「非生息」地域とした。画像処理方 法としては,まず分布地図を,行政界を用いてジオリフ ァレンスし,「生息」地点に対する処理と同様に地図画 像の座標変換を行った。そして,生息域と非生息域を分 別する境界線をデジタイズし,「非生息域」のポリゴン を作成した。この非生息域のポリゴン内に,無作為に分 布させたポイントデータ(以下,ランダムプロットとす る)を,「非生息」地点とみなした。非生息地点とみな したポイントデータは,生息地点とほぼ同数になるよう 770地点とした。今回,回帰モデルの精度の比較に用い たAdjusted D2およびAUC等の指標は,閾値に左右され ず,定量的な値が得られるため,同数のサンプルを取っ ても,結果に影響が生じないと考えられる。 3)データ編集と学習用データ・テストデータ準備 合計で1541 地点の生息,非生息地点を得たが,研究対 象地域にプロットした際に24 地点は,研究対象地域外 であることが判明した。よって残った1517 地点を解析 に用いた。ArcMap 9.1 ®を用いて,独立関数として用意 された環境指標の地図から,各々の座標系におけるピク セル値を読み取った。ロジスティック回帰モデルの精度 検証のため,構築のための学習用データ,精度検証のた めのテストデータを準備した。一般的に,学習用データ とテストデータのセットは重複していないことが望ま しい。しかし通常は記録数が不十分であるなどの理由で 別個のデータセットを用いることは難しいのが現状で ある。しかし,本研究において,生息地点のデータは, 富士地域個体群地域ではフィールド痕跡の調査と記録, 南アルプス地域個体群地域では,ラジオ・トラッキング による4 年間の追跡調査であり,調査手法もデータ取得 日時も異なる。ゆえに,互いに完全に独立したデータセ ットとみなせるので,ランダムプロットの非生息地点デ ータ715 地点と南アルプス地域個体群の生息地点の記
録698 地点を学習用データに用い,ランダムプロットの 非生息地点データ55 地点と,富士地域個体群の生息地 点の記録49 地点をテストデータとして用いた。 1.2 統計解析とロジスティック回帰モデル構築 図2 に独立変数の選別方法を示す。手法は,1)スピア マンの順位相関係数による検定,2)カイ 2 乗検定,3)ジ ャックナイフ検定,4)箱ヒゲ図の組み合わせによる選別 である。まず,多重共線性を避けるため,スピアマンの 順位相関係数による検定を行った結果,道路に関する変 数間,標高と気温の変数間は,相関係数ρが0.85 より 大きかったため,独立変数としての併用は避けた。相関
係数の閾値は,Bonn and Schröder (2001)の手法を参考
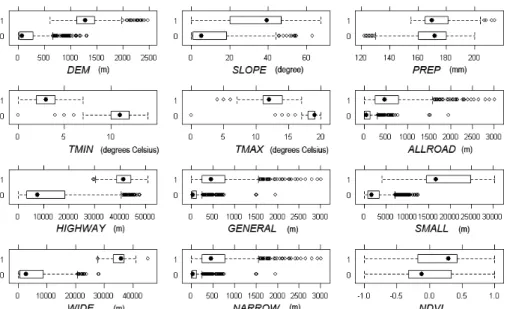
に決定した。多くの環境指標を考慮する必要性から,主 成分分析を用いることも検討したが,主成分分析は,直 接的に独立変数との関係性を示すことが困難であり,生 態学的な解釈がしづらくなるため,本研究では用いなか った。植生分類に関しては,名義尺度であるため,カイ 2乗検定を行った結果,植生分類14 種類が有意であっ た。次に,ジャックナイフ検定を行い,相対重要度が1 以上のものを選別した。最後に,量的データに対して, 箱ヒゲ図により,生息地点と非生息地点における環境指 標の分布差異を可視化した(図3)。降水量やNDVIに おける分布の違いはほとんど見られなかったものの,標 高,傾斜,気温,道路への距離などの独立変数に関して は,生息地点と非生息地点に顕著な分布の差が観察され た。標高は,700m付近が分布を分離する閾値であり, 生息地点は700m~2000mの高標高地帯に分布していた。 道路に関して,「徒歩道・庭園路等への距離」を取り上 げてみると,5 km~7.5 km付近での小さな重複があるも のの,分布は明瞭に区別され,生息地点はこの道路から 5km~30km離れた地域に分布していた。生息地点と非 生息地点との間に顕著な分布の差が見られるものを選 別し,相関関係のあった標高と気温の間で,ジャックナ イフ検定の重要度がもっとも高い変数として標高を選 んだ。これらの統計解析の結果,標高,徒歩道・庭園路 等への距離,幅の広い道路(13m以上),14 種類の植 生分類が,独立変数として選択された。 DEM SLOPE PREP TMIN TMAX HIGH WAY GENE
RAL WIDE SMALL
NARRO W NDVI 変量の対毎の散布図と 相関係数ρ 箱ひげ図 ジャックナイフ検定:相対重要度>=1 クロス集計表と カイ2乗検定 ALL ROAD VEG1 VEG2 ρ>0.85 ρ<=0.85 Correlation group I Correlation group II 有意なp 有意でないp
DEM TMIN TMAX SMALL WIDE Correlation group I
DEM
生息地点と非生息地点に 顕著な分布の差が見られる
SMALL WIDE VEG2 選別され、採択された
独立変数
DEM TMIN TMAX SMALL WIDE
1.5 1.4 1.3 1.6 1.7 VEG2 1.6 HIGH WAY 1.1 0.7 0.7 0.7 0.3 0.4 0.3 ジャックナイフ検定の重要度が もっとも高い変数 ジャックナイフ検定の数値 本研究では,ロジスティック回帰モデルの構築は,ス テップワイズ変数増加法により変数選択を行った。ロジ スティック回帰モデルのモデルとしての適正の判断は, 学習用データを用いて,AIC, adjusted D2 (Guisan and
Zimmermann ,2000), AUC (Hanley and McNeil 1982), sensitivity (Hanley and McNeil 1982), specificity(Hanley and McNeil 1982), および係数に対 する有意確率により,定量的に評価,比較した。その中 で上記の指標でもっともよい精度であったモデルを最 適モデルと名付け,最終的なモデルの精度検証を実施し た。以上の統計解析には,R version 2.4.0 を用いた。 図2 独立変数の選別方法フローチャート 1.3 モデルの精度検証 最適モデルの精度検証にはテストデータを用い,比較 のため学習用データでの精度検証も行った。定量的指標 としては,AUC と Kappa statistics を用いた。Kappa statistics では,精度は生息・非生息という 2 値変数で 評価するため,閾値に関しては,Proportion agreement の値を最大化するような値を取った。 2.結果と考察 2.1 ロジスティック回帰モデル構築 表2 は,構築されたモデルの比較を示している。Wide,
Wide + Small, Wide + DEM, Wide + Veg2 などのモデ
ルは,AUCによれば,完璧(AUC=1.00)あるいは秀 逸(1>AUC>0.9)であったが,係数に対する有意確率 (p)によれば,有意性が見られなかった(0.1<p<1)。 係数に有意性が見られないモデル(0.1<p<1)は候補か ら外し,非常に有意な確率(0<p<0.001)である係数を 持つモデルを選別した。選別されたモデルは,①DEM, ②Small,③Small + DEMの3つのモデルである。これら 3つのモデルの中では,③は,①②と比較すると,AIC, adjusted D2,AUC, sensitivity, specificityなど他
図 3 箱ヒゲ図:従属変数によって 2 グループに分類された独立変数の分布図(Y 軸の 0 は非生息,1 は生息を意味する) 表 2 ツキノワグマ生息環境予測のためのロジスティック回帰モデルの比較 「徒歩道・庭園路等への距離」との組み合わせが最適な モデルだと考えられた。 2.2 最適モデル 最適モデルは,「標高」と「徒歩道・庭園路等への 距離」を独立変数とした以下の回帰式により導かれた。 log(p/(1-p))=(-1.486e+01)+(7.335e-04)*
x
1+(9.470e- 03)*x
2. ここでx1 は「徒歩道・庭園路等への距離(m)」, x2 は「 標高(m)」である。ここで,p はツキノワグ マの生息確率を示している。表3 に,ロジスティック 回帰モデルの回帰係数を示す。切片・回帰係数は,非 常に有意であった。ここで言う,「徒歩道・庭園路等」 は,国土地理院の刊行する2 万 5 千分の 1 の地形図に 収録されている道路の種別の凡例2)に対応しており, 本研究対象地域には庭園路は存在しないため,実質的 に登山道の存在を示していると解釈される。よって, 開発されたロジスティック回帰モデルの結果は,登山 道への距離が遠ければ遠いほど,ツキノワグマの生息 確率が高まることを意味する。これは,ツキノワグマ の生息地選択に対して,登山道の存在が負の人為的影 響となっており,ツキノワグマは,登山道に対し,回 避行為を取っていることを示唆している。一方で,ツ キノワグマは標高の高い地域を選択的に利用している ことが明らかになった。ツキノワグマの食性(Huygens et al., 2003)から,餌資源となる植生分類は,生息地 選択の上で重要であると思われたが,標高のほうが植 生分類よりツキノワグマの環境選択性が強いことが判 明した。これは,植生の垂直分布で知られるように, 標高は,植生を決定付ける要因のひとつとなっており, 本研究においては,植生より標高の方が有意な環境指 標であったのだろうと考察される。また,ツキノワグ マは冬眠をする哺乳類であり,冬眠を決定する要因に モデルの名前 (Model name) AIC Adj. D2 AUC Sensitivity Specificity p(>|z|)DEM 221 0.88 0.99 0.99 0.96 0<p<0.001 Small 162 0.91 0.99 0.99 0.98 0<p<0.001 Wide 15 0.99 1.00 0.98 1.00 0.1<p<1 Veg2 479 0.77 0.97 0.93 0.96 0.1<p<1 Wide + Small 6 1.00 1.00 1.00 1.00 0.1<p<1 Wide + DEM 6 1.00 1.00 1.00 1.00 0.1<p<1 Wide + Veg2 36 1.00 1.00 1.00 1.00 0.1<p<1 Small + DEM 73 0.97 1.00 0.99 0.99 0<p<0.001 Veg2 + DEM 217 0.90 0.99 0.99 0.97 0.1<p<1 Small + Veg2 99 0.96 1.00 0.99 0.99 0.1<p<1
DEM + Small + Wide 8 1.00 1.00 1.00 1.00 0.1<p<1
DEM + Small + Veg2 61 0.98 1.00 1.00 1.00 0.1<p<1
DEM + Wide + Veg2 32 1.00 1.00 1.00 1.00 0.1<p<1
Small + Wide + Veg2 32 1.00 1.00 1.00 1.00 0.1<p<1
表 3 ロジスティック回帰モデルの回帰係数 表 4 ロジスティック回帰モデルの精度検証結果 気温が大きく関わってくると推察される。本研究の統 計解析結果も示したように,標高は,通常,気温と高 い負の相関関係にあるため,気温という環境条件も包 含している環境条件であると解釈できる。 2.3 モデルの精度検証 最適モデルの精度の検証結果を表4 に示す。テスト データの sensitivity は,specificity よりやや低め
(0.71)であった。Bias index, prevalence index は,
テスト用・学習用データともに,0 に近い値であった。 AUC に関しては,学習用データ,テスト用データと もに,0.95 以上であり卓越した精度であった。本研究 では,南アルプス地域個体群の生息確認地点を学習用 データとして,富士地域個体群の生息確認地点をテス トデータとして用いたが,モデルの精度は,非常に高 いことが検証された。これは,南アルプス地域個体群 に基づき構築したモデルの,富士地域個体群への汎用 性を示唆している。 おわりに 本研究は,ツキノワグマの生息確率予測モデルの方程 式の開発に資した。最適モデルは,次の回帰方程式で表 され,p は本種の生息確率,
x
1 は「徒歩道・庭園路等 への距離(m)」,x
2 は「 標高(m)」である。log(p/(1- p))=(-1.486e+01)+(7.335e-04)*x
1+(9.470e-03)*x
2. 本種は,害獣・保全指標種の観点から,有効な生息地管 理が期待されている。本最適モデルは,生息地管理への 応用が可能である。一方で,本種は,登山道など人為的 影響の強い地域を選択的に避けており,標高の高い地域 を選好している傾向があると推察されるため,このよう な生息選択性を考慮した上で,保全対策を立てるべきだ と思われる。今後の課題として,本モデルの,他地域の ツキノワグマ地域個体群の生息環境推定への適用可能 性を検証することが望まれる。 補注 1) WORLDCLIM (2008 updated) Worldclim.<http://www.worldclim.org/>, March 2008 referred.
2) 国土地理院 2 万 5 千分の 1 地形図の読み方・使い方.によれば, 徒歩道とは,道路幅 1.5 メートル未満の道路を指し,登山道はこれに 含まれる。一方で,庭園路とは,公園,住宅地等で自動車の通行を規 制している道路および工場等の特定の敷地内道路を指す <http://watchizu.gsi.go.jp/riyou/tizukigou/h01-01-05tohodou.htm >。国土地理院 基盤地図情報閲覧サービス.においては,これを「徒 歩道・庭園路等」と総称している <http://fgd.gsi.go.jp/view/>。 引用文献
Bonn, A. and B. Schröder (2001) Habitat Models and Their Transfer for Single and Multi Species Groups: A Case Study of Carabids in an Alluvial Forest. ECOGRAPHY 24, 483-496.
Guisan, A. and J. P. Theurillat (2000) Equilibrium Modelling of Alpine Plant Distribution and Climate Change: How Far Can We Go? Phytocoenologia, 30, 353 - 384.
Guisan, A. and N. E. Zimmermann (2000) Predictive Habitat Distribution Models in Ecology. Ecological Modelling, 135, 147-186.
Hanley, J. and B. McNeil (1982) The Meaning and Use of the Area under a Roc Curve. Radiology, 143, 29-36.
Huygens, O. C., T. Miyashita, B. Dahle, M. Carr, S. Izumiyama, T. Sugawara and H. Hayashi (2003) Diet and Feeding Habits of Asiatic Black Bears in the Northern Japanese Alps. Ursus, 14(2), 236-245.
環境省自然環境局生物多様性センター (2004). 第 6 回自然環境保全 基礎調査 種の多様性調査 哺乳類分布調査報告書, 232pp. 望月敬史・ 澤井謙二・大場孝裕 (2005) 静岡県ツキノワグマ生息調
査報告書(1998~2002 年度). 静岡県林業技術センター, 104pp. Morrison, M. L., B. G. Marcot and P. W. Mannan (1992) Wildlife-Habitat Relationships, Concepts and Applications. University of Wisconsin Press, Madison.
坂田宏志・三谷雅純 (2002) 人間の影響下での野生動物の生息状況を 予測する. 生物の科学 遺伝, 56(5), 59-64,4. 米田政明 (2001) ツキノワグマの地域個体群区分と保全管理 -生息 環境と必要な面積-. ランドスケープ研究, 64(4), 314-317. テストデータ Test data 学習用データ Train data 回帰係数 標準偏差 z 値 p(>|z|) Coefficient SD Optimized p 0.033 0.613 Kappa 0.69 0.99 Proportion agreement 0.85 0.99 Bias index -0.12 0 Prevalence index -0.17 -0.01 Sensitivity 0.7143 0.9914 Kappa statistics, sensitivity, specificity (切片) intercept
-1.486e+01 2.003e+00 -7.418 1.19e-13 ***
Small 7.335e-04 1.081e-04 6.782 1.18e-11 ***
DEM 9.470e-03 1.593e-03 5.947 2.74e-09 *** Specificity 0.9636 0.9972 AUC 0.95 1.00 AUC SE 0.1924 0.0002 z 12.856 300.767 p <0.0001 1.00000 lower 0.91 1.00 upper 0.99 1.00