起 電 性(Na,
K)-ATPase
の メカニズ ム
福 島 義 博*・ 熊 沢 紀 之**
* 生 理 学 研 究 所 能 動 輸 送 部 門 〒444 岡 崎 市 明大 寺 町
現 在 国 立 小 児 病 院 小 児 医療 研 究 セ ンタ ー 薬 理 部 門 〒154 東 京 都 世 田 谷 区 太 子 堂3丁 目35-31 ** 和 歌 山 県 立 医 科 大 学 第 一 生 理 〒640 和 歌 山市 九 番 町9
Mechanism
of Action of Electrogenic
(Na, K)-ATPase
Yoshihiro
FUKUSHIMA
* and
Noriyuki
KUMAZAWA
**
* National Institute for Physiological Sciences , Okazaki 444 ** Wakayama Medical College , Wakayama 640
Plasma
membrane
(Na, K)-ATPase
transports
3 Na + out of and 2 K+ into the cell, and thus the
pump
is electrogenic.
This
review
describes
experiments
on topical
aspects
of the
cation
movement
mechanism
of
(Na, K)-ATPase,
putting
emphasis
on
electrogenic
properties,
reconstitution,
and cation occlusion
or deocclusion
from the transport
enzyme molecule.
Key words : (Na, K)-ATPase,
Electrogenic
pump, Reconstitution,
Cation occlusion,
Amine
inhibition
1 は じめ に 2 輸 送 の化 学量 論 3 ポ ンプ 電 流 と起 電 力 の 測定 4 外 部 電 場 の 影響 5 イ オ ンの 結 合 と遊 離 6 イ オ ンの 取 り込 み部 位 7 イ オ ン輸 送 と酵 素 の リン酸 化 1. は じめ に 細 胞 膜 に お け る イオ ン輸 送 の根 幹 を 司 るナ ト リウ ム ポ ンプ,(Na, K)-ATPase, の反 応機 構 は,多 種 多 様 の 手 法 で 研 究 さ れて きた.初 期 の 電 気生 理 学 的研 究 で は,細 胞 膜 の 膜 電 位 が膜 内外 の イオ ン濃 度 差 に よ る拡 散 電 位 に 起 因 す る も の と理 解 され,ナ トリウ ム ポ ンプ の存 在 が 示 唆 され た.ナ ト リウ ム ポ ンプが 酵 素 反 応 と して認 識 され た後 は,ATP加 水 分 解 の 中 間 体 と イ オ ン移 動 の エ ネル ギ ー共 役 の 関 係 が 明 らか に され た(本 文 第4節).そ こで は,ATPに よ る リ ン酸 化 に 従 って,Na+とK+が 序 列 的 に 輸 送 さ れ るこ と,そ れ に 伴 う酵 素 の2つ の 異 な る コ ン ホ メ ー シ ョ ンが あ る こ と が 明 らか に され た.最 近 は,遺 伝 子 の 方 か ら一 次構造 も解 か れ た.一 方,細 胞 内 電解 質 組 成 を 調節 で きる パ ッチ ク ラ ンプ に よ り,膜 電 位 に 依 存 した ポ ンプ電 流 の 変化 も報 告 され て い る(本 文 第3節). さ らに,精 製 した ポ ンプ 酵 素 を リボ ソー ムや 平 面 膜 な ど の 人 工膜 に組 み 込 ん だ 再 構 成 系 も用 い られ,脂 質 と ナ ト リウ ム ポ ン プの み の 単 純 な 系 にお い て 輸 送 の 計 測 や 膜 電 位 の 作 用 が 検 討 さ れ て い る(本文 第4節).本 総 説 で は, 酵 素 反 応 の 素過 程 に膜 電 位 が どの よ うな影 響 を与 え る の か とい う,方 向性 を も った 場 に お け る ナ ト リウ ム ポ ンプ の挙 動 に は じま り(本文 第3お よび4節),酵 素 分 子 の レ ベ ルで の イ オ ン輸 送 機 構(本 文 第5か ら7節)に つ いて, ど こ ま で解 って い る の か を概 説 す る. 2. 輸 送 の化 学 量 論(Na, K)-ATPase が 起 電 的(electrogenic)に 正 味 の 陽 電 荷 を 細 胞 外 へ 輸 送 し,細 胞膜 内 側 を負 に帯 電 させ る こ とは,(Na, K)-ATPase が ポ ンプ 酵 素 と して 発 見 さ れ た1}と 同 じ年,1957年 に既 に認 識 され て い る2).そ れ は 次 の よ うな 実 験 に よ る.ま ず 低 温 に お いて 赤 血 球 内 に Na+を 負 荷 した 後,そ れ を37℃ に 移 行 し,時 間 を 追 っ て 細 胞 か ら流 出 す るNa+量 が 炎 光 分 析 で 測 定 さ れ た.
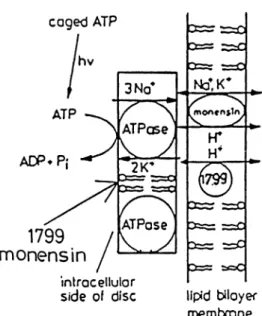
Na+流 出 には細 胞 外 にK+の 存 在 が 必 要 で あ り,こ の強 く 共 役 したNa+とK+の 流 出 ・流 入 の比(coupling ratio) が,Na+3に 対 しK+2で あ った.赤 血球 の ポ ンプ 分 子 数 は,興 奮 性 細 胞 や 腎 細胞 に くらべ て 極 め て 少 な く,ま た イ オ ン測 定 も炎 光 法 で な され て お り,サ ンプ リン グ は時 間 単 位(hour)で 行 わ れ た.こ の よ うな 当 時 に あ って, 輸 送 比 が1対1で な く1.5対1で あ る こ と を結 論 づ け た とい う こ とは や は り一 驚 に値 す る.こ の 種 の 測 定 に必 ず 伴 う非 特 異 的 な イ オ ン移 動 に よ るバ ック グ ラ ン ドの 問 題 もあ った で あ ろ う.そ の差 し引 きに は,(Na, K)-ATPase の 特異 的 阻 害 剤 と して い ち早 く取 り上 げ られ た 強 心 性 ス テ ロイド3,4)が 有 効 で あ っ た よ う だ.同 じ頃,類 似 の 実 験 でNa+とK+の 輸 送 比 を1対1と した 仕 事 もあ っ た が5),上 記 の 説 得 力 あ る仕 事 の 前 に消 え て しま った. 続 いて,や は り赤 血 球 を もち い て,ATPの 加 水 分 解 との 量 論 関 係 も求 め られ,濃 度 勾 配 の緩 急 にかか わ らず, 1分 子ATP分 解 に つ き3原 子Na"が 細 胞 外 へ,2原 子 K+が 細胞 内 へ輸 送 さ れ る こ とが 明 らか に な った6∼8).さ らに,神 経,筋 等 に お い て も結 果 は 同 じで あ る こ とが 示 さ れ た9∼12).現在 の教 科 書 の 輸 送量 論 に関 す る記 載 の ほ と ん ど は こ の辺 の仕 事 に尽 きて い る と言 って よ い,こ の よ うな事 情 の た め,Na+とK+の 輸 送 比 が 強 く固 定 され た もの で な く,条 件 に よ って 可 変 で あ ろ う と言 わ れ なが ら,(Na, K)-ATPase 分 子 の 基 本 特 性 と して の3対2 が 定 ま った か らに は,そ れ 以 上 の 興 味 を 多 く引 か ず に今 日に 至 った.む しろ その 後 の 実 験 は,3対2量 論 を さ ら に 支 持 す る もの が 多 い(例 え ば,後 述5節 参 照). 1970年 代 後 半 に は,あ る 程 度 精 製 され た腎(Na, K)-ATPaseを リボ ソー ムに 再 機 成 す る こ と が 可 能 に な っ た13).そ う な る と,放 射 性 同位 元 素22Naや86Rbを 使 い 改 め て 輸 送 比 が 測 定 され た.そ して 上 述 の 赤 血 球 に お け る測 定 結 果 が 再 構 成 系 で も再 現 され た14∼17).さ ら に 最 近,K+無 添 加 の リボ ソームの 内 外 にNa+を 与 え た 時, ATPに 依 存 性 のNa+/Na+交 換 が起 こ り,こ れ もNa+/ K+交 換 同様 に起 電 的 で あ る こ とが 示 され た18).こ の時, K+の 代 りにNa+が ご く遅 い過 程(Na+/K+交 換 の6%) で 輸 送 され て い る と考 え られ て い る. 3. ポ ンプ 電 流 と起 電 力 の測 定 ATP 1モ ル の 加 水 分 解 の 標 準 自 由 エ ネ ル ギ ーを7.3 kcalと し,こ の エ ネル ギ ーが 膜 を介 して1個 の電 荷 の移 動 に全 て 使 用 さ れ た とす る と,0.31Vと 計 算 され る.ナ トリウ ム ポ ンプ の起 電 力 を実 測 す る こ とは,こ の ポ ンプ の大 ま か な エ ネ ル ギ ー変 換 効 率 を 調 べ る こ と に もな る. ま た,電 圧 に 依 存 した ポ ンプ 電 流 の 変 化 を 調 べ る こ と に よ り,ポ ンプ 酵 素 の 反 応 回 転 に対 す る電 圧 の影 響 を知 る こ とが で き る.細 胞 膜 にお け る ポ ンプ起 電 力 を実 測 す る た め に は,細 胞 内 に電 極 を挿 入 し外 部 電 極 か ら電 圧 を 印 加 して ポ ン プ に よ り膜 を通 過 す る電 流 を計 測 す る.そ し て ポ ンプ 電 流 が 抑 制 され る電 圧(reversal potential)を 求 め れ ば よ い わ け で あ る.こ こで,計 測 に 関 して考 慮 し な けれ ば な らな い 問題 点 が あ る.す なわ ち.細胞 を もち い た 場 合,能 動 輸 送 の結 果,膜 内側 と外 側 水 相 にNa+とK+ の濃 度 勾 配 が生 じ る.そ れ に伴 い 電 気 的 中 性 を 保 つ た め に ア ニ オ ンの 移 動 も起 こ る.そ の 結 果,こ れ らの イオ ン の拡 散 電 位 に よ る膜 電 位 が形 成 され る.し た が って,細 胞 内 外 の イ オ ン組 成 を 調節 し膜 電 位 を 自 由に 変 化 させ 得 る実 験 系 を 構 築 しな けれ ば な らな い.加 えて,細 胞 膜 に は 電 位 に 依 存 して イ オ ンを 通 す 各 種 チ ャ ンネ ル タ ンパ ク が 存在 す るの で,そ れ らを 抑 制 した 状 態 で,ナ トリウ ム ポ ンプ に よ る電 流 の み を 測 定 しな けれ ば な らな い. 最 近 のGadsbyら の 実 験 で は,単 離 した モ ル モ ッ ト心 筋 細 胞 の ホ ー ル セ ル ク ラ ン プ(whole cell clamp)に よ り細 胞 内 液 を還 流 し,印 加 電 圧 を-140∼+60mV間 で 変 化 させ た 時 に膜 を 通 過 す る電 流 を 測 定 して い る19).ウ ア バ ィ ン感 受 性 の 電 流 値 を ナ ト リウ ム ポ ン プ によ る もの とみ な した 時,ポ ンプ 電 流 は 電 圧 に 依 存 して 変 化 し,膜 電 位0mV付 近 で最 大値 と なる.ま たreversal potential よ り求 め た ポ ンプ の 起 電 力 は,140mV程 度 とされ て い る. ア フ リカ ツメ ガ エ ル(Xenopus Laevis)卵 を使 った 類 似 の 測 定 で は,印 加 電 圧-200∼+100mVで 調 べ た 結 果, +20mV付 近 で ポ ン プ電 流 が 最 大 に な る と報 告 さ れ て い る20).ポ ンプ 電 流 が 膜 電 位 に 対 して 直線 的 に変 化 せ ず, 極 大 値 を 示 す こ とは,ポ ン プの 回 転 の 素 過 程(次 節4.参 照)の うちに,反 応 促 進 と抑 制 の相 反 す る電位 感 受 性 の 過 程 が あ る もの と推 定 され て い る.し か しい ず れ に しろ, この よ うな 実 験 は ポ ンプの 回転 の 全 体 に 対 す る電 位 の 影 響 を トー タ ルで 観 察 して い る にす ぎ な い.分 子 論 的 機 構 を 研 究 す るた め に は,も っ と単 純 な 再 構 成 系 に 頼 る こ と にな る(次 節4.参 照). 一 方,ト ータル としての ポンプ電流や起電力 を,脂 質 二 分 子 膜(黒 膜)等 の 再 構 成 系 を も ち い て 測 定 しよ う と す れ ば,そ れ は 非 常 に困 難 で あ る.な ぜ な ら,ポ ンプ1 分 子 が 運 ぶ 電 流 値 は,ポ ンプが 最 大 の 分 子 活 性 を 示 した と して も,約10-18A程 度 で あ り,こ の 値 は 電 流 計 測 の 限 界 値10-13Aよ りは るか に 小 さい か ら で あ る.し た
が って,少 く と も105個 以 上 の ポ ンプ が 同 時 に イオ ン輸 送 を 行 わ な い とポ ンプ電 流 と して 検 出 で きな い.ま た 通 常 の黒 膜 作 製 法 で は,膜 中 に デ カ ン等 の有 機 溶 媒 が 含 ま れ る た め ポ ンプ のATP分 解 活 性 が 阻 害 さ れ る と も言 わ れ て い る.つ ま り平 面 膜 系 で ポ ンプ電 流 を 計 測 するには, 有 機 溶 媒 を 含 ま な い脂 質 二 分 子層 に105個 以 上 の ポ ンプ 分 子 を 方 向 を そ ろ え て組 み込 み,そ れ らが 同 調 して 作 動 す る こ とが 要 求 され る.こ の よ うな条 件 を 全 て 満 足 す る 系 は ま だ 開 発 され て い な い. い ろ い ろ の工 夫 は 試 み られ て い る.Fendlerお よ び Nagelら は,精 製(Na, K)-ATPase を 含 む 脂 質二 分 子 膜 シ ー トが 負 に荷 電 して い る こ とに 着 目 し,陽 電荷 を も つ 脂 質 で 形 成 し た 黒 膜 に(Na, K)-ATPase を 吸 着 (condensar-coupling)さ せ た(Fig.1)21,22).位 相 を そ ろ え て い っせ い に ポ ンプ を 作 動 さ せ る た め に,(Na, K)-ATPaseを 含 む シ ー トの 接 す る 側 の 液 相 にcaged ATP(Fig.2)を 加 え て お き レーザ ー光 照 射 に よ りATP を 遊 離 さ せ る.こ の 時,黒 膜 を通 過 す る過 渡 的 電 流 が観 測 され て い る.同 様 の方 法 で,さ らに時 間 分 解 能 を 向上 さ せ た 実 験 も報 告 され て お り,過 渡 電 流 はATPに よ っ て 引 き起 こ され た ポ ンプ酵 素 内のNa+移 動 に よ る もの と 結 論 され て い る23,24).し か しこ の 実 験 系 で は,膜 に吸 着 す る シー ト数 が 不 明 で あ り,全 て の シ ー トが 同 一 方 向 で 吸 着 す る と い う証 拠 もな い.ま た,黒 膜 の 透 過 性 を 上 げ る た め に イ オ ノホ ア が 添 加 され て い るが,こ の イオ ノ ホ アが シ ー ト部 分 に も取 り込 ま れ る こ と は 十 分 考 え 得 る.そ うだ とす る と,シ ー トと黒膜 の 間 隔 に,(Na, K)-ATPase作 用 を十 分 に反 映 した イ オ ン濃度 勾 配 は保 持 さ れ な い と思 わ れ る(Fig.1参 照).し たが って,過 渡 電 流 後 の 定 常 電 流 は ポ ンプ電 流 の間 接 的反 映 と は見 な し得 て も,起 電 力 の定 量 的 議 論 に も ち い る に は さ らに 検 討 を要 す る と考 え られ る. 4. 外 部 電場 の影 響 上 述 した よ うに(3節,第2パ ラ グ ラ フ)膜 電 位 と ポ Fig. 1 吸 着膜 形成 の 模 式 図(Nagelら22)か ら 改 変 して 引 用).
Fig. 2 Caged ATPと 光 の反 応 に よ るATPの 遊 離 Fig. 3 (Na, K)-ATPase に よ る イ オ ン 輸 送 とATP 加 水 分 解 の 共 役 反 応 の 順 路
ン プ機 構 の分 子 論 的 対 応 は,こ れ か ら の 課 題 で あ る が25,26),こ こで は この 問 題 に 対 す る い くつ か の 実 験 を 述 べ る.そ の た め に,(Na, K)-ATPase のPost-Albers ス キ ー ム と呼 ば れて い る輸 送機 構(Fig.3)を,ま ず 簡 単 に 記 す.く わ し くは他 の 文 献 を 参 照 して 欲 しい27∼32).
細 胞 質 側 にあ るNa+お よ びATPの 各 結 合 部 位 に, Na+お よびATPが 結合 す ると,ATP7位 リン酸 基 がAsp-369に67)転 位 し共 有結 合 アシル リン酸 を形 成 し,Naは(Na, K)-ATPase内 部 に 取 り込 ま れ る33)(occlusion).い った ん この 形 態 に な る と平 衡 はNaを 細 胞 外 へ 遊 離 す る方 に 偏 り,Na+を 遊 離 し細 胞 外 液 側 か らのK+に 高 親 和 性 を もっ よ うに な る34).そ してK+が 結 合 す る と水 と リン酸 エ ス テ ル の 反 応 は 著増 し,た だ ち に 正 リ ン 酸(pi)を 細 胞 質 側 に遊 離 し,か つ 結 合Kが 酵 素 分 子 内 部 へ 取 り込 ま れ る35)(occlusion).こ こでmMオ ー ダー のATPが あ る と,取 り込 まれ たKが 細 胞 質 へ 放 出 さ れ,酵 素 は再 び 細 胞 質Na+と 結 合 で き るよ うに な る35).生 理 的 濃 度 の ATPが 存 在 す る時 は,細 胞 外へ のNa+遊 離[(Na) E1P →E2P]が ,ま たATP濃 度 が 低 い 時 は,細 胞 質 へ のK+ の遊 離[(K) E2→E1]が 各 々 律 速 段 階 に な る. ポ ンプ酵 素 は,カ チ オ ン輸 送 の 一 回 転 に上 記 の よ うな 順 路 を廻 る わ けで あ るが,電 圧 に感 受 性 を も つの は ど の 段 階 な の だ ろ う か.我 々 は こ の 問 題 を 解 く た め に, Coronadoら の 方 法 に 従 い36),パ ッチ ク ラ ン プ 用 の ピペ ッ トの先 端 に一 定 方 向 を も った(Na, K)-ATPase を 保 持 した 脂 質 二分 子 層 を作 った37).酵 素 が 方 向 性 を 保 っ て い る こ とは,ウ ア バ イ ン とATPが 反 対 側 か らの み 効 果 を 示 し,他 の 阻害 剤 バ ナ ジ ウ ム酸 はATPと 同 方 向 か ら効 くことによ り推 測 され た.さ て この よ うな状 態 の(Na, K)-ATPase の 両 側 に ほぼ 生 理 条 件 に等 しい イ オ ン組 成 を与 え 電 圧 を か け た.す る とATP非 存 在 下 で は 約40 mV以 上 の 印 加 電 圧 に よ り,阻 害 剤 感 受 性 の チ ャ ン ネ ル 開 閉 様 の コ ン ダ ク タ ンス 変 化 が 記 録 され た.と ころが,1 mM ATPを 一 方 の 液 に加 え る と,コ ン ダ ク タ ン ス変 化 に は少 な くと も約80mV印 加 が 必 要 で あ った.し か も最 小 コ ンダ ク タ ンス変 化 の値 もATP非 存 在 下 の 約1/4∼ 1/7に 減 少 した.こ の場 合,お そ ら くイオ ンは受 動 的 に動 い て い る と思 わ れ るが,そ うだ とす れ ば,高 濃 度ATP は 生 理 的 な 大 きさ の膜 電 位 に よ るNa+やK+の 漏 洩 を 抑 制 す る効 果 が あ るの か も しれ な い.Fig.3に 即 して 言 え ば,ATPが 無 い時,Na・E1〓(K) E2に よ り下 り坂 方 向 に イオ ンが 流 れ,い わ ば(Na, K)-ATPase 自 身 が 漏 洩 チ ャ ン ネ ル に もな り得 る と こ ろ を,細 胞 質 の高 濃度ATP が それ を 防 いで い る と言 え よ う.な お,脂 質 二 分 子 膜 に 再 構 成 した(Na, K)-ATPase が 受 動 的 な チ ャ ン ネル 開 閉 を示 す こ と は,黒 膜 系 に お いて も報 告 され て い る38,39). 他 方,(Na) E1P→E2P変 化 が 電 圧 感 受 性 で あ る こ と が,Karlishの グ ル ー プ に よ って 次 の よ うな 実 験 で 示 さ れ た40).彼 らはLys-501を 蛍 光 剤FITCで 標 識 化 した41) (Na, K)-ATPase を リボソ ー ム に再 構 成 した.レ ポ ー タ ーFITCの 蛍 光 強度 は(Na) E1P→E2P遷 移 に伴 って 減
少 す る こ とが 知 られ て い るの で42),こ の 減 少 速 度 に対 す る電 位 効 果 を 調 べ よ う と い う わ けで あ る.リ ボ ソ ー ム に お い て は,酵 素 分 子 は,細 胞 質 側 ドメ イ ン(そ の 中 に Lys-501は 存 在 す る)を 外 液 に 向 け て 埋 め 込 まれ て い る (inside-out).し た が って,外 液 にNa+,Mg2+お よ び ATPを 加 え れ ば 酵 素 は リン酸 化 され る(Fig.3).た だ し, Lys-501はATPの プ リン部 分 との 結 合 に必 要 な 残 基 ら し く,FITC修 飾 酵 素 はATPと 結 合 で きな い.そ こで 彼 らはATPに 代 わ って ア セ チ ル リ ン酸 を使 って酵 素 を リン酸 化 した.こ の リボ ソ ー ム 内 にK+を 負 荷 し,バ リ ノマ イ シ ンに よ り拡 散 電 位 を 作 る と,FITC蛍 光 強 度 の 減 少 速 度(ス トップ トフ ロー で 追 跡)は,拡 散 電 位 が 無 い 時 の4倍 に な って い る.ま た,ATP濃 度 が高 い 時[(Na) E1P→E2Pが 律 速]は 拡 散 電位 に よ りNa+/K+交 換 速 度 が30∼40%増 加 す るが,ATP濃 度 が μMオ ーダー で あれ ば[(K)E2→Na・E1が 律 速],拡 散 電 位 は何 の効 果 も 示 し て い な い43).す な わ ち,リ ン酸 化 状 態 で の コ ンホ メ ー シ ョ ン変 化 あ るい は3Na+の 輸 送 は電 圧 感 受 性 で あ るの に 対 し,非 リ ン酸 化 状 態 で の コ ンホ メ ー シ ョ ン 変化 あ るい は2K+の 輸 送 は電 圧 非 感 受性 とい う こ とに な る. これ らの 実 験 との 関 係 を論 じ る こ とは 難 しい が,も う 一 つ,赤 血球を交流場(10-20V/cm)に さ ら し膜 電 位 変 化 を 与 え る と,ATP非 存 在 下 で(!)Rb+が 細 胞 内 へ, しか も上 り坂 の 条 件 下 で も,輸 送 され る と い う44,45).そ こで は,Rb+の 流 入 の みが 起 こ り,逆 に流 出や,Na+ の 移 動 は起 こ らな い とされ て い る.外 部 電 場 の効 果 が,ち ょ うどATPに よ るE2P形 成 と 同 じよ うに 酵 素系 に 作 用 して い るわ けで あ る.ジ ュー ル 熱 に よ り細 胞 内 代 謝 が 促 進 しATP濃 度 が 増 加 した わ け で はな い こ と は 対 照 実 験 にお いて 示 され て い る.(Na, K)-ATPase に 必 ら ず, ATP合 成 酵 素 も含 め て膜ATPaseの 機 能 に対 して,電 場 の影 響 が 大 き い こ とは,同 じ著 者 に よ って 強調 さ れ て き た46).
5. イ オ ンの 結 合 と遊 離 こ こ ま で 述 べ て き た よ う に,イ オ ン輸 送 を外 か ら眺 め た時,起 電 性 を 含 め て い くつ か の 現 象 論 は確 立 され た と み な して よい だ ろ う.で は,ポ ンプ酵 素 は ど の よ うに し て そ れ を 行 うの だ ろ うか.輸 送 過 程 を 内 か ら眺 めて み よ う とい うわ け で あ る.例 え ば,ATP 1モ ル の 分 解 に対 す る輸 送 比3Na+対2K+(1.参 照)は,酵 素 と これ ら イ オ ンと の 結 合 比 と い う形 で 説 明 され るの だ ろ うか.こ の よ うな 問題 の 研 究 は(Na, K)-ATPase の 精 製47,48)に よ りよ うや く進 歩 を見 せ た. と は言 え,多 量 の リン脂 質 を も った(Na, K)-ATPase 標 品 において,輸 送 に特 異 的 なNa+ま た はK+の 結 合 を 十 分 な精 度 で 測 定 す る こ とは 簡 単 で はな い.山 口 と殿 村 は, ミ リポ ア フ ィ ル タ ーの 上 に ヌ ク レオ ポ ア フ ィ ル タ ー を重 ね た二 重 膜 上 に,22Na+,42K+,お よ び3H-グ ル コ ー ス と イ ンキ ュ ベ ー シ ョン した 酵 素 懸 濁 液 をの せ 濾 過 し,ミ リ ポ ア フ ィ ル タ ー 中 に 捕留 さ れ た22Na/3Hと42K/3Hを 測 定 す る こ と に よ り,結 合 比3Na+対2K+を 得 た49,50).彼 らは,過 剰 量 の2価 イ オ ン添 加 に よ り酵 素 か ら追 放 さ れ る1価 イ オ ンを ンを特 異 的 結 合 と み な した.さ らに,松 井 と誉 田 は,遠 心 沈 殿 とな る膜 標 品 に存 在 す る22Na+と 42K+の うち ,ウ アバ イン共存 によ り減少 する分 を特異 的 結 合 と み な し,や は り結 合比3対2を 得 た30,51,52).彼 ら は,ア ミノ酸 分 析 か ら求 め た分 子 量 に よ る タ ンパ ク濃 度 を 使 う こ と に よ り,(Na, K)-ATPase 1分 子 あた り, ATPと ウ アバ イ ンは各1分 子,Na+とK+が 各 々3原 子 お よ び2原 子 結 合 す る と結 論 し,輸 送 の 量論 関 係 は 結 合 比 に基 づ くこ どが 示 され た. あ るい は ま た,Fig.3に 示 した 酵 素 の コ ンホ メー シ ョ ンに依 存 した リン酸 化 反 応 を使 って,結 合 の化 学量 論 を 推 定 す る こ と もで き る.Fig.3に は 通 常 の輸 送 方 向 の み の矢 が 書 いて あ るが,試 験 管 中 で は,リ ガ ン ド条件 に よ り逆 方 向 の反 応 も有 意 に起 こ り得 る.す な わ ち逆 反 応 で は,E2にPiを 与 え ると リン酸 化(E2P形 成)が 起 こる53,54). 一 方E 1はATPに よ って の み リン酸 化 さ れ る.こ の よ う な リン酸 供与 体 と コ ンホ メ ー シ ョ ンの密 接 な 関 係 が あ る の で,例 え ばpiか ら リン酸 化 さ れ る か ど う か を 指 標 と しなが ら,コ ンホ メ ー シ ョ ンを支 配 す るNa+とK+の 力 関 係 を滴 定 す る こ とが で き る55).そ の 結 果 も や は り, Na+濃 度R倍 に 対 しK+濃 度R3/2が 相 拮 抗 す る結 果 を与 え て い る. さ ら にFig.3は,Na+とK+に 対 す る 酵 素 の 親 和 性 が 序 列 的(consecutive)に 変 化 し,そ れ に よ つてNa+とK+ の 輸 送 も 序 列 的 に 起 こ る こ と を 主 張 して い る が,誉 田 と 松 井 の 結 合 実 験 は そ れ を 支 持 し,Na+とK+が そ れ ぞ れ 細 胞 内 外 か ら 同 時 に 結 合 して 一 段 階 で(simultaneous) 交 換 さ れ る の で は な い こ と も示 し た30,51).ま た,E1P→ E2Pの 段 階 でNa+結 合 の 親 和 性 が 実 際 に 減 少 す る こ と56), E2が ら のK+遊 離 が 高 濃 度ATPで 促 進 さ れ る こ と57,58) な どFig.3の 主 張 す る 内 容 は 次 々 と 証 明 さ れ た.そ れ ら に 加 え て,Cys-964に 導 入 さ れ た 蛍 光 色 素BIPM59)に よ る コ ンホ メー シ ョン 変 化 の 追 跡 と,リ ガ ン ド と の 反 応 性 の 対 応 が,Fig.3に 基 づ い て 谷 口 の グ ル ー プ に よ っ て 詳 細 に 研 究 さ れ て い る60,61). 輸 送 イ オ ン が ポ ン プ 酵 素 内 部 に 取 り 込 ま れ た 状 態 は, Fig.3の 中 で は(K) E2や(Na) E1Pと 表 わ され て い るが, こ れ ら は,イ オ ン 交 換 樹 脂 を 通 し た 時 も は ず れ て こ な い33,63,64).文 字 通 りoccluded form,イ オ ン は 外 液 か ら 隔 離 さ れ て い る35)(4節 参 照).イ オ ン の 分 子 内 ポ ケ ッ トへ の 取 り 込 み は,可 溶 化 酵 素 で も膜 結 合 酵 素 で も全 く 同 様 に お こ る64).こ の 状 態 か ら の1価 カ チ オ ン の 遊 離 は ゆ っ く り と進 行 し,特 にK+の 遊 離(K) E2→E1が ATP 濃 度 に 依 存 す る こ と は 上 述 し た 通 りで あ る(4節 参 照). 取 り込 ま れ たNaの(Na)E1Pか らの 遊 離 は,生 理 的 条 件 に 近 い と こ ろ で は 外 的 に 制 御 さ れ な い の と対 照 を な す.Na+ Fig. 4 E2のK+-ポ ケ ッ トに 取 り 込 ま れ たRb+の 細 胞 外 液 へ の 遊 離 を 示 す モ デ ル(Forbush68)か ら 改 変 して 引 用) 生 理 的 輸 送 で は,こ の モ デ ル を 逆 に た ど り,外 液 K+が 酵 素 分 子 内 へ 取 り込 ま れ る.逆 反 応 に っ い て は5節 第3パ ラグ ラ フに 少 し記 述 が あ るが,く わ し くは文 献32,34,53,ま た は54を 参 照 され た い.
とK+の 移 動 の メ カニ ズ ム に,あ る非 対 称 性 が 存 在 す る こ と は,4節 最 後 の パ ラ グ ラ フ に述べ た実 験44,45)に も 反 映 さ れ て い る のか も しれ な い. Forbushは 最 近,新 し く考 案 した迅 速 濾 過 装 置 を もち いてK+あ るい はRb+の 遊 離 の 速 度 を実 測 した65,66).そ の 装 置 は,約10ミ リ秒 の 時 間 分解 能 を そ なえて い るので, イ オ ン交 換 法 で は で きな か ったATPに よ るK+やRb+ 放 出 の 加 速 も測 定 可 能 で あ った.彼 は,(86Rb)E2にMg2+と piを 加 えE2Pを 作 り(逆 反 応),こ の 時86Rb+が 酵 素 か ら遊 離 して くる 時間 経 過 を解 析 した66).そ の 結 果,K+ (ま た はRb+)ポ ケッ トは2つ か ら成 り,酵 素 外 液 に接 し た方 に 入 った イ オ ンか ら順 次 放 出 され る とい うモ デ ル が 提 出 され た(Fig.4).酵 素 内 側 サ ブポ ケ ッ トか ら外 液 側 サ ブ ポ ケ ッ トへ の イオ ン移 行 が 比 較 的 遅 い 過 程 で,2 番 目のRb+の 放 出 に は時 間 の お くれが 観 察 され て い る. 6. イ オ ンの 取 り込 み部 位 前 節 最 後 に述 べ た よ う な酵 素 分 子 レベ ルの メ カニ ズ ム を 解 明 す る ため に は,現 在 の と こ ろ,(Na, K)-ATPase の 立 体 構 造 に関 す る知 見 が 全 く欠 除 して い る.(Na, K)-ATPaseの 一 次 構 造 が 得 られ た こ と は67∼69)は,メ カ ニ ズ ム を解 明 す る の に欠 か せ な い情 報 とい う点 で 重 要 で は あ るが,そ れ を生 か す研 究 は これ か らで あ ろ う.複 数 個 (お そ ら くα と βの2つ)の サ ブ ユ ニ ッ トで構 成 され, そ の 合 計分 子 量 が 約150,000に もの ぼ り,加 え て膜 を貫 通 して い る酵 素(Fig.5)と あ って は,今 す ぐに 三 次 元 構 造 が 明 らか に され る望 み は 薄 い.結 晶 を 作 らず に,溶 液 中で の タ ンパ ク構 造 が 解 け る よ うに な ったNMRに お い て も,扱 え る タ ンパ ク の分 子 量 は ま だ10,000ほ どまでで あ る.ま た,(Na, K)-ATPase の 二 次 元 結 晶 の電 子 顕 微 鏡 像 の画 像 解 析 も20Aそ こそ この 分 解 能 にす ぎ な い70,71). このよ うな現 状 で は,イ オ ン取 り込 み ポ ケ ッ トを 直接 見 る こ とは極 め て 困難 で あ ろ う.ま して や,輸 送 過 程 に お け る ポ ケ ッ トの動 的 変化 を 具体 化 す る こ とは 不 可能 に 近 い. しか し,い つ の 日か 立 体 構 造 が 解 かれ た時,イ オ ン輸 送 機 構 を 具 体 的 に知 る た め に,Na+やK+が 相 互 作 用 す る ア ミ ノ酸 残 基 を調 べ て お か な けれ ば な らな い.す な わ ち,カ チ オ ン取 り込 み ポ ケ ッ トは,一 次配 列 の どの 部 分 で形 成 さ れ て い るの だ ろ うか.解 明 され た一 次 構 造 の 中 か らイ オ ンを 捕捉 す る可 能 性 の あ る配 列 を 探 し,そ れ と 同 じペ プ チ ドを 合 成 し,イ オ ン との 結 合 を 測 って み る の も興 味 あ る実 験 か も しれ な い.実 際,骨 格 筋 小 胞 体 に あ って(Na, K)-ATPase と 極 似 と言 わ れ て い る カ ル シウ ム ポ ンプ に つ いて,Ca結 合 ペ プ チ ドを こ の よ うな方 法 で 推 定 した 報 告 も 出て い る72).し か し,短 い ペ プ チ ドと して 溶 液 中 に あ る時 と,現 実 の酵 素 分 子 中 に あ る同 じ配 列 が,同 一 立 体 構 造 を と る とい う保 証 は な い.む し ろ異 る場 合 が多 く知 られ て い る73).そ もそ も結 合 部 位 が,一 次 構 造 上 で 連続 した ア ミノ酸 残 基 で構 成 され て い る と い う保 証 もな い.Ca2+結 合 能 と い う こ とだ け に 限 れ ば, カ ル シ ウ ムポ ンプ 中 の 配 列 と は無 関 係 に,既 に い くつ か の 合 成 ペ プ チ ドは 報 告 され て い る74,75).し たが って,他 の 実 験 と組 み 合 わ せ な い と,合 成 ペプチ ド実 験 の みか らは な か な か 結 論 に至 らな い よ う に思 われ る.着 目 した 残 基 配 列 に対 す る抗 体 が 及 ぼす 酵 素 作 用へ の影 響 か ら,機 能 部 位 を推 定 す る 際 もや は り同様 の 限界 が あ る.こ の 場 合 は さ らに,抗 体 分 子 自身 の大 き さ を考 え る と,少 な くと も立 体 障 害 とい う二 次 的影 響 の 可 能 性 を 除 外 しな くて は な らな い. そ うか とい って,Na+やK+の 結 合 部 位 を 探 索 す る た め の 化 学 修 飾 も末 だ 有 効 な 成 果 を 上 げて い な い.例 えば, カ チ オ ン結 合 に は カ ルボ キ シ ル基 が 関 与 して い る と の予 Fig. 5 (Na, K)-ATPase の 存 在 様 式 を 表 わ す 模 式
想 の 下 に,DCCD修 飾 が 試 み られ たが,酵 素 に 導 入 され たDCCDと 近 接 した ア ミノ基 が 分 子 内 架 橋 を 形 成 す る な どの 複 雑 な現 象が 起 こ り,DCCD結 合残 基 が カ チ オ ン 結 合 に か か わ って い る と一 意 的 に 結論 で き な い76∼78). 我 々 は あ る偶 然 か ら,プ ロ トン化1価 陽 イ オ ン型 ア ミ ン類 が,(Na, K)-ATPase の輸 送 カ チ オ ン 取 り込 み ポ ケ ッ トに入 る こ とを 見 出 した79∼81).す な わ ち,普 通 に 緩 衝 液 成 分 と して使 わ れ て い る ト リス,イ ミダ ゾー ル, あ る い は リジ ン,ア ル ギ ニ ン,ブ チ ル ア ミン等 で カ チ オ ン ポ ケ ッ トを塞 い だ酵 素 はATPase活 性 を示 さな い.失 活 力 の 強 さ は,.ア ミンのpKaつ まりプロ トン化 度 と,分 子 の 大 き さの2つ に依 存 して い る.小 さな 分 子 ほ ど酵 素 内 部 領 域 へ の 接 近 が 容 易 と考 え られ よ う.イ オ ン に くらべ て大 き な 有 機 化 合物 を カ チ オ ンポ ケ ッ トに 到 達 させ るた め に は,実 際 に は,何 らか の形 で 酵 素 の構 造 に 少 しゆ るみ を 与 え る必 要 が あ った(Fig.6).そ の た め に,稀 薄 なSDS や 塩 酸 グ アニ ジ ン,あ る い は高 濃 度 の2-メ ル カ プ トエ タ ノ ー ルやDTTを も ち い た.こ れ らの 試 薬 は そ れ の み で は酵 素 を 失 活 させ な い濃 度 で加 え て い る.お そ ら く,疎 水 性 基 を持 った ア ミンを 合 成 す れ ば(で き る だ け分 子 サ イ ズ は小 さ い 方 が よ い)タ ンパ ク 構 造 を人 為 的 に ゆ る め て や る こ とな しに,そ の ア ミン化 合物 を カ チ オ ンポ ケ ッ トへ 取 り込 ま せ る こ とが で き るだ ろ う82).ま た,イ オ ン 取 り込 み部位 残 基 を 同定 す ると い う目的 か らす れ ば,疎 水 性 ア ミンに ア ジ ド基 を 付 け,光 標 識 化 も 試 み て み た い83). 7. イ オ ン輸 送 と酵 素 の リン酸 化 ア ミンに よ る 阻 害 は,実 は 部 分 反 応,E2P加 水 分 解 (Fig.3参 照)に お いて も顕 著 で あ る84). E2Pは 既 にNa+を 外 液 に 遊 離 し 終 っ た 状 態 で あ り, Na+の 取 り込 み ポ ケ ッ トが 外 液 に向 いて 開 い た 後 と考 え Fig. 6 タ ン パ ク 不 安 定 化 剤 の 共 存 下 に お け る ト リ ス(A)ま た は ア ル ギ ニ ン(B)に よ る(Na, K)-ATPの 失 活 Fig. 7 Na遊 離 残 基 に よ る 水 分 子 活 性 化 と,ア ミ ン塩 基 ま た は高 濃度Na+に よ る水 活性 化 の妨 害 本 文7節,第2パ ラ グ ラ フ参 照. Fig. 8 イ オ ン の 結 合 部 位 と 遊 離 部 位 の 位 置 に 関 す るATPaseの コ ン ホ メ ー シ ョ ン変 化(Yamamoto ら87)か ら 引 用)
A B C られ る.こ の 時 の リン酸 基 は,Naを 取 り 込 ん で い る (Na) E1Pの リン 酸 基 よ り も水 との反 応 性 に富 む ので あ るが,EIPとE2Pは い ずれ もAsp-369が リン酸 化 され て お り,2つ の 状 態 間 で リン酸 基 の 転 位 は 認 め られ て い な い85).2つ の リン酸 エ ス テ ル 近 傍 の微 環 境 のpHや 誘 電 率 の 差 異 が お そ ら く水 感 受 性 の 違 い の 原 因 で あ ろ う79). それ に 加 え て,Na遊 離 残 基 が 水 分 子 の 活 性 化 に あず か って い る と仮 定 す る と,Na放 出 後 に は じめ て 水 と リ ン 酸 エス テ ル と の反 応 性 が 激 増 す るの が説 明 しや す い(Fig. 7).そ うす ると,リ ン酸 基 とNa+遊 離 残 基 は 極 く接 近 して 存 在 しな けれ ば な らな いが,少 な くと もNa結 合 部 位 と リ ン酸 基 は10A程 度 し か 離 れ て い な い と の 予 測 が あ る86).結 合 部 位 と遊 離 部 位 は 同 じ残 基 の立 体化 学 的 差 異(コ ン フ ィギ ュ レー シ ョ ン変化)で は な か ろ う か (Fig.7).リ ン酸 基 は 細 胞 質 領 域(Fig.5参 照)に 存 在 す る こ と が わ か って い るの で,上 の仮 定 を 認 め る と,Na+ 遊 離 は膜 を 越 え た 反 対 側 で 起 こ るの で は な く,酵 素 分 子 内 の 細 胞 質 ドメ イ ンで す で に 起 こ って い る こ と に な る (Fig.8C). プ ロ トン化 ア ミンは,こ のNa遊 離 残 基 に作 用 し水 の 活 性 化 を 妨 げ るの で はな い だ ろ うか(Fig.7).事 実,高 濃 のNa+自 身 も同 じ よ うにE2P水 解 を 阻 害 す るし84,88), Na+非 存 在 下 で はNa+に 代 わ って ア ミ ン類 がATPか ら
の リン酸化 ま で も触 媒 す る89)など,プ ロ トン化 ア ミン と Na+の 類 似 性 が 示 唆 され て い る. Na遊 離 部 位 は外 液 に開 いて い るの で,ア ミン(あ る い はNa+)阻 害 は 添 加 後 た だ ち に表 わ れ,実 際 に は氷 中 の反 応で 秒 単 位 で 観 測 され る.そ れ に 対 し,6節 で 述 べ た 非 リン酸化 状 態での カチオン取 り込 み部位 への 影 響 は,氷 中 に お いて 数 十 分 の単 位 で ゆ っ く りと,し か もタ ンパ ク 構 造 不 安 定 化 剤 の 共 存下 に は じめ て あ らわ れ て くる.取 り込 み 部位 と遊 離 部 位 に 対 す るア ミン接 近 度 の 違 いが わ か る(Fig.9). Na+とK+の 輸 送 機 構 に は,あ る非 対 称 性 が あ る と書 い た(5節)が,Na+遊 離 は リン酸 化 酵 素 で 起 こ り,K+遊 難 は非 リン酸 化 酵 素 で 起 こ る(Fig.3)し,結 合量 論 も等 し くな い(2お よび5節).Na+とK+の キ レー ト試 薬 との 結 合定 数 を見 渡 して み る と,ど ん な キ レー ト剤 で もNa+ の方 を よ り強 く結 合 す る.ア ミノ酸 残 基 と カチ オ ンの 相 互 作 用 に お け る 非 対 称 性 に,こ うい う こ と も何 か関 係 し て い るの だ ろ うか90).ま た,酵 素 へ の リ ン酸 基 の 導 入 は,酵 素 分 子 に どん な 変 化 を与 え てNa遊 離 へ 導 くの だ ろ う.今 日,リ ン酸 化 が タ ンパ ク の機 能 発 現 に重 要 で あ る例 は 多 岐 に渡 って 記 載 さ れ て い るが,リ ン酸 化 に よ る タ ンパ ク分 子 の変 化 に,何 か 共通 の原 理 が 存 在 す るの だ ろ うか.Lipmanに よ る"高 エ ネ ル ギ ー リン 酸 結 合"概 念 の提 唱か ら久 し いが,分 子 レベ ルで の 具 体 的 理 解 が 望 まれ る. 引用 文 献
1) J. C. Skou : Biochim.
Biophys.
Acta, 23, 394
(1957)
2) R. L. Post,
P. C. Jolly : Biochim.
Biophys.
Acta, 25, 118 (1957)
3) H.-J. Schatzmann
: Helv. Physiol.
Acta,
11,
346 (1953)
4) T. Akera : Science, 198, 569 (1977)
5) I. M. Glynn : J. Physiol.,
134, 278 (1956)
6) A. K. Sen, R. L. Post : J. Biol. Chem.,
239,
345 (1964)
7) R. Whittam,
M. E. Ager : Biochem. J., 97, 214
(1965)
8) P. J. Garrahan,
I. M , Glynn : J. Physiol., 192,
217 (1967)
9) P. F. Baker : J. Physiol.,
180, 383 (1965)
10) M. Dydynska,
E. J. Harris : J. Physiol.,
182,
92 (1966)
11) S. L. Bonting,
L. L. Caravaggio
: Arch.
Bio-chem. Biophys., 95, 416 (1963)
12) L. Beauge : Curr. Tot. Membranes
Transport,
22, 131 (1984)
13) L. E. Hokin : "Membrane
Transport
Processes"
(D. C. Tosteson,
Yu. A. Ovchinnikov,
R.
Latorre,
ed.) p. 399, Raven Press, New York
(1987)
14) K. J. Sweadner,
S. M. Goldin : J. Biol. Chem.,
250, 4022 (1975)
15) S. Hilden, L. E. Hokin : J. Biol.
Chem.,
250,
6296 (1975)
16) S. M. Goldin : J. Biol. Chem., 252, 5630 (1977)
Fig. 9 カチ オ ン取 り込 み 部 位 と,遊 離 部 位 に対 す る,1価 陽 イオ ン型 ア ミ ンの 接 近 しや す さの 違 い
17) B. M. Anner,
L. K. Lane, A. Schwartz,
B. J.
R. Pitts : Biochim.
Biophys.
Acta, 467, 340
(1977)
18) F. Cornelius,
J. C. Skou : Biochim.
Biophys.
Acta, 904, 353 (1987)
19) D. C. Gadsby,
J. Kimura,
A. Noma :
Nature,
315, 63 (1985)
20) A. V. Lafaire,
W. Schwartz
: J.
Membr. Biol.
91, 43 (1986)
21) K. Fendler,
E. Grell, M. Haubs,
E. Bamberg:
EMBO J., 4, 3079 (1985)
22) G. Nagel,
K. Fendler,
E. Grell, E. Bamberg
:
Biochim. Biophys. Acta, 901, 239 (1987)
23) R. Borlinghaus,
H. J. Apell,
P. Lauger : J.
Membr.
Biol., 97, 161 (1987)
24) H. J. Apell,
R. Borlinghaus,
P. Lauger : J.
Membr. Biol., 97, 179 (1987)
25) P. DeWeer : "Electrogenic
Transport,
Funda-mental
Principles
and
Physiological
Impli-cations"
(M. P. Blaustein,
M. Lieberman
ed.)
p. 1, Raven Press, New York (1984)
26) P. Lauger
: Biochim.
Biophys. Acta,
779, 307
(1984)
27) 谷 口 和 弥:生 物 物 理,21,11(1981)
28) L. C. Cantley : Curr.
Top. Bioenerg.
11, 201
(1981)
29) 福 島 義 博,中 尾 真:日 本 臨 床,40,197(1982)
30) H. Matsui : "Transport
and Bioenergetics
in
Biomembranes"
(R. Sato, Y. Kagawa
ed.)
p.
165, Japan
Scientific
Societies Press,
Tokyo
(1982)
31) 福 島 義 博:蛋 白 質 核 酸 酵 素,28,1131(1983)
32) 福 島 義 博:"バ イ オ エ ナ ー ジ ェ テ ィ ク ス"(中 尾 真 編)p.95,学 会 出 版 セ ン タ ー,東 京(1986)
33) I. M. Glynn, Y. Hara,
D. E. Richard
: J.
Phy-siol., 351, 531 (1984)
34) K. Taniguchi,
R. L. Post : J. Biol. Chem., 250,
3010 (1975)
35) R. L. Post,
C. Hegyvary,
S. Kume : J.
Biol.
Chem., 247, 6530 (1972)
36) R. Coronado,
R. Latore : Biophys. J., 43, 231
(1983)
37) N. Kumazawa,
T. Tsujimoto,
Y. Fukushima
:
Biochem. Biophys.
Res. Commun.,
136,
767
(1986)
38) T. A. Last, M. L. Gantzer,
C. D. Tyler : J. Biol.
Chem., 258, 2399 (1983)
39) R. Reinhardt,
B. Lindemann,
B. M. Anner
:Bio-chim. Biophys. Acta, 774, 147 (1984)
40) A. Rephaeli,
D. E. Richards,
S. J. D. Karlish
:
J. Biol. Chem., 261, 12437 (1986)
41) R. A. Farley,
C. M. Tran,
C. T. Carilli,
D.
Hawke,
J. E. Shively : J.
Biol. Chem.,
259,
9532 (1984)
42) C. Hegyvary,
P. L. Jorgensen
: J. Biol. Chem.,
256, 6296 (1981)
43) S. J. D. Karlish,
A. Rephaeli,
W. D. Stein : " The
Sodium Pump"
(I. M. Glynn, J. C. Ellory ed. )
p. 487, The Company
of Biologist,
Cambridge
(1985)
44) E. H. Serpersu,
T. Y. Tsong : J. Membr. Biol.,
74, 191 (1983)
45) E. H. Serpersu,
T. Y. Tsong : J.
Biol.
Chem.,
259, 7155 (1984)
46) T. Y. Tsong, R. D. Astumian :
Bioenereg.
15,
457 (1986)
47) P. L. Jorgensen,
J. C. Skou, L. P. Solomonson:
Biochim.
Biophys. Acta, 233, 381 (1971)
48) P. L. Jorgensen
: Methods
Enzymol.,
32, 277
(1975)
49) M. Yamaguchi,
Y. Tonomura
:
J. Biochem.,
86, 509 (1979)
50) M. Yamaguchi,
Y. Tonomura
:
J.
Biochem.,
88, 1365 (1980)
51) H. Matsui, H. Homareda
: J. Biochem., 92, 193
(1982)
52) H. Homareda,
H. Matsui : J. Biochem., 92, 219
(1982)
53) R. L. Post, G. Toda, F. N. Rogers :
J.
Biol.
Chem., 250, 691 (1975)
54) R. L. Post, G. Toda, S. Kume, K. Taniguchi : J.
Supramol.
Struct.,
3, 479 (1975)
55) R. L. Post :
"Biochemistry
of
Membrane
Transport"
(G. Semenza, E. Carafori
ed.)
p.
352, Springer-Verlag,
Berlin (1977)
56) M. Yamaguchi,
Y. Tonomura
: J. Biochem., 88,
1387 (1980)
57) M. Yamaguchi,
Y. Tonomura
: J. Biochem., 88,
1377 (1980)
58) H. Homareda,
T. Nozaki, H. Matsui : J.
Bio-chem., 101, 789 (1987)
59) M. Nagai,
K. Taniguchi,
K. Kagawa,
H.
Matsuo,
S. Nakamura,
S. Iida :
J.
Biol.
Chem., 261, 13197 (1986)
60) K. Taniguchi,
K. Suzuki,
D. Kai, I. Matsuoka,
K. Tomita,
S. Iida : J. Biol. Chem., 259, 15228
(1984)
61) K. Taniguchi,
K. Suzuki,
T. Sasaki,
H,
Shimokobe,
S. Iida : J. Biol. Chem., 261, 3272
(1986)
62) I. M. Glynn, D. E. Richards
: J. Physiol.,
330,
17 (1982)
63) I. M. Glynn, J. L. Howland.
D. E. Richards : J.
Physiol.,
368, 453 (1985)
64) B. Vilsen, J. P. Andersen,
J. Petersen,
P. L.
Jorgensen
: J. Biol. Chem., 262, 10511 (1987)
65) B. Forbush : J. Biol. Chem., 262, 11104 (1987)
66) B. Forbush : J. Biol. Chem., 262, 11116 (1987)
67) G. E. Shull, A. Schwartz,
J. B. Lingrel : Nature,
316, 691 (1985)
68) K. Kawakami,
S. Noguchi,
M. Noda,
H.
Takahashi,
T. Ohta,
M. Kawamura,
H.
Nojima, K. Nagano,
T. Hirose, S. Inayama,
H.
Hayashida,
T. Miyata,
S. Numa : Nature,
316,
733 (1985)
69) M. M. Shull, J. B. Lingrel : Proc.
Natl.
Acad.
Sci. USA, 84, 4039 (1987)
70) H. Hebert,
P. Jorgensen,
E. Skriver,
A. B.
Maunsbach
: Biochim.' Biophys. Acta, 689, 571
(1982)
71) H. Hebert,
E. Skriver,
R. Hegerl,
A. B.
Maunsbach
: J. Ultrastru.
Res., 92, 28 (1985)
72) P. Gangola,
A. E. Shamoo : J.
Biol.
Chem.,
261, 8601 (1986)
73) W. Kabsch, C. Sander : Proc. Natl. Acad. Sci.
USA, 81, 1957 (1984)
74) M. Calapietro,
P. DeSantis, A. Palleschi :
Bio-Polymers, 25, 2227 (1986)
75) C. M. Deber, D. A. Lannigan : Biophys. J., 41,
380a (1983)
76) C. H. Pedemonte,
J. Kaplan : J.
Biol.
Chem.,
261, 3632 (1986)
77) C. H. Pedemonte,
J. Kaplan : J.
Biol.
Chem.,
261, 16660 (1986)
78) F. R. Gorga : Biochemistry,
24, 6783 (1985)
79) 福 島義 博,篠 原 康 雄:生 体 の科 学,印 刷 中(1987)
80) Y. Fukushima
:
"Na, K-ATPase"
(A. B.
Maunsbach
ed.) in press, Alan R. Liss, Inc.,
New York (1987)
81) 福 島義 博:生 体 エ ネ ル ギ ー 研 究 会 第13回 討 論 会 要 旨 集,164,(1987)
82) 日置善 雄,高 田準 治(萬 有 製 薬株 式 会 社):私 信 83) 長 宗 秀 明(徳 島大 学,歯 学 部):私 信
84) Y. Fukushima
: J.
Biol.
Chem.,
262, 11000
(1987)
85) R. L. Post, S. Kume, T. Tobin, B. Orcutt,
A.K.
Sen : .J. Gen. Physiol.,
54, 306s (1969)
86) S. E. O'Connor,
C. M. Grisham : Biochemistry,
18, 2315 (1979)
87) T. Yamamoto,
H. Takizawa,
Y. Tonomura
:
Curr. Tot. Bioenerg.,
9, 179 (1979)
88) F. M. A. H. Schuurmans
Stekhoven,
H. G. P.
Swart,
J. J. H. H. M. dePont,
S. L. Bonting :
Biochim.
Biophys. Acta, 855, 375 (1986)
89) F. M. A. H. Schuurmans
Stekhoven,
H. G. P.
Swart,
J. J. H. H. M. dePont,
S. L. Bonting :
Biochim. Biophys.
Acta, 815, 16 (1985)
90) 江 橋節 郎(生 理 学 研 究 所):私 信 書 評: 膜 と 神 経 ・ 筋 ・ シ ナ プ ス2 堀 田 健 ・田 中 亮 (名古 屋 市 立 大 学 医 学 部) 編 集 1987,Xii+466 page, A5 判,定 価6,800円,喜 多 見書 房,東 京 書 評 を 書 く よ う依頼 され て,こ の 本 を読 ん だ.読 み 通 して み る と予 想 外 にお も し ろ く,有 益 な 本 だ と思 った. とに か く,よ くも これ だ け 多 くの 事 項 を 盛 り込 め た もの と感 心 した.し た が って,こ の 本 は膜 につ い て 体 系 的 に ま とめ た も ので は ない.し い て 云 えば 「寄 せ 鍋 」 的 で あ り,そ して そ れが,読 み や す さ と気 安 さを 読 者 に与 え る の で あ る.す な わ ち,ど こか らつ っつ いて も味 わ え る. 生 体 膜 の研 究 とい えば,-と りわ け従 来 の 「膜 学 会 」 の 中 で は,「 透 過 性 」 と い う コ トバが,意 識 の 中心 に な って い るよ うに見 え るが,こ の本 は,そ れ が 如 何 に 認 識 不 足 で あ るか とい う こ とを教 えて くれ よ う. こ の 本 は,ま え が きに も述 べ られ て い る よ う に,「 生 体 を情 報 シス テ ム と して と らえ,そ の な か で の膜 の 果 た す役 割 り と意 義 の 展 開」 とい う こ とを 方針 と して 編 集 さ れ て い る とい う.体 系 的 と はいえ な い に して も,そ のmind は と りあ げ られ た 項 目か ら もよ く伝 わ って く る.内 容 は 4編 か ら構 成 され,生 体 膜 の 生 理 学 的,生 化 学 的,形 態 学 的 な基 本 的 性 質 か らは じま り,シ ナ プ ス,心 筋 や 平 滑 筋 を含 む筋 肉 まで 及 び,エ ネル ギ ー変 換 系 と して の 膜, ア セ チ ル コ リ ン受 容 体,イ オ ンチ ャ ネ ル,筋 細 胞 に お け る興 奮-収 縮 連 関 で のcharge movement説 な ど,膜 研 究 の 前 線 ま で わ か りや す く述 べ られ て い る. した が って,生 体 膜 に 関 して は 専 門 で な い人―― 膜 学 会 で は 非生 物 系 の 人 ―― や 学 生 た ち に と って は 絶好 の 入 門書 に な る と同 時 に,現 代 の 生 体 膜 研 究 の 動 向 を 知 る う え で も好 著 で あ る. た だ,我 田 引 水 的 な評 に な るが,評 者 が 専 門 と して い る膜 電 位 の 光 学 的 測 定 に つ いて は 明 らか に 誤 解 と思 わ れ る説 明 が な され て い るが(第II編 の2,第IV編 の1),こ れ に つ いて は,本 誌6(2),93-103(1981)の 綜 説 を 参 考 に い ただ け た らと思 う. 評 を終 わ る に あ た って,こ の よ うな ユ ニ ー クな 解 説 書 を編 集 され た堀 田 健,田 中 亮 両教 授 に敬 意 を表 した い. (東 京 医 科 歯 科 大 学 医 学 部 第 二 生 理 神 野耕 太 郎)