814 油 化 学
総
説
微細 藻 類脂質 とその応用
松 永
是 ・中 村
徳 幸
東京農工大学工学部生命工学科
(〒184東 京都小金井市 中町2-24-16 )
Microalgal Lipid and Its Application
Tadashi MATSUNAGA and Noriyuki NAKAMURA
Department of Biotechnology , Tokyo University of Agriculture and Technology (24-16,Nakacho-2 , Koganei-shi , Tokyo ,〒184 )
Docosahexaenoic
acid (DHA)
and icosapentaenoic
acid (EPA)
are valuable
nutrients
be
-cause they have important
roles as the constituents
of structural
lipids in the nervous tissues of
human
brain and retina.
In addition,
DHA, EPA and other omega-3 fatty acids are important
factors in the prevention of several human diseases.
Fishes have been the dietary source of these
fatty acids and they originate
from the marine microbes
upon which the fishes feed. Therefore ,
these marine
microbes
cultured
under
control
could be utilized
as alternative
source of the
omega-3 fatty acids .
Marine microalgae
are rich in DHA and EPA. However, very few species of freshwater
al
-gae contain
significant
amounts
of these compounds.
The marine
unicellular
alga Isochrysis
galbana
is well known as a source of such fatty acids and this species has been used extensively
as a food source for aquaculture.
The advantage
of the marine
microalgae
such as I. galbana
over other microbes as sources of polyunsaturated
fatty acid is that this alga may be grown pho
-tosynthetically
using carbon dioxide as the sole carbon source and seawater
as a source of min
-erals .
In this paper, we focused on the production
of marine
microalgae
as an alternative
source of
polyunsaturated
fatty acid. We have screened a number
of species of marine
microalga
specifi
-cally for their ability to produce DHA and EPA and have optimized the growth conditions
for I .
galbana.
The utilization
of microalgae
as feed for cultivation
of rotifers
is also reported.
More
-over, the applications
of genetic engineering
for EPA production
are also demonstrated
.
1 緒 言 DHA (ド コ サ ヘ キ サ エ ン酸), EPA (イ コサ ペ ンタ エ ン酸), GLA (γ-リノ レン酸) な どの 高度 不 飽和 脂 肪 酸 は,生 体 の代 謝 生 理 に大 き く関 与 し,生 体 の 正 常 な機 能 発 現 に重 要 な働 き を有 して い る と考 え られ て お り, そ の 薬 理 作 用 な どにつ い て研 究 が 進 め られ て い る 。近 年, 高 齢 化 が 進 む わ が 国 に お いて,食 生 活 の 欧米 化 に伴 い血 栓 症 を ベ ー ス に して発 症 す る脳 こ うそ く(梗塞)な ど の疾 患 が 増 え て い る が,そ れ ら を 予 防 す る 物 質 と して, DHAや EPAが 注 目を 浴 び て い る1)∼3)。 これ ま で に DHA の 薬 理 作 用 と して は コ レス テ ロー ル 及 び 中 性脂 肪 低 下 作 用,制 が ん 作用, 抗 ア レルギ ー作 用,学 習 機 能 向 上 作 用,健 脳 作 用, さ らに老 人 性 痴 ほ う(呆)症 な どに対 す る効 果 が 示 され て い る 。現 在,こ れ らの 高 度 不 飽和 脂 肪 酸 は,化 学 合成 が 困難 な た め天 然 物 か ら摂 取 しな くて はな らな い 。 これ らの高 度 不 飽 和 脂 肪 酸 は魚 油 に 多 く含 まれ て い るが,そ の起 源 は魚 の え さで あ る微 細 藻類 や プ ラ ン ク トンな どの海 洋 微 生 物 に 由来 す る こ とが知 られ て い る。 海 洋 微生 物 の 中 で も微 細 藻 類 は,太 陽光 と海 水 中 の 栄 養 塩 及 び二 酸 化 炭 素(CO2) で生 教 で き,そ の 速 度 は大 型 藻類 や魚 介 類 な ど に比 べ て 早 く,大 量培 養 が可 能 な た め,微 細 藻 類 を 用 い た 有 用 物 質 生 産 に 関 す る 研 究 が, マ リ ンバ イオ テ ク ノ ロ ジー の 一 分 野 と して注 目を 集 連絡者:中 村徳幸
め て い る。 本 稿 で は, 海 洋 微 細 藻 類 を利 用 した DHA の効 率 的 な生 産 とDHA 高 含 有 微 細 藻 類 の 水 産 し(餌) 料 へ の利 用, さ らに 海 洋微 細 藻 類 に お け る有 用 物 質生 産 を 目 的 と した 不 飽 和脂 肪 酸合 成 遺 伝 子 の ク ロー ニ ング に つ いて 述 べ る。 2 海 洋 微 細 藻 類 に よ る DHA 生 産 2・1 DHA 高 含 有 微 細 藻 類 の ス ク リー ニ ン グ ま ず,薬 理 機 能 が注 目され て い る高 度 不飽 和 脂 肪 酸 の 含 量 を 指 標 に,各 地 の 微 細 藻 類 の カ ル チ ャー コ レク シ ョ ンか ら得 られ た株 を培 養 し,ス ク リー ニ ング を行 っ た。 そ の 結 果, 藻 体 中 のDHA 含 量 は 真 核 藻 類 の ハ プ ト藻 に 属 す るIsochrysis galbana が 他 の 株 に比 べ て 最 も高 い こ とが 明 らか に な っ た4)。 ま た この 他 に も,EPA 含 量 が 高 いChaetoceros gracilisや, この2つ の脂 肪 酸 を 比 較 的 多 く有 す る Pavlova lutheriが 見 い だ され た (Table-1)。 こ れ ら の 脂 肪 酸 が 局 在 す る脂 質 の 種 類 は,そ の生 理 作 用 及 び 利 用 法 を考 え る上 で重 要 で あ る。 I. galbanaの 含 有 す る脂 質 を 薄 層 ク ロ マ トグ ラ フ ィー で調 べ た と ころ,DHA は主 に リ ン脂 質 中 に存 在 し, そ の他 の 中性 脂 質 に も含 ま れ て い る こ とが 分 か った 。 さ ら に 培 養 時 の 温 度 に つ い て も検 討 を 行 っ た 。 培 養 温 度 10∼35℃ の 範 囲 でI. galbana のDHA 生 産 を藻 体 のバ イ オ マ ス 量 と 藻 体 中 の DHA 含 量 か ら比 較 した 。 温 度 に関 して は脂 肪 酸不 飽 和 化 酵 素 が 低 い 温 度 で働 く とい う 報 告 が あ る5)。 この 実 験 で も DHA 含 量 は17℃ と比 較 的低 い 温 度 で, 15mg/g dry wt. と高 くな るが,藻 体 のバ イ オ マ ス量 が 低 く,実 際 のDHA 生 産 を 考 慮 す る と25℃ で の 培養 が適 切 で あ っ た。 また,世 界 各地 の海 よ り微 細 藻 類 を 分 離 し,大 量 培 養 可 能 な 株 に つ い て,そ の脂 肪 酸 組 成 につ い て 検討 した。 海 洋 緑 藻 Chlorella sp. NKG 042401 の 主 な 脂 肪 酸 と して は リノ ー ル 酸 とパ ル ミチ ン酸 で あ り,α-リ ノ レ ン 酸 と γ-リノ レ ン酸 も藻 体 内 に蓄 積 して い る こ と 6), 及 び 糖 脂 質 中 の37.9% が γ-リノ レ ン酸 で あ る こ とが 示 さ れ て い る7)。 ま た,ス トロ マ トラ イ トを 形 成 して い る と 考 え られ る黄 褐 色 の 藻 類 の 脂 肪 酸 組 成 を調 べ た と こ ろ, I. galbana よ り も DHA含 量 が 高 い 株,及 び 五 gal -bana と 同程 度 のDHA を 含 み しか もEPA 含量 が 非 常 に 高 い株 が得 られ,い ず れ も単細 胞 でべ ん(鞭)毛 を 持 つ 遊 泳 可 能 な もの で あ った 。 細 胞 の形 態 な ど に よ り前 者 は 渦 鞭 毛 藻 のGyrodinium sp. と,後 者 は ハ プ ト藻 の Pavlova sp. とそ れ ぞ れ 同 定 され た 。 これ ら新 た に分 離 した株 の生 教 を 比 較 した と こ ろ, Pavlova sp. が生 育, バ イオ マ ス量 と もに 高 い値 を示 した。 そ こで,最 適培 養 条 件 で のバ イ オ マ ス, DHA及 びEPA生 成 をI. gal -bana で の生 成 と比 較 した 。I. galbanaの1d当 た りの バ イ オ マ ス 生 成 は 0.42g/L で あ るの に 対 し,Pavlova sp. で は1.7 倍 の 0.72g/L で あ っ た。 この と きのDHA 生 成 は,I. galbanaで5.2mg/L/d, Pavlova sp. で
816 油 化 学 4.8mg/L/d とほ ぼ 同 等 の 値 で あ った 。 EPA 生 成 につ い て は, I.galbana で は検 出 され な か った た め, 含 ま れ て い て も微 量 で あ る と推 察 さ れ る が,Pavlova sp. で は11.3mg/L/d と な った。 以 上 の こ とに よ り高 度 不 飽 和 脂 肪 酸 の 生 成 に 関 して は Pavlova sp.の 方 が I.gal -bana の 約3倍 高 い こ とが 明 らか にな った 。 2・2 フ ォ トバ イ オ リア クタ ー を 用 い たI. galbana に よ る DHA 生産 微 細 藻 類 の 培 養 を制 御 す る条 件 と して,光,CO2, 温 度,培 地 成 分 等 が 挙 げ られ る 。 これ らの要 因を 最 適 化 す る こ と に よ って,そ の 微 細 藻類 が もつ 能 力,特 に光 合 成 能 を充 分 に発 揮 させ る こ とが可 能 に な る。 中で も光 は微 細 藻 類 の光 合 成 に不 可 欠 な 唯一 の エ ネ ル ギ ー源 で あ り, そ の供 給 方 法 は微 細 藻 類 の培 養 に お いて 最 も重 要 な 因 子 で あ る と考 え られ る。 藻体 の懸 濁 液 に外 部 か ら光 を 照 射 す る と,藻 体 に光 が 遮 られ,培 養 液 の 内部 まで 光 が 充 分 に供 給 され な い とい う問題 点 が あ っ た。 微 細 藻 類 を 効 率 よ く培 養 す るた め に は,培 養 液 内部 に充 分 に光 を 供 給 す る必 要 が あ る。 培 養 液 内部 の光 強 度 を均 一 に し,か つ, 培 養 に必 要 な 光 を 充 分 に供 給 可 能 な フ ォ トバ イ オ リア ク タ ー を作 製 した8)。 この フ ォ トバ イ オ リア クタ ー は, 側 面 か ら光 が 分 散 す る 処理 を した光 フ ァイ バ ー を 挿 入 して い る。 この フ ォ トバ イ オ リア ク タ ーを 用 い,培 養 液 内 の 光 環 境 を最 適 化 す る こ と に よ り,海 洋 藍 藻 を10g/L 以 上 の 高 密 度 ま で培 養 可能 な こ とが 報 告 され て い る9)。 こ の フ ォ トバ イ オ リア ク タ ー を利 用 して I. galbana に よ るDHA 生 産 を試 み た 。そ の 結 果,リ ア ク タ ー 内 に 挿 入 した 光 フ ァ イバ ー の 表 面 の 光 強 度 が 4.5μE/m2/ s の 時,1d 当 た りの 藻 体 増 殖 量 は0.36g/Lと 最 大 と な り, これ まで の培 養 法 に よ る生 産 量 の 約3倍 で あ った4)。 培 養 液 中 の 無 機 炭 素 濃 度 が減 少 す る と,I. galbana 細 胞 へ のCO2 の 供 給 が 律 速 とな り生 教 速 度 が 減 少 す る。 フ ォ トバ イオ リア クタ ー でI. galbana を 培 養 す る と,培 養 液 のpH は9.5 に ま で達 す る。 この 場 合, リア ク ター へ のCO2 の 供 給 は空 気 を800mL/min で通 気 す る こ と に よ って 行 った が,こ の 程 度 のCO2供 給 で はI. galbanaが 利 用 で き るCO2が 不 足 す る た め,pHが 上 昇 す る もの と考 え られ た 。 そ こで,I. galbanaの CO 2 固 定 量 に 合 わせ て,培 地 のpHを 指標 に して,CO2 を供 給 す る こ と と した。こ の方 法 に よれ ば,過 度 のCO2 の 供 給 に よ る培 養液 のpH の低 下 が 起 こ らず に,I. galbana の 生 育 に 適 したpH 範 囲 で 容 易 に 制御 す る こと が で き る。 この よ うな シ ス テ ム を用 いてpH 制 御 下 で培 養 を行 っ た と こ ろ,DHA生 産 量 はpHを コ ン トロー ル しな い 場 合 の4.3mg/L/d か ら5.2mg/L/d に増 大 した 。 2・3 暗処 理 ・低 温 処 理 に よ る DHA 生 産 の 促 進 微 細 藻類 の脂 肪 酸 組 成 は培 養 条 件 に左 右 さ れ る こ とが 示 され て い る10),11)。例 え ば,藍 藻Spirulina platen -sisに お い て 培 養 液 を一 時 的 に 暗 所 に置 くと,藻 体 重 量 当 た りの γ-リノ レ ン酸 の 含 量 が増 え た と い う報 告 が あ る6)。 ま た,紅 藻Polysiphonia coactaに お い て 低 温 で 培 養 す る と, EPAの 含 量 が 増 加 す る こ とが 報 告 さ れ て い る12)。 DHAの 生 産 性 を 上 げ る た め に は,藻 体 の バ イ オ マ ス 量,藻 体 中 のDHA含 量 を 上 げ る こ と も必 要 で あ る が,こ れ らの 報 告 を 基 に,生 教 した細 胞 に対 す る各 種 の 処 理 (暗 処 理 ・低 温 処 理)を 試 み た。 そ の 結 果,低 温 (15℃) 処 理 及 び 暗 処 理 を 同 時 に行 う こ と に よ って, DHA 含 有 量 を12h の 処理 で1.7 倍 に増 加 させ る こ とが で き た。 3 海 洋 微 細 藻 類 か ら の DHA 抽 出 及 び 精 製 DHA の 精 製 に は ク ロ マ トグ ラ フ ィ ー法,尿 素 付 加 法,銀 担 持 粘 土 鉱 物 を 用 い た 方 法 な ど様 々 な手 法 が あ る が,時 間 と コ ス トの 面 や 食 品 に 添加 す る とい う点 で 難 点 が指 摘 され て いた 。 筆 者 らはI. galbana か ら有 機 溶 媒 を用 い た 簡 単 な 抽 出方 法 を確 立 した13)。凍 結 乾 燥 したI. galbana を ク ロ ロ ホ ル ム と メ タ ノ ー ル の 混 合 溶 媒(容 量 比,2:1) に よ って 抽 出 し,そ の 後 ア ル カ リけ ん 化 した サ ン プ ル を n- ヘ キ サ ンやn- ヘ プ タ ン によ って 精 製 した結 果,濃 厚 硝 酸 銀 溶 液 を 用 い た 方 法 よ り高 い 回収 率 が得 られ た 。 こ の結 果 よ り,安 全 で 簡 易 な 方 法 に よ るDHA の 精 製 が 可 能 とな った 。 4 DHA 含 有 海 洋 微 細 藻 類 の ワ ム シ餌 料 へ の 応 用 DHA を 含 有 す る微 細 藻 類 の 直 接 的 な 利 用 方 法 と し て,養 殖 で の じ(餌)料 が挙 げ られ る。 海 産 魚 の 種 苗 生 産 で は 稚 魚 の 生 存 率 を 高 め る こ と が 重 要 な 点 で あ る 。 DHAやEPA な どの 高 度 不 飽 和 脂 肪 酸 が生 存 率 を 高 め る こ とが 明 らか に な った後,稚 魚 の 初 期 餌 料 と して用 い られ る ワム シな どの不 飽 和 脂 肪 酸 に よ る栄 養 強 化 が行 わ れ て い る14)。 ワ ム シの 培 養 に は ク ロ レラ や 酵 母 が 利 用 され て い る。 ま た, I. galbana は二 枚 貝 の 幼 生 や 海 産 魚 の稚 魚 の 餌 料 と して 利 用 さ れ て い る。 そ こ で 筆 者 ら は,DHA含 量 の 高 いI. galbanaに 着 目 し,ワ ム シ用 餌 料 へ の 応 用 を試 み た 。 I. galbana を二 次 培 養 に使 用 した 場 合 の脂 肪 酸 組 成 を 調 べ た。Chlorella regularis で ワム シを 増 殖 させ て か ら(一 次 培養), I. galbana を ワ ム シに与 え た。 そ の 結 果,二 次 培 養 開始 か ら24h 後 にDHA 含 量 が 4.3 % にな り,高 度不 飽 和 脂 肪 酸含 量 も10.3% に達 した(Ta -ble-2)。 この よ うにI. galbana を二 次 培 養 に使 用 す る こ とに よ り効 率 よ く餌 料 価 値 の 高 い ワム シを生 産 す る こ とが で き た 。
5 海 洋 微 細 藻 類 の 遺 伝 子 組 み 換 え と 不 飽 和 脂 肪 酸 合 成 遺 伝 子 の ク ロ ー ニ ン グ 微 細 藻類 の 中で も遺 伝 子 組 み 換 え 系 が発 達 して い るの は,原 核 藻類 で高 等 植 物 と同 様 な 光 合成 系 を有 す る藍 藻 (シ ア ノバ ク テ リア)で あ る。 最 近 の マ リ ンバ イ オ テ ク ノ ロ ジー の発 展 に伴 い,海 洋 微 細 藻 類 か らの有 用 物 質 生 産 が 注 目を集 め て い る の と 同時 に,遺 伝 子工 学 の適 用 に よ る 生 産 性 の 向 上 が 必 要 な 課 題 と な って き て い る。 実 際,筆 者 らの 研 究 室 で世 界 各 地 よ り分 離 した 海洋 藍 藻 か らは 植 物 成 長 制 御 物 質15)や,紫 外 線(UV-A) 吸 収 物 質16),多 糖17),酵 素 チ ロ シ ナ ー ゼ 阻 害 物 質 等 多 くの有 用 物 質 が 発 見 され て い る 。 しか しなが ら,こ れ ら海 洋 か ら分 離 した有 用 物 質 生 成 株 の ほ とん ど は 自然 形 質 転 換株 で は な い ため,遺 伝 子 導 入技 術 を適(応し,そ の 技 術 が 確 立す る こ と に よ り物 質 生 産 へ の展 開 が期 待 で き る。 そ こ で こ こで は,海 洋 藍 藻 で の 遺伝 子 組 み換 え,及 び 不 飽 和 脂肪 酸合 成 遺 伝 子 の ク ロー ニ ング につ い て紹 介 す る。 5・1 海 洋藍 藻 の 遺 伝 子 組 み 換 え 海 洋 藍 藻 に お け る遺 伝 子 操 作 系 は 後 述 す る接 合 伝 達 法 も含 め,ま だ始 ま っ た ばか りで あ る。 これ ま で に報 告 さ れ て い た 例 と して は 自然 形 質 転 換 能 力 を 有す る海 洋 藍 藻
Agmenelum quadruplicatum PR 6, Synechococ -cus sp. PCC 7002 に お いて の み で あ った18),19)。 この 系 で は,導 入 した い 遺 伝 子 と細胞 を混 合 す るだ けで 細 胞 がDNAを 取 り込 み,ゲ ノ ムへ の 組 み 換 え あ る い は プ ラス ミ ドの複 製 が 行 わ れ て い る 。A. quadruplicatum PR 6に お い て も内 存性 の プ ラス ミ ドが 発 見 され,こ れ を 用 い た大 腸 菌 と の シ ャ トル ベ クタ ー 系 を用 い外 来 性 タ ンパ ク質 の 発 現 が 試 み られ て い る。 例 え ば, Bacillus thuringiensis の 産 生 す る 殺 虫 タ ンパ ク 質 を A. qua -druplicatum に お い て 発 現 させ る試 み が な さ れ て い る20)。 しか しな が ら,こ れ ま で の と こ ろ海 洋 藍 藻 に お いて は遺 伝 子 操 作 が行 え る系 は数 株 に限 定 され た もの で あ り,今 後 マ リンバ イ オ テ クノ ロ ジ ーが 益 々発 展 して い く中 で続 々 と ス ク リー ニ ング さ れて くる有 用 物 質 生 産 能 や特 殊 能 力 を有 す る海 洋 藍 藻 につ い て対 処 で き る遺 伝 子 操 作 技 術 を 開発 す る こ とは 急 務 で あ る。 筆 者 ら は,海 洋 藍 藻 Synechococcus sp. NKBG 042902に つ い て 遺 伝 子 操 作 系 の 開 発 を 試 み た21)。 ま ず,こ の株 に お いて 形 質 転 換 系 に 用 い る た め の シ ャ トル ベ クタ ー に利 用 で き る プ ラ ス ミ ドの 存在 に つ いて 検 討 し た 。 そ の 結 果,4種 類 の プ ラ ス ミ ド pSY 08(>10kb) , pSY 09(∼10kb), pSY 10(2.7kb) 及 び pSY 11 (2.3 kb)が 見つ か っ た。 この 中で 比 較 的 コ ピー数 が高 く,分 子 量 の 小 さいpSY 11を 用 い て 大 腸 菌 との シ ャ トル ベ ク タ ー を構 築 した 。pSY 11はFig.-1に 示 す よ うな 制 限 酵 素 サ イ トを有 し,コ ピ ー数 は1ゲ ノ ム あ た り30∼ 50 の 高 コ ピー プ ラ ス ミ ドで あ った 。 こ のpSY11の Hin dIIIの1.4kb 断 片 をpUC 18の マ ル チ ク ロー ニ ング サ イ トに導 入 し,ハ イ ブ リ ッ ドプ ラ ス ミ ドpUSY 02を 作 成 し た。 一 方, ホ ス ト株 と して はSynechococcus sp . NKBG 042902自 身 がpSY 11を 有 す る こ とか ら不 和 合 性 に よ る シ ャ トル ベ ク タ ーの 排 除 が懸 念 さ れ た ため, 同 株 の キ ュ ア リ ング株 で あ るYG 1116を 作 成 した 。 この 株 は042902株 を ア ク リジ ンオ レ ンジ処 理 し,任 意 に選 択 した コ ロニ ーか ら得 られ た キ ュア リ ン グ株 で pSY 10 及 びpSY11を 欠 損 して い る。 本 株 は淡 水 藍 藻Ana -cystis nidulans等 の様 な顕 著 なDNA取 り込 み 能 力 は 有 して い な い こ とか ら,大 腸 菌 にお け る形 質 転換 時 の様 に,細 胞 を コ ン ピテ ン ト化 させ る こ とが 必 要 で あ った 。 こ の結 果,構 築 した シ ャ トルベ ク タ ーpUSY 02を 用 い て 海 洋 藍 藻Synechococcus sp. NKBG 042902を 形 質 転 換 で き る こ とが 明 らか とな った。 ま た,こ の シ ャ トル ベ ク ター はA. nidulans R 2を 同 様 に 形 質 転 換 で き る こ と が 示 さ れ た 。 しか しな が ら,こ れ ま でA. nidu -lans R 2で 用 い られ て き た シ ャ トル ベ ク タ ー プ ラ ス ミ ドpECAN 8を 用 い て 海 洋 藍 藻 を 形 質 転 換 す る こ と は で き な か った。この こ と は,本 シ ャ トル ベ ク タ ーが 淡 水, 海 洋 藍 藻 の両 藍 藻 に広 く用 い る こ とが で き る可 能 性 を 示 Table - 2 Fatty acid composition and content of
the L-type retifer B. plicatilis initially cultured with C. regularis and secon -dary cultured with I. galbana for 1 day .
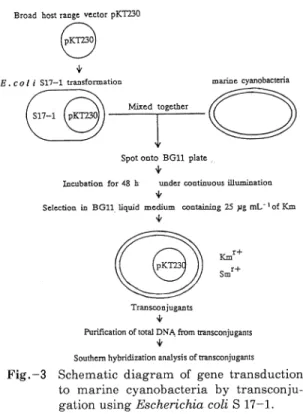
818 油 化 学 して い る と と もに,海 洋 藍 藻 の た め の独 自の 遺 伝 子 操 作 系 の 開発 の 必 要 性 が 改 め て強 調 され た結 果 とな った 。 一 方 ,最 近注 目されてきている形質転換技術の一つに パ ーテ ィ クル ガ ン(マ イ ク ロ プ ロ ジ ェ ク タイ ル)法 が あ る。 こ の 方 法 は導 入 した い 遺 伝 子 や プ ラス ミ ドで コ ー テ ィ ング した 金 属 微 粒 子 (DNAキ ャ リヤ ー)を 高 圧 圧 縮 空 気 や 火薬 を用 い て高 速 で 打 ち 出 し,こ れ を 直接 目的 とす る 細 胞 に 打 ち 込 み 形 質 転 換 させ る とい う方 法 で あ る。 この 方法 に よ って,植 物 細 胞 を プ ロ トプ ラ ス ト化 せ ず に 形 質 転換 が で き る よ う にな った ほ か,細 胞 中 のオ ル ガ ネ ラ に DNAを 導 入 す る こ と も可 能 とな っ た 。 著 者 らは,海 洋 藍 藻 の 形 質 転 換 の 効 率,操 作 性 の 向上 を 目指 し,パ ー テ ィ ク ル ガ ン法 の応 用 を 試 み た22)。 こ れ ま で パ ー テ ィ ク ル ガ ン法 で 用 い ら れ て い た DNAキ ャ リ ヤ ー は タ ング ステ ンや 金 微粒 子 で あ った が,そ の 直 径 は 0.1∼0.5μm 程 度 と大 き い た め に原 核 生 物 の 形 質 転 換 用 と して は不 向 きで あ った 。 そ こ で,磁 性 細 菌 が 細 胞 内 に 合 成 す る磁 気 微 粒 子 (磁 性 細 菌 粒 子)を DNAキ ャ リ ヤ ー と して 応 用 す る こ とを 試 み た 。 磁 性 細 菌 粒 子 を γ -APTES, グ ル タ ル ア ル デ ヒ ド及 び トリア ミ ンで 修 飾 す る こ と に よ りDNAの 固 定 化 量 が2μg/mg Particle と,従 来 用 い られ て き た DNAキ ャ リヤ ー の 倍 以 上 の DNA を 吸着 固定 す る こ とが で き た。 この磁 性 細 菌粒 子 DNA キ ャ リヤ ー を用 い て,高 圧 縮 空 気 式 パ ー テ ィク ル ガ ン法 に よ る 海 洋 藍 藻 Synechococcus sp.と 淡 水 藍 藻 Anabaena sp. PCC 7120の 形 質 転 換 を 試 み た 。 そ の結 果,圧 力100kgf/cm2 に よ って 射 出 した と こ ろ海 洋 藍 藻 で は最 大1.5%,淡 水 藍 藻 で は最 大7.5% の細 胞 が磁 気 応 答 性 を示 し, DNAキ ャ リヤ ー が 細 胞 内 に極 め て 高 い 効 率 で 導入 で き た こ とが 明 らか とな った 。 こ の実 験 に お い て は,海 洋 藍 藻 で は上 述 の シ ャ トル ベ クタ ー pUSY 02が, 淡 水 藍 藻 で は 同 様 の ベ ク タ ーpRBL 5が DNA キ ャ リヤ ー に 固定 され て い るに もか か わ らず,こ の 条 件 で は形 質 転 換 体 が 得 られ な か った 。 これ らの 磁 気 応 答 性 を示 す 細 胞 を 透 過 型 電 子 顕 微鏡 に よ り観 察 した と こ ろ, い ず れ も膜 近 傍 に DNAキ ャ リヤ ー が 存 在 して い る の が確 認 され た23)。 従 って 今 後, DNAキ ャ リヤ ー の射 出 速 度 を 上 昇 させ る こ とに よ って,形 質 転 換 が 行 え る もの と期 待 され る。海 洋藍 藻 の多 くは表 面 が 多 糖 で被 覆 さ れ て いた り,あ る い は そ れ らを洗 浄 す る と凝 集 ま た は増 殖 しな くな って しま う細 胞 も多 い 。 従 って,こ の よ うに単 純 に形 質 転 換 が 行 え な い よ う な 海 洋 藍 藻 に お い て は, パ ー テ ィ クル ガ ン法 は有 力 な 手 段 とな る で あ ろ う。 最 近, 著 者 ら は前 述 の 海 洋 藍 藻Synechococcus sp . 中 の プ ラ ス ミ ドの 一 つ, pSY 10の 興 味 深 い性 質 に つ い て 報 告 した24)。 これ ま で 述 べ て き た シ ャ トル ベ クタ ー pUSY 02は 同株 の 高 コ ピ ー プ ラ ス ミ ドpSY 11を も と に作 製 した 。pSY 11の コ ピー数 は培 養 条 件 に よ らず 一 定 の コ ピー 数 が 確 認 さ れ て い る が,pSY 10は 通 常 の 培 養 条 件 で はpSY 11と 同等 の コ ピ ー数 を 呈 す るが,培 養 条 件 に よ って 飛 躍 的 に そ の コ ピー数 が増 加 す る。 この 株 を あ らか じめ 塩 濃 度 の 低 い 培地 で培 養 した場 合 は, pSY 10の コ ピー数 は ゲ ノ ム あ た りpSY 11と 同 様30∼50 程 度 で あ る が,こ れ を 塩 濃 度 が 高 い 培 地 に 移 す と塩 濃 度 0.5∼2% で は コ ピー 数 は150,3% で は250程 度 ま で 20h以 内 に急 増 す る(Fig.-2)。 この よ うな現 象 は pSY 11を 含 め 同 株 の 他 の3つ の プ ラ ス ミ ドで は全 く観 察 さ れ な い 。 ま た, こ の よ う なpSY 10の コ ピー 数 の 増 加 は,塩 化 ナ トリウ ム と 同様 に浸 透 圧 や イ オ ン強 度 の ス ト レス を与 え る こ とが で き る塩 化 カ リウ ム や ソル ビ トール を 添 加 した 状 態 で は 見 られ な か っ た。 以 上 の こ とか ら, pSY 10は 塩 化 ナ トリウ ム の濃 度 変 化 に 対 して 選 択 的 に 応 答 し,コ ピ ー数 が 急 増 す る こ と が 明 らか と な った 。 従 って,こ の こ とを 利 用 す る こ とに よ り塩 濃 度 に よ って コ ピー数 が コ ン トロー ル で き る シ ャ トルベ ク タ ーが 構 築 で き る と期 待 され る。 これ は外 来 遣 伝 子 発 現 シス テ ム を 開発 す る際,制 御 が 可 能 な プ ロ モ ー タ ー と と もに 有 力 な 手 段 とな るで あ ろ う。 さ らに,よ り多 くの 種類 の海 洋 藍 藻 へ 効 率 的 に 遺 伝 子 を導 入 す る技 術 の 開発 を 目的 と して,海 洋 藍 藻 に対 して 接 合 伝 達 法 に よ る遺 伝 子 操 作 を 行 った25)。 供 与 菌 と してR. Simonら が1983年 に発 表 したE . coli S 17-1 株 を 用 い て い る26)。 この 株 はゲ ノ ム 中 に プ Fig. - 1 Restriction map of pSY 11 and construc
ラ ス ミ ドRP 4が 組 み込 まれ て い る。 従 って こ の 株 に RP 4が 接 合 伝 達 時 に 特 異 的 に認 識 す るMob サ イ トを 有 す る プ ラ ス ミ ドあ る い はmob, bom を 有 す る プ ラ ス ミ ドが 形 質転 換 さ れ れ ば他 の ヘ ル パ ー プ ラス ミ ドや カ ー ゴ プ ラス ミ ドを用 い な い で高 効 率 で 接 合 伝 達 が行 え る。 この プ ロ セ ス をFig.-3 に示 す 。 ま ずTn 5が 組 み 込 ま れ て い るsuicide vectorプ ラ ス ミ ドpSUP 1021を 海 洋 藍 藻 に接 合 伝 達 に よ って 導 入 す る こ と を試 み た。pSUP 1021の レプ リコ ンは pACYC 184由 来 で あ り,大 腸 菌 以 外 で は ほ とん ど複 製 さ れ な い。 従 って,接 合 伝 達体 は Tn 5が 受 容 菌 の ゲ ノ ム 中 に組 み 込 まれ, 同 トラ ンス ポ ゾ ンが コ ー ドす るカ ナ マ イ シ ン耐 性 の 発 現 に よ って 確 認 さ れ る。 す な わ ち,こ の 系 で は 接 合 時 に お い て 複 製 が 伴 って受 容 菌 に この プ ラス ミ ドが導 入 され るが,一 た ん ゲ ノ ム に組 み 込 まれ た 後 は プ ラ ス ミ ドの不 安 定 さや 種 間 の相 違 に よ る レプ リコ ンの 差 を考 慮 せ ず,接 合 伝 達 法 の 有効 性 が評 価 で き る。 この 実験 で は海 洋 藍 藻 と して 以 下 の7 株 を 用 い た 。 単 細 胞 性 海 洋 藍 藻Synechococcus sp. 5株,Synechocystis 1株, 糸 状 性 海 洋 藍 藻Pseu -danabaena 1株 を 受 容 菌 と して, E. coli S 17-1を 供 与 菌 に 用 い, プ ラス ミ ドpSUP 1021の 接 合伝 達 を 行 っ た 。 そ の結 果,接 合 を 行 った7株 す べ て の細 胞 にお いて カ ナ マ イ シ ン耐 性 を 有 す る株 が 高頻 度 で 得 られ た。 海 洋 藍 藻 に お い て接 合 伝 達 に よ り遺 伝 子 導 入 が行 われ たの は 本 研 究 が 初 め て で あ り,ま た こ の方 法 に よ り種 々の Tn 5挿 入 失 活 変 異 株 が 作 製 で き る こ とか ら,今 後 の 海 洋 藍 藻 の遺 伝 子 解 析 に大 い に応 用 で き る と期 待 さ れ る。 さ らに,こ の研 究 に用 い た海 洋 藍 藻 の 中 で最 も増 殖 の 早 か っ たSynechococcus sp. を 用 い て, 広 宿 域 ベ ク タ ーpKT 230の 接 合 伝 達 に よ る 導 入 が 試 み られ た。 pKT 230はRSF 1010の レ プ リコ ンを 有 す るInc Q 群 の 広 宿 域 ベ クタ ー で あ り,カ ナマ イ シ ン, ス トレプ トマ イ シ ン耐 性 が コ ー ドされ て い る。 同 様 にE. coli S 17-1 を 用 い 接 合 を 行 った と ころ,カ ナ マ イ シ ン とス トレプ ト マ イ シ ン両 耐 性の 表 現 形 を 有 す る株 が10-5∼10-4 (接 合 体/受 容 菌)の 頻 度 で 得 られ た。 この 広 宿 域 ベ ク タ ー は安 定 に 回収 され, 再 びE. coliに 戻 す こ と がで き る こ と か ら,海 洋 藍 藻 とE. coliの シ ャ トルベ ク タ ー と して 充 分 利 用 で き る こ とが わ か った。 同様 に海 洋 藍 藻Syne -chococcus sp., Synechocystis sp.,補 色 適 応 能 力 を 有 す る 海 洋 藍 藻Synechococcus ATCC 29403 (PCC 7335)に お いて も,接 合 に よ るpKT 230の 導 入 に成 功 して い る。 以 上 の 結 果 は,接 合伝 達 法 が 広 く海 洋 藍 藻 に 適 用 で き る可 能 性を 示 して い る。 5・2 海 洋 藍藻 に お け る遺 伝 子 組 み 換 え 技 術 を 用 い た EPA, DHA 生 産 海 洋 藍 藻 で の 外 来 遺 伝 子 の発 現 は,前 述 した よ う に 広 域 宿 主 ベ ク タ ーpKT 230に ク ロ ラ ム フ ェニ コー ル ト ラ ンス フ ェ ラー ゼ(CAT) 遺 伝 子 を 導 入 した プ ラ ス ミ ド pKCAT を 用 い たCAT の発 現 に よ り,す で に確 認 して い る 。次 に,藍 藻 で は合 成 で きな いEPAやDHA な ど の 有 用物 質生 産 を 目的 と して,そ れ らの遺 伝 子 の ク ロ ー Fig. - 2 Effect of sodium chloride concen
-tration on pSY 10 copy number .
Fig. - 3 Schematic
diagram
of gene transduction
to marine
cyanobacteria
by transconju
-gation using Escherichia
coli S 17-1 .
820 油 化 学
ニ ング を行 って い る。
す で に矢 澤 ら に よ って 単 離 され たEPA 合 成 遺 伝 子 を 用 い て 海 洋藍 藻 内 で の発 現 につ いて 検 討 した 。 全長 約 38 kbpの EPA遺 伝 子 群 を 含 む ベ クタ ー(pEPA) を 制 限 酵 素Sau 3 AI を 用 い て 部 分 消 化 し,海 洋 藍 藻 syne -chococcus sp. NKBG 042902内 で 複 製 可 能 な こ とが す
で に 確 認 さ れ て い る コ ス ミ ドベ ク タ ーpJRD 215と の 連 結 物 で あ る コ ンカ テ マ ー を作 製 した。 これ をin vitro パ ッケ ー ジ ン グ後, E. coli NM 554 に 導 入 し,カ ナマ イ シ ン(25μg/mL) に よ って 形 質 転 換 体 を 選 択 した 。 全EPA 合 成 遺 伝 子 群 の ク ロ ーニ ング の 確 認 はE. coli の 形 質 転 換 体 よ り抽 出 した プ ラス ミ ドの 制 限 酵 素 (Xba I,Spe I)に よ る 切 断 パ タ ー ン とEPA 合 成 の 確 認 に よ って 行 っ た 。 得 られ たE. coliの 形 質 転 換 体 の20 コ ロニ ー 中, 3コ ロニ ー に お い て 全EPA 合 成 遺 伝 子 群 が クロ ー ニ ング さ れ て い た 。得 られ た3株 で は乾 燥 菌 体 1 g当 た り,お よ そ3 mgの EPAが 生 成 され て い た 。 ク ロ ー ニ ング され た全EPA 合 成 遺 伝 子 群 を接 合 伝 達 法 に よ り海 洋 藍 藻Synechococcus sp. NKBG 042902へ 導 入 し,E. coliと 同 様 に カ ナ マ イ シ ン に よ って 形 質 転 換 体 を 選 択 した。 こ の 海 洋 藍 藻 の 生 教 速 度 が早 い29℃ の 条 件 で 接 合伝 達 体 と野 生 株 を そ れ ぞ れ 培 養 し,そ の脂 肪 酸 を ガ ス クロ マ トグ ラ フ ィ ーで 測 定 した と ころ,野 生 株 で は 検 出 され な か ったEPA 及 び そ の前 駆 体 と考 え られ る ア ラ キ ジ ン酸 の 合 成 が 確 認 され た 。 以 上 の よ う に遺 伝 子 組 み 換 え 技 術 を適 用 す る こ とで , EPA の 合 成 が 海 洋 藍 藻 で 可 能 な こ と が 示 さ れ た 。今 後,適 切 な プ ロモ ー ター の検 索 す る こと で生 成 の 増 大 が 期 待 され る。 ま た,前 述 したI. galbanaか らの DHA 遺 伝 子 の 単 離 も試 み られ て お り,将 来 的 に はDHA , EPAを 同 時 に しか も効 率 的 に 生 産 す る遺 伝 子 組 み 換 え 海 洋 藍 藻 の 取 得 が 可能 に な る と考 え て い る。 6 お わ り に DHAやEPA の 高 度 不 飽 和 脂 肪 酸 は,そ の 薬 理 効 果 か ら健康 食 品 や製 薬 原 料 と して 期 待 され て い る。 しか し な が ら,現 在 魚 類 か らの 粗 抽 出物 は,に お い を有 す る交 雑物 が混 在 しや す く,精 製 が 必 要 で あ る 。微 細 藻 類 を 利 用す る ことで,こ れ らの 欠 点 を 克服 で き,食 品等 へ の 添 加 も充 分 可 能 にな る と考 え られ る 。 さ らに,遺 伝 子 組 み 換 え技 術 の適 用 に よ って,藍 藻 で のDHAや EPAの 生 産 も可 能 とな り,タ ンパ ク質 や ビタ ミ ン等 を 摂 取 す るの と 同 時 に これ ら高 度 不 飽和 脂 肪 酸 を得 る食 料 と して も期 待 され る。 〔平 成7年(1995年)4月 12日 受理 〕 文 献 1) 松永 是, 岩本 香, ブ レイ ンテクノニ ュース, 43, 5 (1994)
2) 松 永 是, 竹 山 春 子, New Food Industry, 37 , 6 (1995 )
3) 矢 澤 一 良, Petrotech.,15 (9), 844 (1992 )
4) J.G. Burgess, K. Iwamoto, Y. Miura , H. Takano , T. Matsunaga, Appl. Microbiol. Biotechnol ., 39 , 456 (1993)
5) R.W. Eppley, R.W. Holmes, J.D.H. Strickland , J . Exp. Mar. Biol. Ecol., 1, 191 (1967)
6) M. Hirano, H. Mori, Y. Miura, N. Matsunaga, N . Nakamura, T. Matsunaga, Appl. Biochem . Biotechnol., 24/25, 183 (1990)
7) Y. Miura, K. Sode, N. Nakamura, N. Matsunaga , T. Matsunaga, FEMS Microbiol. Lett., 107, 163 (1993)
8) T. Matsunaga, H. Takeyama, H. Sudo, N. Oya -ma, S. Ariura, H. Takano, M. Hirano, J.G . Burgess, K. Sode, N. Nakamura, Appl. Biochem . Biotechnol., 28/29, 157 (1991)
9) H. Takano, H. Takeyama, N. Nakamura, K. Sode, J.G. Burgess, E. Manabe, M. Hirano, T . Matsunaga, Appl. Biochem. Biotechnol., 34/35 , 449 (1992)
10) M. Pirreck, P. Pohl , Phytochemistry, 23, 217 (1984)
11) M. Tsuzuki, E. Ohnuma, N. Sato, T. Takaku , A . Kawaguchi, Plant Physiol., 93, 851 (1990)
12) R.H. Al-Hasan, F.M. Hantask , S.S. Radwan, Ap -pl. Biochem. Biotechnol., 35, 530 (1991)
13) M. Kiffe, K. Nokihara, T. Matsunaga , J. Mar . Biotechnol., in press
14) T. Takeuchi, T. Arakawa, S. Satoh , T. Watanabe , Nippon Suisan Gakkaishi, 58, 707 (1992)
15) H. Wake, H. Umetsu, Y. Ozeki , K. Shimomura , T. Matsunaga, Plant Cell Reports, 9 , 655 (1991) 16) T. Matsunaga, J.G. Burgess, N. Yamada , K. Ko
-matsu, S. Yoshida, Y. Wachi, Appl. Microbiol . Biotechnol., 39, 250 (1993)
17) H. Sudo, J.G. Burgess, H. Takemasa, N. Naka -mura, T. Matsunaga, Curr. Microbiol., 30, 219 (1995)
18) E. Stevens Jr., R.D. Porter, Proc. Natl. Acad . Sci. USA, 77, 6052 (1980)
19) E. Essich, E. Stevens Jr., R.D. Porter, J. Bacteri -ol., 172, 1916 (1990)
20) C. Angsuthanasombat, S. Panym, Appl. Environ . Microbiol., 55, 2428 (1989)
21) T. Matsunaga, H. Takeyama, N. Nakamura, Ap -pl. Biochem. Biotehnol., 24/25, 151 (1990)
22) 竹 山春 子, 工 藤 聰 子, 早 出 広 司 ,中 村 徳 幸, 松 永 是, 高分 子 論 文 集, 48 ,319(1991 )
23) H. Takeyama, C. Nakamura, A. Yamazawa , T . Matsunaga, Biotechnol. Techniques , in press 24) H. Takeyama, J.G. Burgess, H. Sudo , K. Sode, T .
Matsunaga, FEMS Microbiol. Lett., 90, 95 (1991) 25) K. Sode, M. Tatara, H. Takeyama, J.G. Burgess ,
T. Matsunaga, Appl. Microbiol. Biotechnol. , 37 , 369 (1992)
26) R. Simon, U. Priefer, A. Puhler, Bio/Technology , Nov., 784 (1983)