XYLITOL DEHYDROGENASE O F BACILLUS PUMILUS
Kei YAMANAKA
Polyol dehydrogenases, D-sorbitol and D-mannitol dehydrogenases from microbial origins have been reported to be inducible with D-sorbitol and D-mannitol(1). Pentitol dehydrogenases were also produced with specific inducers, respective pentitols(2). Among four pentitol de- hydrogenases, D-arabitol and ribitol dehydrogenases were extensively studied on Klebsiella (Aerobacter) aerogener (3-8). L-Arabitol dehydrogenase was found, but few papers were available on the production and properties of this enzyme in Klebsiella aerogenes and Penzcillzum chryro- genum(gsl0). Diversity of xylitol dehydrogenase is of interest and occurrence of three xylitol dehydrogenases has been reported and identified as D-xylulose forming and L-xylulose forming enzymes. One is NAD-dependent and two are NADP-dependent enzymes Xylitol: NAD 2~oxidoreductase (D-xylulose forming, EC 1.1.1.9) is a common xylitol dehydrogenase and is called as D-xylulose reductase The second NADP-dependent enzyme is xylitol: NADP 1- oxidoreductasc (D-xylose forming, EC 1.1.1.139) which is called as xylose reductase The third NADP-dependent enzyme is Xylitol: NADP 4-oxidoreductase (L-xylulose forming, EC 1.1 1.10) and is also called as L-xylulose reductase D-Xylulose and L-xylulose forming two xylitol de- hydrogenases catalyze the transformation of D-xylulose to L-xylulose via xylitol in the glucuronate cycle of liver. Comparative studies on the specificities and reaction mechanisms of three xylitol dehydrogenases together with their location is of great interest for elucidation on the metabolism of xylitol, xylose and xyluloses and the significance of xylitol in the convergent reactions
From microbial origins, NAD-specific xylitol dehydrogenase (D-xylulose forming) was demon- strated in Klebrzella aerogener (2.9) The enzyme was produced only with xylitol as a n inducer. Ribitol dehydrogenase was also simultaneously induced with xylitol and properties of xylitol dehydrogenase were always similar to ribitol dehydrogenase(2). However, xylitol dehydro- genase was not demonstrated after growth on ribitol, even ribitol dehydrogenase was present most abundant in the same cells(2).
This brief note describes the demonstration of production of xylitol dehydrogenase (xylitol: NAD 2-oxidoreductase, D - X ~ ~ U ~ O S ~ forming) from D-sorbitol grown cells of Baczllus pumilur.
Materials and Methods Culture
Cells of Baczllur pumzlur, I F 0 3813 were grown aerobically by reciprocal shaking a t 30°C Each 500 ml flask for shake culture contained 100 ml of medium The cultural medium was composed of (glliter) K2HP04 7, K H 2 P 0 4 3, (NH4)2S04 2, yeast extract 1, MgS04 7 H 2 0 0.1, FeS04 0 01 and D-sorbitol 20. Sorbitol and magnesium sulfate were autoclaved separately. Inoculum was made in 5 ml of medium in a test tube and incubated by shaking overnight. Inoculum (0.1 ml) was transferred to 100 ml of the medium and incubated for 21 hours.
Activity measurements
Activity of' xylitol dehydrogenase was assayed in the following reaction system: 0.1 M phos- phate buffer (pH 7.0) 0.05 ml, 0.1 M D-xylulose 0..02 ml, 0..01 M NADH 3 p1 in total volume of 0.20 ml. For the D-sorbitol dehydrogenase assay, 0.01 ml of 2 M D-fructose was replaced to D-
234 Tech. Bull. Fac Agr Kagawa Univ..
xylulose
For the dehydrogcnation of xylitol, D-sorbitol or ribitol, 0 05 M Tris-HC1 buffer (pH 9 0) 0 10 ml, 0 5 M xylitol, D-sorbitol or ribitol 0 04 ml and 0 01 M NAD 0 0 15 ml in total volume of 0 20 ml Activities assays were conducted with a Hitachi model 139 spectrophotometer a t 340 nm equipped with microcuvette chamber and thermospaced a t 30'6 Microcuvette was purchased from Pyrocell Manufacturing Co
,
New Jersey, U S AActivity was calculated from the dehydrogenation of D - X ~ ~ U ~ O S ~ and expressed as xylitol de-
hydrogenase. A unit of dehydrogenase activity was defined as the amount oi enzyme required to produce a change in absorbance a t 340 nm of 1 0 per minute with D - X Y ~ U ~ O S ~ and NADH.
Chemicals
Xylitol, D- and L-arabitols and ribitol were purchased from General Biochemicals Inc., Ohio and NAD and NADH were obtained from Sigma Chemical Co
,
Missouri and P. L. Biochemicals Inc., Wisconsin, respectively D-Xyl~lose and D- and L-ribuloses were prepared as described previously (1) Purified preparations were free from aldopentoses by paper chromatography.Results Production of the enzyme
B. pumilus was used as a producer of xylitol dehydrogenase from D-sorbitol medium. The
production of enzyme was strongly enhanced with addition of 0 1% yeast extract Peptone showed neither concerted effect for enzyme production with yeast extract medium nor stimulat- ing effect with mineral medium D-Sorbitol was effective carbon source for production of the enzyme Optimum concentration of D-sorbitol was 2 0% and maximum lebel of the enzyme activity was reached after 20-22 hours' culture by shaking The bacterium was able to grow on sucrose or mannitol as a carbon source, but failed to produce the dehydrogenase.
Purification
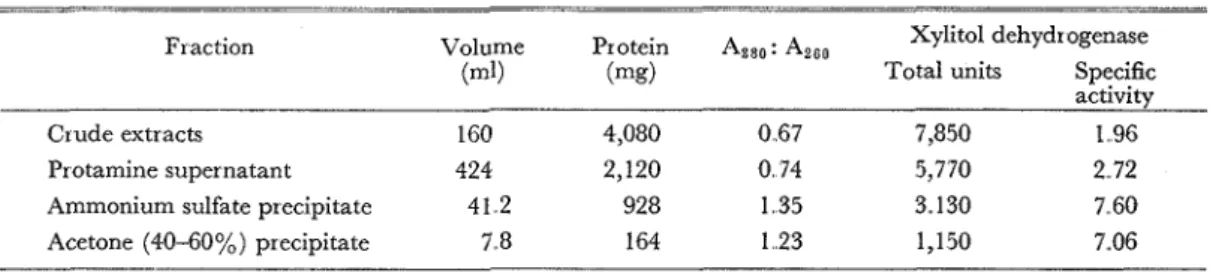
Cells were harvested by centrifugation from 30 flasks (3 1 of medium) after 21 hours incubation a t 30°C Cells (wet weight 25 g) were washed once with 0 05 M Tris-HC1 buffer (pH 7.0) containing 1 0 - 3 ~ mercaptoethanol and centrifuged The washed cells, suspended in lOOml of the same buffer, were divided two portions. Each portion was subjected to the sonic oscillator for 10 minutes. Crude extracts were obtained by centrifugation (108 ml) and residues were resuspendcd for repeated sonic treatment Second extracts (52 ml) was combined to the first extracts which was then diluted to 400 nll with the same buffer Freshly prepared 2% pro- tamine sulfate solution (40 ml) was added dropwise to the extracts Precipitate was centrifuged off Solid ammonium sulfate (94.5 g) was added to the supernatant (424 ml) to give 40% of saturation. Trace of precipitate was removed by centrifugation Addition of more am- monium sulfate (55 5 g) gave 60% of saturation. Precipitate was collected by centrifugation. Further addition of ammonium sulfate (87,7 g) yielded the second precipitate. Activity was recovered in two precipitates Both precipitates were dissolved in buffer, combincd and dialyzed against the same buffer for overnight (Ammonium sulfate precipitate) Fractionation the en- zyme was conducted by adding acetone, previously chilled a t -15"C, and precipitates from 0-20, 20-40 and 40-60% were collected by short time centrifugation a t -10°C. One third of activity was recovered in the precipitate of 20-40%, but remaining activity was found in the third precipitate. This preparation was used throughout this experiment Purification is summa-
Vol. 27, No.. 59 (1976) 235 Table 1
..
Purification of Xylitol Dehydrogenase--
Fraction Volume Protein Azso: AZe0 Xylitol dehydrogenase
(ml) (mg) Total units Specific
activity
Crude extracts 160 4,080 0.67 7,850 1.96
Protamine supernatant 424 2,120 0 74 5,770 2 72
Ammonium sulfate precipitate 41.2 928 1.35 3.. 130 7.60
Acetone (40-60%) precipitate 7.8 164 1.23 1,150 7..06
rized in Table 1.
Properties
Effect of pH. Effects of pH for the dehydrogenation of three sugar alcohols, xylitol, D-sorbitol and ribitol were found to be identical and maximal activities were obtained at pH 11.0. For reduction of ketose, activities were maximum with the pH range of 6.6 to 8.2 with D-xylulose, D-fructose and D-ribulose and NADH.
Substrate specificity. As polyol dehydrogenases are regarded to show affinity on several hexitols and pentitols, substrate specificity of the enzyme preparation was tested with hexitols, pentitols and ketoses with NAD or NADH.. As summarized in Table 2, the enzyme exhibited
Table 2. Substrate Specificity
Substrate Activity*' Relative Substrate Acti~ity."~ Relative
(As.ro/min) ratio (%) (Astolmln) ratio (%)
Xylitol 0 194 100 D-Xylulose 0.248 100
D-Sorbit01 0 161 83 D-Fructose 0 260 105
Ribitol 0 055 28 D-Ribulose 0 212 86
D-Mannitol 0 0008 0 4
L-Arabitol 0 0026 1.3 L-Ribulose trace 0 5
D-Ar abitol 0 0
z-Er ythritol 0 0
*'
Activity was measured with 0, l~ substrate at p H 9.0.h 2 Activity was measured with 0 . 0 . 5 ~ substrate at p H 7.1..
the highly affinity on xylitol and D-sorbitol at pH 9.0 D-Ribitol was also received the dehydro- genation with relatively slow rate. L-Arabitol and D-mannitol were innert for the enzyme For the reduction of ketose, enzyme showed equal affinity on D-xylulose and D-fructose. D-Ribulose was also reduced with NADH. L-Ribulose was innert ibr the enzyme. The Michaelis constants for these substrates were calculated from the Lineweaver-Burk's plots shown in Figs. 1 and
2:
1.47, 2.38 and 10 mM for xylitol, D-sorbitol and ribitol a t pH 11, and 4.5, 71 and 15 mM for D-xylulose, D-fructose and D-ribulose at pH 7.0, respectively. These characters which is most specific for oxidation-reduction reaction between xylitol and D-xylulose supported to name the enzyme as xylitol dehydrogenase, even the enzyme was induced in the D-sorbitol mediumCoenzyme specificity. The enzyme required specifically NADH for the reduction of D-
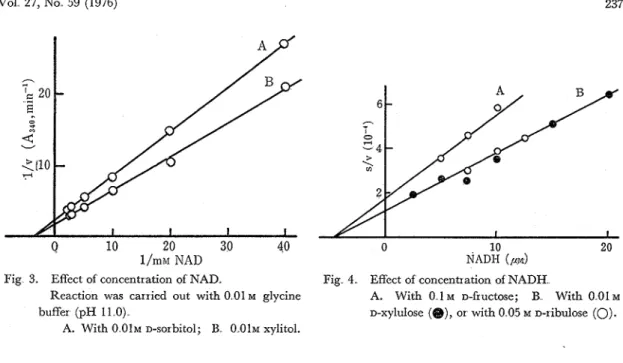
xylulose and D-fructose. These activities reduced to 4 6-6% when NADH was replaced to NADPH as shown in Table 3. The Michaelis constants for NAD and NADH were calculated as 0 28 and 0.047 mM, respectively (Figs. 3 and 4).
Vol. 2'7, No.. 59 (1976)
l / m ~ NAD
Fig 3. Effect of concentration of NAD. Fig 4 Effect of concentration of NADH
Reaction was carried out with 0 01 M glycine A. With 0 1 M D-fructose; B With 0 01 M
buffer (pH 11 0) D-xylulose (@), or with 0 05 M D-ribulose
(0).
A. With 0 0 1 ~ D-sorbitol; 13 0 0 1 ~ xylitol.
Discussion
Several evidences suggested the identity of L-iditol dehydrogenase and xylitol dehydrogenase with NAD-specific L-iditol (D-sorbitol) dehydrogenase from sheep liver (crystalline) (11) and
Azotobacter agilis(l2). Substrate specificities of two dehydrogenases were compared in Table 4.
Table 4. Substrate Specificity of L-Iditol, D-Sorbitol and Xylitol Dehydrogenases
Dehydrogenase Relative activity ratio
(Origin) D-Sorbitol L-Iditol Xylitol Ribitol D-Mannitol D-Arabitol L-Arabitol
L-Iditol dehydrogenase 100
-
90 79-
- 23 (sheep liver) (") L-Iditol dehydrogenase 100 102 121 9-
- (Azotobacter agilir) ( I 2 ) Polyol dehydrogenase 87 4 1 100 23 23 7 1 (Candzda utilis) ( I 3 ) Polyol dehydrogenase 72 - 100 50 4 2 5 (Candida albicans) (") B. pumilur*
83 - 100 28 0 4 0 1 3*
Table 2 in this paperCH20H R : CH20H xylitol
-
H-C-OH D-sorbitol-C
CHPOH I HO-C-H L-iditol I CH20HDifference may be found only on the relative activity ratio on four sugar alcohols B pumilus enzyme in this paper was closely resembled to L-iditol dehydrogenase but more likely to polyol
238 Tech. Bull.. Fac. Agr. Kagawa Univ.
dehydrogenase (xylitol dehydrogenase) from Candzda utzlzr(13) or C. albicans(l4).
Dissimilation of sorbitol in K aerogenes, PRL-RS involved a PEP-dependent sorbitol kinase (PEP: sorbitol 6-phosphotransferase) and sorbitol 6-phosphate dehydrogenase system. The possibility on the direct oxidation of sorbitol to D-fructose was conclusivcly ruled out(l5). More- over, activity on the oxidation of sorbitol by ribitol dehydrogenase from K. aerogenes was not demonstrated in papers. It can be concluded that specific sorbitol dehydrogenase does not presented in this bacterium.
Xylitol dehydrogenase (NAD-specific) was concomitantly induced with ribitol dehydrogenase in K aerogene~(2.9) Wild type of K. aerogener, 1033 was unable to utilize xylitol. But after the ability to utilize xylitol was achieved upon successive transfer, a mutant was able to produce constitutive ribitol dehydrogenase(8). Xylitol was a substrate for ribitol dehydrogenase but not a n inducer. Purified ribitol dehydrogenase from mutant strain of K. aerogenes showed affinity toward ribitol and xylitol The K m value for xylitol was high as 0.12-0.29 M compared with that for ribitol as low as 2 0-3 1 m ~ ( 8 ) . I n consequence, ribitol dehydrogenase shows affinity to xylitol, but the enzyme is different from xylitol dehydrogenase Another evidence for two differ- ent enzymes is the difference of the Michaelis values for pentitols. L-Iditol dehydrogenase from sheep livercll), polyol dehydrogenase from Candzda utilis(l3) and B pumilus enzyme in this paper showed high affinity to xylitol and D-sorbitol than ribitol. K m values for xylitol, D-sorbitol and ribitol were expressed as mM as follows: 11, 32 and 70 (Candzda utzlis(l3)), 0.18, 1.1 and 1.8 (sheep liver(l1)) and 1 47, 2.38 and 10 (B. pumzlus).
I n conclusion, B pumzlus enzyme in this paper was close to L-iditol dehydrogenase or xylitol dehydrogenase. I t seems to be more preferable to call this enzyme as xylitol dehydrogenase or D-xylulose reductase
This enzyme can be used for the enzymatic assay of D-xylose isomerase activity as the coupling enzyme and may be useful for the enzymatic preparation of D-xylulose from xylitol.
Summary
Xylitol dehydrogenase (Xylitol: NAD 2-oxidoreductase, EC 1.1.1.9) was demonstrated in the D-sorbitol grown cells of Baczllus pumzlur. Several properties were reported for the enzyme partial- ly purified by protamine treatment, ammonium sulfate and successive acetone fractionations The enzyme exhibited three dehydrogenase activities: xylitol2~-xylulose, D-sorbitol2~-fructose and ribitol2~-ribulose. Enzyme was NAD-dependent for these substrates and NADPH showed 4 6-6% of activity with NADH Optimum pH were 11 0 for the dehydrogenation of polyols (NAD reduction) and broad pH range a t 6 6-8 2 for the reduction of ketoses (NADH oxidation) The Michaelis constants are: 4 5 mM for D-xylulose, 71 mM for D-fructose, 15 mM for D-ribulose,
1.47 mM for xylitol, 2.38 mM for D-sorbitol, 10 mM for ribitol, 280 PM for NAD and 47 ,UM for NADH
Literatures
( 1 ) YAMANAKA, K., SAKAI, S. : Catz.. J . Microbiol.., ( 3 ) LIN,E..C.C.: J. Biol. Chem.,236, 31 (1961).
14, 391 (1968). ( 4 ) WOOD, W. A , , MCDONOUYH, M. J., JACOBS,
( 2 ) MORTLOCK, R. P., Foss~rr, D. D., PEIERING, L. B.: , J,. Bwl.. Chem.., 236, 2190 (1961)..
D.. H., WOOD, W.. A. : , J. Bacterial., 89, 129 ( 5 ) FROMM, H. J..: , J. Biol. Chem., 233, 1049 (1958)..
Vol.. 27, No. 59 (1976) 239 C.: Bzochzm Bzophys Acta, 67, 219 (1963)
( 7 ) BISSON, T M , MORILOCK, R P.: J Bac-
terzol
,
95, 925 (1968)( 8 ) Wu, T T , LIN, E C. C , TANAICA, S : J
Bacterzol., 96, 447 (1968).
( 9 ) FOSSII I, D D , MORTLOCK, R P , AND~RSON,
R L , WOOD, W A : J Bzol Chem , 239,
21 10 (1964)
(10) CHIANG, C., KNIGHT, S. G : Bzochzm Bzophys
Acta, 46, 271 (1961)
(1 1) SMITH, M G : Biochem J., 83, 135 (1962).
(12) MARCUS, L , MARR, A G. : J. Bacterzol., 82,
224 (1961)
(13) CHAKRAVORIY, M , VEIGA, L A., BACILA, M
,
HORECKER, B L : J Bzol Chem , 237, 1014
(1962).
(14) VEIGA, L A : J Gen Abpl Mzcrobzol, 14,
79 (1968)
(1 5) KELKER, N E , ANDERSON, R L : J. Bac-
terzol., 105, 160 (1971)
B..pumilus O D-YILC-Y 1.&.?&%i#i=f~K.+VY I - - / L : NAD f i ; l k R @ ~ ~ @ Q R / l i L , < . & Q @ f i @ g L , %
O B E Q ~ . ~ ~ : . . * ~ ~ i - d r , 4 . 9 9 I--/LOC, Y I L ~ . I., 11
r
I . - - , L ~ C % ~ ~ H L, %oj;f;&.~~-dr+~/ 9 ~ - . I V Q1002 L k % , lkL?-'a283, 28'T&->?z.. pH G-dr@'7/~=.-~/~ O@{kEETCb pH 11,,0, @ 1.
,-%o
D-+ ~ I La ' - % , D - 7 4 9 I.-%, D - ~ ' ~ * D - % ~ % ~ ~ E T ~ ' ~ : pH 6.6-8.2 .T&..r,k. *@$%Otik6%%tCi(f?& < I \
.X 9 %%%l-dr, 1 . 4 7 m ~ (+ 9 9 1. .-/L) , 2 . 3 8 m M ( D - Y / ~ c * ~ I.), lOmM ( 9 c I..-/L), 4.5mM ( D - + ~ / L P , - % ) ,
7 1 m ~ ( D - 7 7 4 I-.-%), 1 5 m ~ (0-9 Y Q - x ) ,T&,=f:. G k ' % % t L ' r NAD Q % % L , NADPH 1% NADH
046-6%O%&L.d~5S6j51.9f:.. COKmlalb280p~(NAD),47p~(NADH) , T i % ) , 2&g9+99 I..-IL,
D - Y I L ~ Y I . ~ ~ : @ D - = ~ ~ I L P , - X 2 D-7 4 P l.-%~Hh7:$2bf:%%%O Km C & & ( H--.E&..>f:.
U k . O % % , *dl.Cd:)&.&SkL&B?%dt, + 9 9 I.,-/1.8#7k%@$% ( D - ~ . ~ I L D . - % 1/&*?4,-,-E, EC1..1..
i , , g ) * ~ % o ~ I J ~ ~ c ~ ~ ~ ~ ~ - ~ I . - s R L ~ : , ,