北海道医療大学学術リポジトリ
Porphyromonas gingivalis由来LPS長期刺激による ヒト歯根膜線維芽細胞におけるDNA高メチル化の網 羅的解析‐細胞外マトリックス関連および老化抑制 関連の遺伝子について‐
著者 ?井 理衣
学位名 博士(歯学)
学位授与機関 北海道医療大学
学位授与年度 平成27年度 学位授与番号 30110甲第272号
URL http://id.nii.ac.jp/1145/00010477/
Porphyromonas gingivalis 由来 LPS 長期刺激による
ヒト歯根膜線維芽細胞における DNA 高メチル化の網羅的解析
‐細胞外マトリックス関連および老化抑制関連の遺伝子について‐
平成 27 年度
北海道医療大学大学院歯学研究科
髙 井 理 衣
要 旨
歯周ポケットは様々な外的因子に曝露されており,Porphyromonas gingivalis (P.
gingivalis) な ど の グ ラ ム 陰 性 細 菌 も 多 く 存 在 す る . ま た , そ れ ら の 菌 が 有 す る Lipopolysaccharide(LPS)により,歯周炎が惹起され歯周組織が破壊されることはよ く知られている.近年,歯周疾患の進行や治癒にエピジェネティクスが関与するとい う報告がなされているが,未だ解明されていない部分が多い.エピジェネティクスと は,DNA 塩基配列の変異を伴わずに遺伝子の化学的修飾によって発現が変化する現 象であり,細胞分化,癌,先天的疾患や薬剤耐性など多岐にわたる生命現象に関わる とされている.代表的な修飾に,DNA メチル化やヒストン修飾などがある.歯科界 でもエピジェネティクス研究が進められているが,扁平上皮癌など腫瘍性疾患に関連 したものが多く,歯周疾患について報告は少ない.さらに,DNA メチル化は長期的 な刺激により生じることが多いのに対し,歯周病原性細菌や LPS を使用した研究では 短期間での実験系が多いためメチル化を観察した報告は少ない.そこで本研究では,
歯周疾患とエピジェネティクスの関係を解明すべく,歯周病原性細菌である P.
gingivalis 由来 LPS 刺激下での長期細胞培養法を確立するとともに,LPS 長期刺激に
よるヒト歯根膜線維芽細胞(HPDLFs)における DNA 高メチル化と遺伝子発現の変化 および脱メチル化処理による影響について検討を行った.
本研究では,細胞に対し LPS を 3 日毎間欠的に刺激することによって,長期にわた り継続的に刺激を行う新たな実験モデルを確立した.この実験系では,多くの遺伝子 に DNA メチル化状態の変化がみられた.特に,細胞外マトリックスに関連するもの では 9 遺伝子(FANK1, COL4A1-A2, COL12A1, COL15A1, LAMA5, LAMB1, MMP25,
POMT1, EMILIN3) ,老化抑制関連では 1 遺伝子(Klotho)で DNA 高メチル化を引き
起こすとともに, mRNA やタンパク質の有意な発現低下が認められた.また,LPS 長 期刺激によって起こった DNA 高メチル化を脱メチル化剤 5-Aza-deoxycytidine で処理 すると,遺伝子発現の低下に改善が確認された.以上のことから,歯周病原性細菌 P.
gingivalis 由来 LPS 長期刺激により, HPDLFs の細胞外マトリックスや老化抑制に関連
する遺伝子に DNA 高メチル化が起こり,遺伝子発現を抑制することが明らかとなっ
た.
目 次
Ⅰ.緒言 ……… 1
Ⅱ.材料および方法 ……… 3 1.細胞毒性試験
2.LPS 刺激下での長期細胞培養
3.メチレーションアレイ解析
4.mRNA 発現解析
5.DNA メチル化解析
6.タンパク質発現解析 7.脱メチル化解析 8.統計分析
Ⅲ.結 果 ……… 6 1.LPS の HPDLFs への影響
2.LPS 長期刺激による HPDLFs の DNA メチル化の網羅的解析
3.LPS 長期刺激による HPDLFs の mRNA 発現解析
4.LPS 長期刺激による HPDLFs の DNA メチル化レベル解析
5.LPS 長期刺激による HPDLFs のタンパク質発現解析
6.脱メチル化処理による遺伝子発現の変化
Ⅳ.考 察 ……… 8
1. LPS 長期刺激による培養系の確立について
2.DNA メチル化状態の変化について
3.高メチル化に伴う mRNA の発現の低下がみられた細胞外マトリックス関連遺
伝子について
4.高メチル化に伴い mRNA の発現低下がみられた老化抑制関連遺伝子について
5.LPS 長期刺激による HPDLFs のタンパク質の発現変化について
6.脱メチル化処理による mRNA の発現回復について
7.本研究の臨床的意義
Ⅴ.結 論 ……… 13
Ⅵ.謝 辞 ……… 14
Ⅶ.文 献 ……… 15
表・付図 ……… 20
1
Ⅰ.緒 言
歯周炎は,歯周組織内で細菌感染に伴う免疫応答や歯周病原性細菌のもつ内毒素,線 毛,ジンジパイン,コラゲナーゼ,プロテアーゼ,ロイコトキシンなどにより,炎症 性サイトカインの産生,コラーゲンやフィブロネクチンなどの細胞外マトリックスの 破壊・減少による歯槽骨の吸収や結合組織の破壊が起こり(Potempa J et al., 2000) , 最終的には歯の喪失を招いてしまう慢性炎症性疾患である.近年の疫学調査において,
高齢になるほど歯周疾患罹患率は上昇しており,また全身への影響も明らかになって きていることから(Cullinan MP & Seymour GJ, 2013) ,超高齢社会を迎えた日本にお いては,さらに重要な疾患と位置付けられてきている(宮下,2013).歯周炎の発症 や進行には,細菌感染を始めとする外的要因以外に,老化や全身疾患,ホルモン,遺 伝的な素因などの内的な要因も大きく関わっている(前田 & 濵地, 2013) .内的要因 の根幹にあるものに遺伝子の表現様式があるが,近年遺伝子の表現様式が外的要因に よって変化するエピジェネティクスが注目されてきている(Eggar G et al., 2004; Barros SP & Offenbacher S, 2014; Larsson L et al., 2015).
エピジェネティクスとは,DNA 塩基配列の変異を伴わず,遺伝子の化学的修飾に よって発現を変化させ,遺伝子機能を後天的に制御する現象であり,DNA メチル化 やヒストン修飾,クロマチン構造の形成,リモデリングなどが知られている(Eggar G
et al., 2004) .中でも DNA のメチル化は,一旦,変化が及ぼされると遺伝子機能に長
期的な影響を与える可能性があり,疾患の発症・進行に深く関わるとされている
(Eggar G et al., 2004).特に,プロモーター領域の高メチル化は,転写因子の結合を 阻害し,遺伝子発現を強く抑制することから,がん抑制遺伝子のプロモーター領域の 高メチル化と発がんとの関係について広く研究が行われてきた(Baylin SB & Jones PA, 2011; Dawson MA & Kouzarides T, 2012) .近年,この領域の研究が糖尿病,肥満,アレ ルギー,自己免疫疾患,精神神経疾患などのがん以外の疾患へと広がりをみせている
(Abiko Y et al., 2014).口腔領域でも,口腔がんや前癌病変の発症・進行への関与に ついて報告がなされてきており(Gasche JA & Goel A, 2012; Mascolo M et al., 2012) ,最 近になり歯周病にもエピジェネティクスが関与するという報告がみられるが(Larsson L et al., 2015; Barros SP & Offenbacher S, 2014; Ben-Avraham D, 2015),その詳細は未だ 不明な点が多い.すなわち, COX-2, IL-2, IL-6, IL-8, IL-10, TNF-αのメチル化異常など,
サイトカインをはじめとした炎症に関わる報告が多く,歯肉上皮,線維芽細胞,骨芽 細胞など歯周組織を構成する細胞の恒常性に関わるものは僅かである(Larsson L et al., 2015) .
歯周炎 の際に 形成 される歯 周ポケ ット には,歯 周病原 性細 菌 Porphyromonas
gingivalis (P. gingivalis) を代表とする多くのグラム陰性菌が存在している.それらの細
胞壁外膜には, Lipopolysaccharide (LPS) が存在し,生体に対して多様な生物活性を発
現させ,歯周炎の病態を形成・進行させる一因になっているといわれている(Ding PH
2
& Jin LJ, 2014) . 歯周炎の in vitro におけるモデル系として, 歯周組織の細胞を培養し,
LPS による刺激で起こる細胞内の変化を観察した報告は比較的多い(Doyle CJ et al., 2015; Jones KJ et al., 2010; Nanbara H et al., 2012; Wen WC et al., 2014) .そのほとんどが 数時間や 2, 3 日程度の刺激を行ったものであり,慢性的な刺激による歯周炎の発症を 想定したモデルとは言い難く,DNA のメチル化に著明な変化を及ぼすことは少ない ものと考えられる.
そこで本研究では,歯周疾患の発症と進行に関わる歯根膜細胞における DNA のメチ ル化変化を明らかにするため,最初に,ヒト歯根膜由来線維芽細胞 (HPDLFs) を用い て,P. gingivalis 由来 LPS による長期刺激を行う新たな実験モデルの確立を試みた.
次に,この実験モデルを用いて,DNA メチル化の変化を網羅的に観察するために,
メチル化アレイによる検討を行った.このデータの中から,発現が低下することによ って歯周組織の恒常性の維持に悪影響を与えると考えられる細胞外マトリックスと,
老化抑制に関連する遺伝子を抽出し,それらの高メチル化と遺伝子発現の変化および
脱メチル化処理による影響について検討を行った.
3
Ⅱ.材料および方法 1.細胞毒性試験
ヒト歯根膜線維芽細胞 HPDLFs (LONZA 社)を 35 mm tissue culture plates (IWAKI 社)に 3.0×10
4cells/mL 播種し,10% Fetal bovine serum (FBS, Sigma 社)含有 Dulbecco's Modified Eagle Medium (DMEM, Sigma 社)にて 24 時間培養後,P. gingivalis 由来 LPS (WAKO 社, ATCC 33277) を濃度 0.1, 1, 10, 100 µg/ml でそれぞれ添加し,72 時間およ び 144 時間培養を行った.その後,細胞を 0.05%トリプシン溶液 (gibco 社)処理にて 回収し,0.5 % Trypan Blue 溶液にて細胞を染色,血球計算盤を用いて細胞数を計測し た.
2.LPS 刺激下での長期細胞培養
HPDLFs を 10% FBS 含有 DMEM にて 24 時間培養後, DMEM に P. gingivalis 由来 LPS
を濃度 1 µg/mL 添加したものと非添加したもので 3 日間ずつ交互に交換し,1 ヶ月間
培養し,細胞形態の観察および DNA メチル化解析のサンプルとした.また,コント ロールには LPS の代わりに滅菌水を添加し,同期間培養したものを用いた(図 1) . 3.メチレーションアレイ解析
培養した細胞から Qiagen
®DNeasy Blood & Tissue Kit (Qiagen 社)を用いて DNA を抽出 した.その後 DNA を超音波処理により断片化,精製した後, cytidine 5-dUTP (Cy5) お よび cytidine 3-dUTP (Cy3) にて蛍光ラべリング,Human CpG islands 224k array に DNA をハイブリダイズ, DNA Microarray Scanner (Agilent technology 社) にて検出,解 析ソフト Gene Spring GX, GCH Explorer (Agilent technology 社)を用いて解析を行った.
得られたアレイ解析結果から,細胞外マトリックスもしくは老化をキーワードにそれ ぞれに関連する遺伝子を検索し, その中でさらに遺伝子がプロモーター領域に位置し,
かつメチル化レベルが 4 倍以上の高値を示すものを選出した.
4.mRNA 発現解析
培養細胞より TRIzol
®(Invitrogen 社)にて total RNA を抽出した後,濃度を 2 µg/µl にな るよう調整し,Super Script
®reverse transcriptase (Invitrogen 社) と oligo(dT)
12-18primer
(Invitrogen 社) を用いて逆転写を行った.また,プライマー設計ソフト Primer3
(http://bioinfo.ut.ee/primer3-0.4.0/) を用いて各遺伝子に対応するプライマーを設計した
(表 1).得られた cDNA とプライマー,SYBR
®Green PCR Master Mix (Applied
Biosystems 社) を用いて ABI 7500 Real time PCR System (Applied Biosystems 社) にて定
量的 real-time PCR を行った. PCR 条件は, 7500 System SDS Software Version 1.3 (Applied
Biosystems 社) にて 50℃ 2 分間,95℃ 10 分間で反応させ,95℃ 15 秒間,60℃ 1 分
間を 40 サイクルで行った.ターゲット遺伝子の発現レベルは ,Glyceraldehyde
4
3-phosphate dehydrogenase (GAPDH) をリファレンス遺伝子として⊿⊿CT 法(Livak KJ
& Schmittgen TD, 2001)を用いて比較した.
5.DNA メチル化解析
DNA メチル化の変化の再現性を確認するため,培養した細胞から Qiagen
®DNeasy Blood & Tissue Kit (Qiagen 社)を用いて DNA を抽出した.DNA 濃度を 2 µg/µl になる よう調整し,Qiagen
®Epitect Bisulfite Kit (Qiagen 社)を用いて Bisulfite 処理を行った.
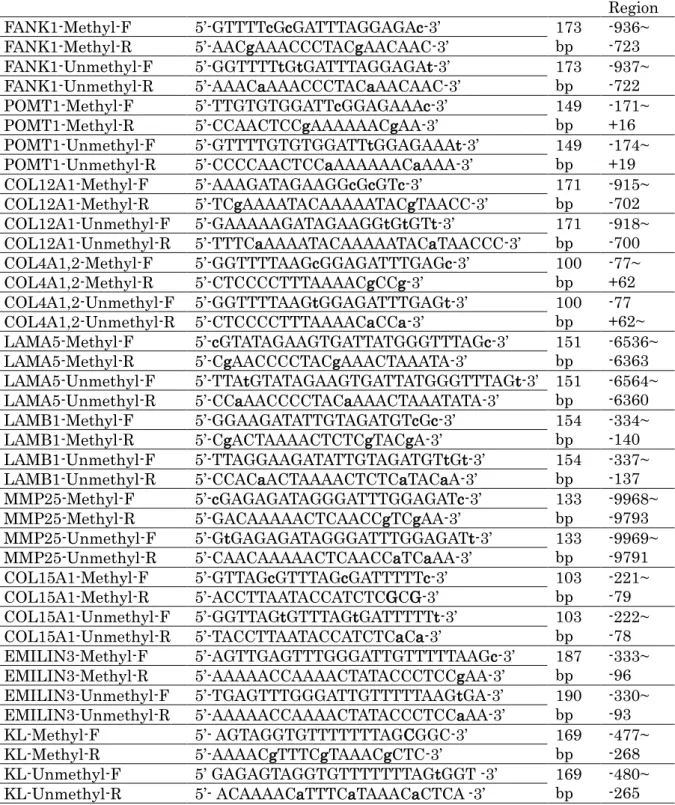
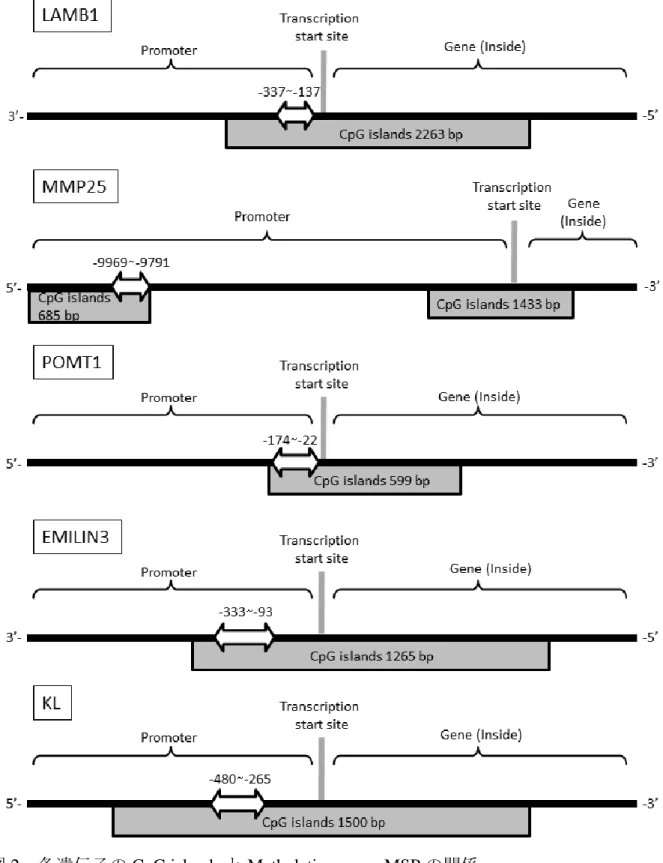
また,2.メチレーションアレイ解析によって得られた結果から検索した細胞外マト リックスおよび老化に関連する遺伝子のプロモーター領域で高メチル化のみられた CpG 配列部位に焦点を絞り,Bisulfite 処理後の DNA に対応した Methylation-Specific PCR (MSP) プライマーを設計ソフト Methyl Primer Express
®(Applied Biosystems 社) に て設計した(表 2,図 2) .その後, Bisulfite 処理 DNA, MSP プライマー, SYBR
®Green PCR Master Mix を用いて ABI 7500 Real time PCR System にて MSP 法によるメチル化 レベル解析を行った.PCR 条件は,7500 System SDS Software Version 1.3 にて 50℃ 2 分間,95℃ 10 分間で反応させ,95℃ 15 秒間,60℃ 1 分間を 50 サイクルで行った.
メチル化レベルの算出には,以下の計算式を用いた(Lu L et al., 2007) .
メチル化レベル
(%) =メチル化レベル
Qty.メチル化レベル
Qty. +非メチル化レベル
Qty.× 1006.タンパク質発現解析
タンパク質の発現は,Cellular Enzyme-Linked ImmunoSorbent Assay (CELISA)法にて解 析した.LPS 刺激を加えた培養細胞を 0.05%トリプシン溶液処理にて回収後,96 well plate (IWAKI 社)に 3×10
4cells/ml 再度播種し, 24 時間培養した. その後, In Cell ELISA Kit (Cosmo Bio 社)と一次抗体として Mouse anti-GAPDH Antibody (In Cell ELISA Kit), Fibronectin type III and ankyrin repeat domains 1 (FANK1) Antibody (E-5,SantaCruz 社), Collagen type XII alpha 1 (COL12A1) Antibody (A-11,SantaCruze 社), Anti-Human Klotho Antibody (PEPROTECH 者)を,二次抗体には HRP-Conjugated Anti-Mouse IgG Antibody (In Cell ELISA Kit)を用いて処理を行い, Bio-Rad
®Model 680 Microplate Reader (Bio Rad
社)にて 450 nm の吸光度を測定,付属の GAPDH 抗体の値にて補正し,タンパク質発
現量を解析した.
7.脱メチル化解析
LPS 刺激を加えた細胞群に,100 µM 濃度の 5-Aza-deoxycytidine (5-Aza, Sigma 社)を加
え 24 時間処理を行った(Uehara O et al., 2014) .その後,TRIzol
®法にて total RNA を
抽出し,方法 4 に準じて脱メチル化処理前後の mRNA の発現を比較した.
5
8.統計分析
得られた結果はすべて,IBM SPSS Statistics 20 (IBM 社)を用いた Mann-Whitney U 検
定および Kruskal-Wallis 検定にて比較・検討し,p <0.05 を有意な変動と判断した.
6
Ⅲ.結 果 1.LPS の HPDLFs への影響
HPDLFs に対する P. gingivalis 由来 LPS の細胞毒性試験では,72 時間および 144 時間 後ともに, LPS 10, 100 µg/mL で細胞数の有意な減少がみられた(p<0.05) (図 3).よ って, LPS 長期刺激培養に使用する LPS 濃度は,有意差のみられなかった最大濃度 1 µg/mL に設定した.P. gingivalis 由来 LPS を HPDLFs に対し継続的に添加すると,1 ヶ月経過する前に細胞が死滅した.そのため,P. gingivalis 由来 LPS を 3 日毎間欠的 に添加したところ,細胞は培養皿上で終始,紡錘形を呈しており,1 か月を経過して もコントロール群と同様な形態であった(図 4) .
2.LPS 長期刺激による HPDLFs の DNA メチル化の網羅的解析
新たに確立した LPS 長期刺激による HPDLFs の DNA メチル化の網羅的解析では,
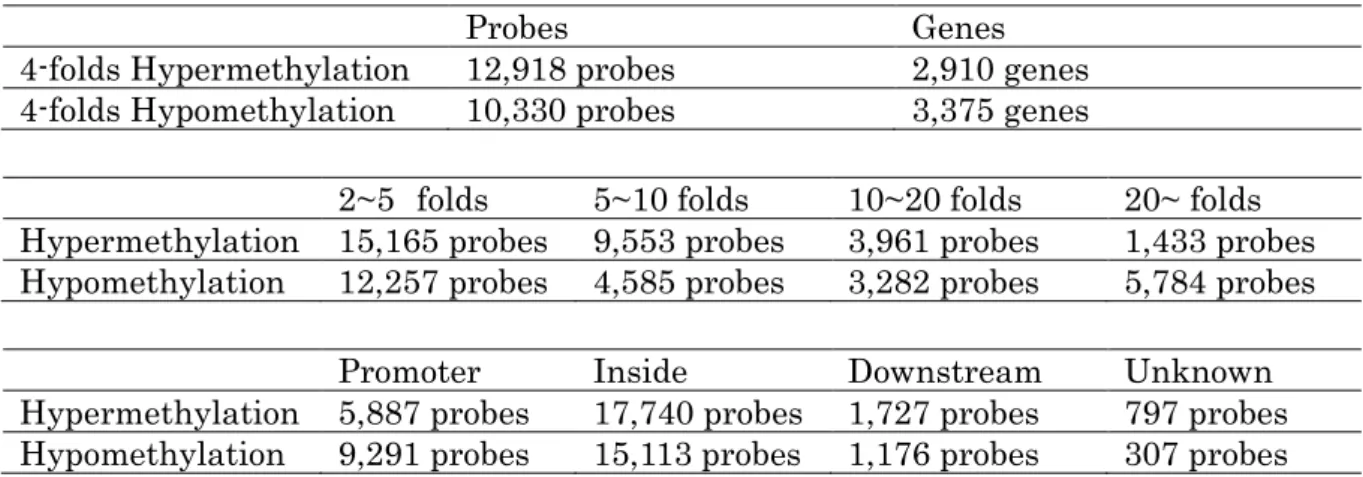
全遺伝子中,4 倍以上の高メチル化がみられたのは 12,918 プローブ(2,910 遺伝子) , 1/4 倍以下の低メチル化がみられたのは 10,330 プローブ(3,375 遺伝子)であった(表 3) .また,転写に関わるプロモーター領域では,高メチル化が 5,887 プローブ,低メ
チル化が 9,291 プローブ認められた(表 3) .さらに,プロモーター領域で 4 倍以上の
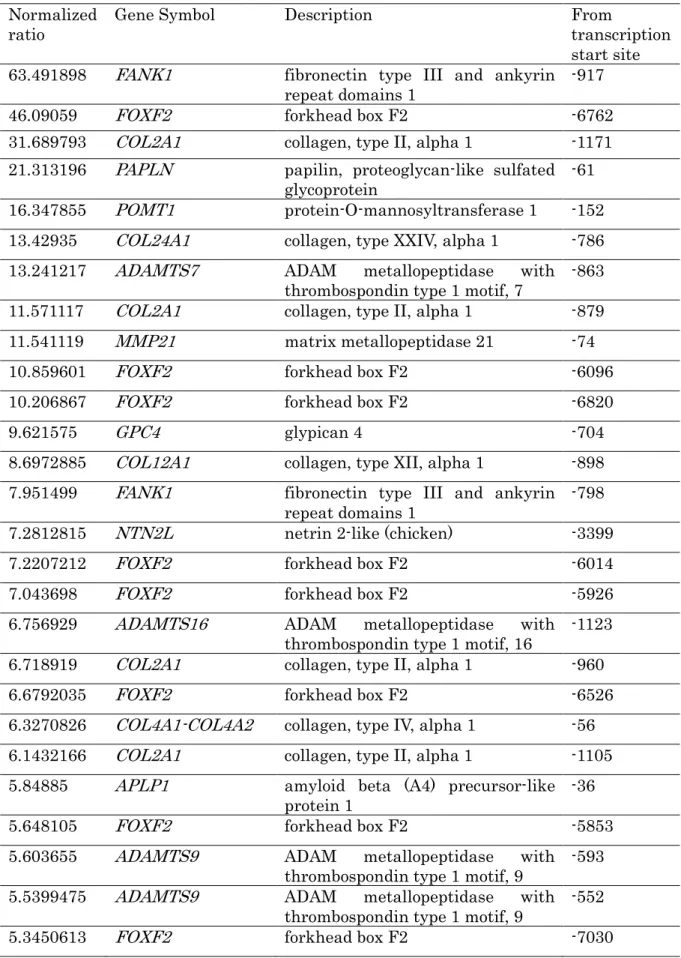
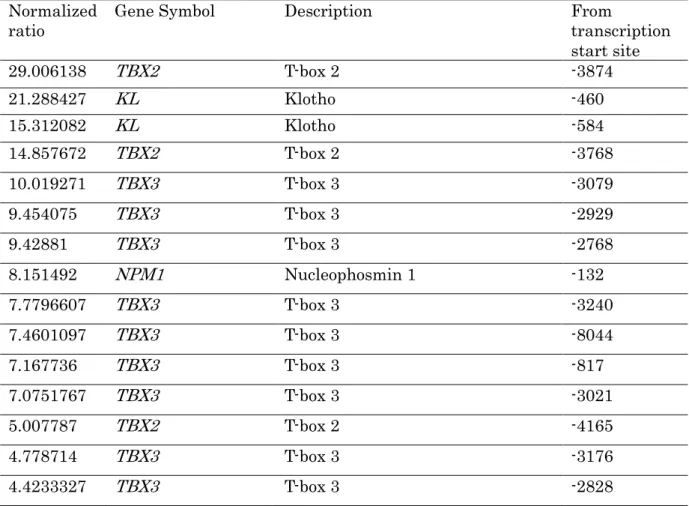
高メチル化がみられたものの中で細胞外マトリックスと老化抑制に関連する遺伝子 を検索すると,細胞外マトリックス関連では fibronectin type III and ankyrin repeat domains 1 (FANK1), forkhead box F2 (FOX2), collagen type II alpha 1 (COL2A1), papilin (PAPLN), protein-O-mannosyltransferase 1 (POMT1), collagen type XXIV alpha 1 (COL24A1), ADAM metallopeptidase with thrombospondin type 1 motif 7 (ADAMTS7), matrix metallopeptidase 21 (MMP21), glypican 4 (GPC4), collagen type XII alpha 1 (COL12A1), netrin 2-like (NTN2L), ADAM metallopeptidase with thrombospondin type 1 motif 16 (ADAMTS16), collagen type IV alpha 1 (COL4A1), amyloid beta A4 precursor-like protein 1 (APLP1), ADAM metallopeptidase with thrombospondin type 1 motif 9 (ADAMTS9), ADAM metallopeptidase with thrombospondin type 1 motif 5 (ADAMTS5), matrilin 4 (MATLN4), laminin alpha 5 (LAMA5), laminin beta 1 (LAMB1), chondroadherin (CHAD), collagen type VI alpha 1 (COL6A1), matrix metallopeptidase 25 (MMP25), collagen type XV alpha 1 (COL15A1), latent transforming growth factor beta binding protein 1 (LTBP1), elastin microfibril interfacer 3 (EMILIN3)の 25 遺伝子,老化抑制関連では T-box 2 (TBX2), Klotho (KL), T-box 3 (TBX3), Nucleophosmin 1 (NPM1)の 4 遺伝子が検 出された(表 4,図 2) .
3.LPS 長期刺激による HPDLFs の mRNA 発現解析
アレイ解析により高メチル化のみられた細胞外マトリックス関連および老化抑制関
連の遺伝子について, mRNA の発現解析を行ったところ,細胞外マトリックス関連で
7
は FANK1(25.15%), COL4A1-A2(50.30%), COL12A1(65.60%), COL15A1(36.97%), LAMA5(57.12%), LAMB1(45.34%), MMP25(35.21%), POMT1(54.41%),
EMILIN3(52.21%)の 9 遺伝子,老化抑制関連では KL(19.39%)のみ LPS 長期刺激によ
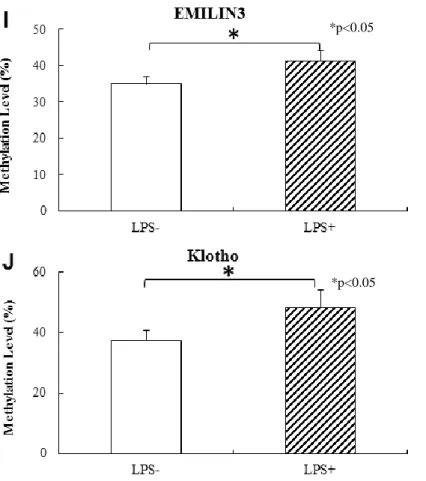
り有意な発現低下が認められた(p<0.05) (図 5) .
4.LPS 長期刺激による HPDLFs の DNA メチル化レベル解析
MSP 法において mRNA 発現に有意な低下にみられた細胞外マトリックス関連 9 遺伝 子 (FANK1, COL4A1-A2, COL12A1, COL15A1, LAMA5 LAMB1, MMP25, POMT1,
EMILIN3)と老化抑制関連 1 遺伝子(KL)の DNA 高メチル化の再現性を確認したところ,
すべての遺伝子で LPS 長期刺激によりメチル化レベルの有意な上昇が確認された(p
<0.05) (図 6) .
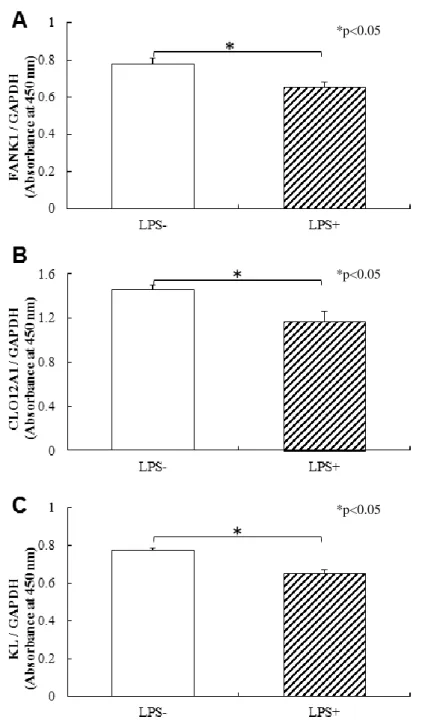
5.LPS 長期刺激による HPDLFs のタンパク質発現解析
mRNA の発現低下がみられたもので,FANK1, COL12A1, KL について CELISA 法に よるタンパク質の発現解析を行った.いずれも FANK1, COL12A1, KL ともに LPS 長 期刺激により有意な発現低下が認められた(p<0.05) (図 7) .
6.脱メチル化処理による遺伝子発現の変化
5-Aza により脱メチル化処理を加えたところ,細胞外マトリックス関連 9 遺伝子
(FANK1, COL4A1-A2, COL12A1, COL15A1, LAMA5 LAMB1, MMP25, POMT1,
EMILIN3)と老化抑制関連 1 遺伝子(KL)では,LPS 長期刺激群と比較して mRNA 発現
に有意な回復が認められた(p<0.05) (図 8) .しかし,コントロール群と 5-Aza 添加
群を比較すると FANK1, EMILIN3, KL では有意に発現が低下しており,完全な回復ま
でには至らなかった.
8
Ⅳ.考 察
1.LPS 長期刺激による培養系の確立について
本研究では, LPS を濃度 1 µg/mL 添加したものと非添加したもので 3 日間ずつ交互に 交換し,1 ヶ月間培養し,線維芽細胞に毒性変化を伴わずに DNA メチル化を観察す る新たな培養系を確立した.DNA メチル化は,細菌やホルモンなどの慢性刺激によ り亢進するといわれているため,DNA メチル化の影響の少ないと考えられる若い男 性のヒト歯根膜線維芽細胞を使用した.P. gingivalis 由来の LPS は宿主細胞の細胞膜 表面に存在する Toll 様受容体 (Toll-like Receptor, TLR) 4 (TLR4) を介して炎症性サイ トカインの分泌をするなど様々な生理作用を引き起こすといわれている(Nociti FH Jr et al., 2004) .LPS の細胞毒性についての報告も多数みられ(Bailey MT et al., 2009 ; Johansson A et al., 1996; Yiemwattana I & Kaomongkolgit R, 2015) ,LPS 10 µg/mL の濃度 で 作 用 さ せ る と 24 時 間 以 降 で 線 維 芽 細 胞 に 毒 性 を 示 し た と の 報 告 が あ る
(Yiemwattana I & Kaomongkolgit R, 2015) .今回行った細胞毒性試験では, LPS 1
µg/mL を用いた 72 時間および 144 時間という短期間での刺激では細胞に対する毒性
はみられなかったが,そのまま1ヶ月間継続すると, 1 ヶ月が経過する前に細胞は死 滅した.細胞毒性に関する研究は長くても 1 週間程度の期間で行っているものが多く 1 ヶ月間という長期の細胞毒性については検討されていない(Bailey MT et al., 2009 ; Johansson A et al., 1996; Yiemwattana I & Kaomongkolgit R, 2015) .短期間では毒性のみ られなかった濃度 1 µg/mL でも,細胞へのダメージが蓄積されることで毒性を示すこ とが推測された.そこで,LPS の継続的な刺激から,培養液の交換に伴い3日毎に添 加と非添加を繰り返す間欠的な刺激としたところ,細胞毒性を伴わずに 1 か月間長期 培養を行うことに成功した.メカニカルストレスによるメチル化変化を観察するため に,長期にわたり間欠的な刺激を加えて報告があることから(Nemoto T et al., 2010) , 本研究のような間欠的で長期的な刺激を行う実験系は,より安定した DNA のメチル 化変化を観察するのに有用なものと考えられた.
2.DNA メチル化状態の変化について
微生物感染,特にウイルス感染が細胞の DNA メチル化状態に影響を及ぼすとの報 告は多いが(Milavetz BI & Balakrishnan L, 2015) ,細菌感染による影響に関する報告は 少ない(Pérez-Novo CA & Bachert C, 2015; Santos JC & Ribeiro ML, 2015; Ushijima T, 2007) .その中で胃がんの発症に関わる Helicobacter pylori による Lysyl oxidase などの DNA メチル化が比較的多く報告されている(Santos JC & Ribeiro ML, 2015; Ushijima T,
2007) .歯周病原菌が直接的に DNA メチル化に変化を及ぼしたとの報告も僅かながら
なされている.すなわち,Treponema denticola との共培養により歯根膜線維芽細胞の MMP2 遺伝子の脱メチル化を引き起こすことや(Miao D et al., 2014) ,Fusobacterium
nucleatum 刺激により口腔上皮細胞の DNA メチル化酵素 DNMT1 の発現低下とそれに
9
伴う Human β-defensin 2 (HBD2) 遺伝子,CC chemokine ligand 20 (CCL20) 遺伝子の高 メチル化を引き起こすこと(Yin L & Chung WO, 2011) , P. gingivalis 由来 LPS 刺激に より DNA メチル化酵素 DNA methyltransferase (DNMT) 1, DNMT3a の発現低下を引き 起こし(de Camargo Pereira G et al., 2013) , Runt-related transcription factor 2 (RUNX2) 遺 伝子の高メチル化を引き起こすなどがある(Uehara O et al., 2014) .歯周病患者から採 取した組織では MMP25 の高メチル化が起こるとの報告もなされている(Barros SP &
Offenbacher S, 2014) .本研究は,歯周病原菌による DNA メチル化を網羅的に観察し
た最初の文献である.
エピジェネティックな化学的修飾において,可逆的であるヒストン修飾はヘテロク ロマチンの局所的形成を引き起こすといわれ,DNA メチル化は安定的で長期的に発 現を抑制するといわれている(Cedar H & Bergman Y, 2009) .また,外部刺激によるメ チル化の変化は,DNA メチル化酵素が作用しやすい間隙を持つ遺伝子に対してラン ダムに起こるといわれているため(Ushijima T & Asada K, 2009) ,歯周病原菌によって 起こるメチル化変化は,メチル化酵素が作用しやすい遺伝子から起こるものと推測さ れる.本研究では,歯周病原菌によって起こるメチル化変化をより詳細に観察するた め,P. gingivalis 由来 LPS により長期刺激を行い,網羅的に DNA メチル化解析した.
その結果 4 倍以上の高メチル化でも, 1/4 倍以下の低メチル化でも 3,000 種類前後の遺 伝子に変化がみられ,高メチル化したものの中には,DNMT1, DNMT3a, MMP25 遺伝 子なども含まれており,先述した文献とも一致している(de Camargo Pereira G et al., 2013; Barros SP & Offenbacher S, 2014) .このことから,P. gingivalis をはじめとした歯 周病原菌でもかなりの種類の遺伝子にメチル化の変化を引き起こし得るものと考え らえた.
3.高メチル化に伴う mRNA の発現の低下がみられた細胞外マトリックス関連遺伝子
について
細胞外マトリックス関連では,25 遺伝子中 9 遺伝子(FANK1, COL4A1-A2, COL12A1, COL15A1, LAMA5, LAMB1, MMP25, POMT1, EMILIN3)で,高メチル化に伴う mRNA 発現の有意な低下が認められた.これら 9 遺伝子は,歯周組織において生理的恒常性 の維持に重要な役割を果たす機能を持つと推測される.
FANK1 は,アンキリンリピートドメインを含むⅢ型フィブロネクチンをコードする
遺伝子である.フィブロネクチン(FN)はインテグリンと結合し,細胞接着や増殖,
遊走,分化に関与し(Halper J & Kjaer M, 2014) ,骨芽細胞の活性にも重要な役割を担 っている(Hosoya S et al., 1998) . FN の DNA メチル化は,頭頸部や肺の悪性腫瘍でみ られるとの報告があり(Ao M et al., 2015; Pereira IT et al., 2014) ,腫瘍細胞の浸潤の際 にメチル化を介した FN の発現低下が結合組織の破壊に関与しているといわれている
(Ao M et al., 2015) .本研究では,LPS 長期刺激によりプロモーター領域の高メチル
化を介した FN の発現低下が生じており,細胞接着力の低下や骨芽細胞の活性低下に
10
よる歯周組織の恒常性の維持・安定に悪影響を及ぼすものと考えられる.
COL4A1-A2 は, IV 型コラーゲンをコードする遺伝子である. IV 型コラーゲンは基
底膜に存在するコラーゲンとしてよく知られているが,皮膚などの線維芽細胞からも 発 現 し て お り , コ ラ ー ゲ ン ネ ッ ト ワ ー ク の 形 成 に 関 与 し て い る と さ れ て い る
(Ricard-Blum S, 2011) .また, IV 型コラーゲンは,矯正力等のメカニカルストレスで 発現が上昇すると言われているが(Anastasi G, 2008) ,in vitro の実験において過度の メカニカルストレスによって DNA の高メチル化が起こり,遺伝子発現が低下すると の報告もある(Nemoto T et al., 2010) .LPS 刺激による COL4A1 の発現低下では,コ ラーゲンネットワークの形成不全によるメカニカルストレス等への抵抗力の減少に つながると考えられる.
COL12A1 は,XII 型コラーゲンをコードする遺伝子である.XII 型コラーゲンは歯
根膜に存在するコラーゲンとして知られており,メカニカルストレスで発現が上昇し,
歯根膜線維の保護に関与している(Karimbux NY & Nishimura I, 1995; Uno K et al.,
2001) .COL12A1 の DNA メチル化に関する報告は示されていないが,突然変異によ
り骨格異常と筋力低下を起こすエーラース・ダンロス/ミオパチー重複症候群の原因 となることが報告されており(Chiquet M, 2014) , COL12A1 ノックアウトマウスでは,
骨芽細胞の分化抑制と骨タンパクの発現低下がみられたとの報告がある(Chiquet M,
2014) .LPS の長期刺激による COL12A1 の DNA 高メチル化で,メカニカルストレス
への抵抗力の減少や骨芽細胞の分化抑制を招くものと考えられる.
COL15A1 は, XV 型コラーゲンをコードする遺伝子である.このコラーゲンは,腎臓
や膵臓,胎盤など全身のあらゆる部分で発現しており,特に基底膜の最も外層にある 基底膜緻密層に存在しているとされている(Clementz AG & Harris A, 2013) . COL15A1 は扁平上皮癌では DNA メチル化による発現低下が起こり,上皮下へのがん細胞の浸 潤に関わると報告されている(Clementz AG & Harris A, 2013) .LPS 長期刺激による
COL15A1 の発現低下では,歯周炎におけるポケット上皮の Down growth に関わるも
のと推察される.
LAMA5, LAMB1 はラミニンをコードする遺伝子である.線維芽細胞により産生され
たラミニンは,上皮の基底膜の構成要素であり,上皮走化性の誘導や基底膜の接着因 子として機能する(Ohshima M et al., 2006; Woodley DT et al., 1988) .LAMA5, LAMB1 の発現低下は,歯周炎において歯肉上皮の走化性に影響を及ぼすと考えられる.
MMP25 は, Membrane-type matrix metalloproteinase 6 として知られるタンパク質分解酵 素である.MMP25 は,線維芽細胞のサイトカイン産生のトリガーとなり,炎症や細 胞遊走,がん細胞の浸潤を起こすといわれている(Mäkinen LK et al., 2014; Uitto VJ et
al., 2003) .また, in vivo 研究において歯周炎患者の歯肉上皮におけるマイクロアレイ
解析でも MMP25 のメチル化が確認されており(Mäkinen LK et al., 2014; Uitto VJ et al.,
2003) ,サイトカイン産生を促進し炎症を助長するものと考えられる.
POMT1 は,分泌タンパク質のセリンまたはスレオニン残基のマンノースの付加を触
11
媒となる protein-O-mannosyltransferase 1 をコードする遺伝子である(Manya H et al.,
2004).POMT1 の高メチル化とヒトの疾患についての報告は示されていないが,
POMT1 遺伝子の変異によるウォーカー・ワールブルク症候群という脳や目の異常を
伴う筋ジストロフィーが報告されている(Manya H et al., 2004; Saredi S et al., 2012) . また,POMT1 の変異は線維芽細胞の遊走に関与するヒトウロキナーゼ型プラスミノ ゲン活性化因子の分泌を増強されるといわれている(Wilcox-Adelman SA et al., 2000) . DNA メチル化による POMT1 の発現低下は,歯周炎における線維芽細胞の遊走に抑制 的に働くものと考えられる.
EMILIN3 は,エラスチン中に含まれるミクロフィブリルという糖タンパクをコードす
る遺伝子である(Colombatti et al., 2012) .EMILIN3 には細胞の分化・増殖さらに細胞 機能の調節に関与する TGF-βへの拮抗作用や(Schiavinato A et al., 2012) ,EMILIN3 のアイソフォームである EMILIN5 のように骨やセメント質の形成に関与する可能性 がある(Doi M et al., 2004) . DNA メチル化を介した EMILIN3 の発現低下により,様々 な細胞の増殖・分化の抑制や,骨やセメント質の形成に抑制的に働くものと考えられ る.
4.高メチル化に伴い mRNA の発現低下がみられた老化抑制関連遺伝子について
老化抑制関連では 4 遺伝子中 1 遺伝子(KL)において高メチル化に伴う mRNA 発現 の有意な低下が認められた.KL は,生体内で老化を抑制する働きを持つとされるタ ンパク質をコードする遺伝子である.主として腎尿細管,パラトルモンを発現してい る副甲状腺の主細胞,脳脈絡膜などに発現し,欠損マウスでは短命であると報告され ている(Bian A et al., 2015; Sopjani M et al., 2015) .Klotho は FGF19 ファミリーの一つ
である FGF23 と FGFR4 の結合を触媒し,生体内もしくは組織でのカルシウム(Ca)代
謝を調整する働きがあり,発現が低下するとカルシウム沈着による動脈硬化を引き起 こすことが示唆されている(Ding HY & Ma HX, 2015) .また,腎臓での循環器疾患を もつヒトやマウスでも, KL 発現に減少が認められている(Aizawa et al, 1998; Sakan H,
2014) .本研究における KL の mRNA 発現の減少幅は先述のした文献より少ないが
(Aizawa et al, 1998; Sakan H, 2014) ,歯周病に罹患すると動脈硬化のリスクとなるこ とが知られており(Bartova J et al., 2014) ,歯周病原菌による Klotho 遺伝子の高メチル 化とそれに伴う発現低下が,その一因を担っていると推測される.
5.LPS 長期刺激による HPDLFs のタンパク質の発現変化について
タンパク質の発現解析は入手の可能であった FANK1 抗体,COL12A1 抗体,KL 抗体
を用いた CELISA 法により解析した. FANK1, COL12A1, KL タンパク質産生量の減少
がみられたことから,少なくともこれらの遺伝子については DNA 高メチル化がタン
パク質発現減少を介した機能的な変化を引き起こし,歯周病の発症や進行に影響を及
ぼしているものと考えられた.また, mRNA の低下のみられた他の遺伝子でも機能
12
的な変化がみられるものと推測される.他の遺伝子についても抗体を入手しさらなる 検討が必要である.
6.脱メチル化処理による mRNA の発現回復について
脱メチル化剤である 5-Aza 処理にて, mRNA 発現とタンパク質発現の回復が認められ た.これは脱メチル化剤である 5-Aza 処理により,LPS 長期刺激により生じた CpG
islands の異常な DNA 高メチル化が改善されたためと考えられる.しかし,FANK1,
EMILIN3, KL の 3 遺伝子おいては脱メチル化処理を行っても完全な発現回復までは至
らなかった.これら 3 遺伝子の発現制御には,エピジェネティクス以外の調整因子が 関わっている可能性が考えられる.
7.本研究の臨床的意義
本研究結果から,歯周炎の進行に, 歯周病原菌による歯周組織の高メチル化を介し た機能的変化が関与していることが示唆された.一旦高メチル化された細胞は,その まま高メチル化のままで安定に維持されることから(Cedar H & Bergman Y, 2009) ,そ の後にプラーク除去を基本とする歯周治療を行っても,高メチル化された DNA は改 善されないままであり,治療の予後に影響を及ぼすことが推察される.しかしながら,
DNA 高メチル化は脱メチル化剤を用いることにより元の状態に戻り得ることから,
脱メチル化剤による処理が効果的であるものと思われる.本研究で脱メチル化剤とし
て用いた 5-Aza は, DNA メチル基転移酵素(DNMT)を阻害することにより脱メチル化
を起こす薬剤であり,日本では骨髄異形成症候群に対するエピゲノム薬として 2011 年に承認されている(Ishikawa, 2014).さらに,急性骨髄性白血病などの血液腫瘍の治 療に対して全身投与を行うなどの臨床応用が試みられている(Kim TK et al., 2015;
Montalban-Bravo G & Garcia-Manero G, 2015).腫瘍に対する全身投与治療薬として承 認されてから期間も短く,局所投与による治療は未だ行われていないが,今回の研究
結果で 5-Aza による発現回復が認められたことから,今後歯周治療に対しての新たな
局所治療薬としての可能性が期待される.そのためには,本研究のような in vitro で
の研究にとどまらず in vivo での研究や臨床研究を含めた更なる検討が必要である.
13
Ⅴ.結 論
本研究では,歯周病原性細菌 P. gingivalis 由来の LPS を用いた線維芽細胞に対し長期 にわたり継続的に刺激を加える,新たな歯周組織の DNA メチル化解析のための実験 モデルを確立した.
この実験モデルにより,細胞外マトリックスと老化抑制関連遺伝子の中で mRNA や タンパク質の転写活性に影響を及ぼす高メチル化 DNA を同定した.また,脱メチル
化剤 5-Aza により発現回復が認められた.これらの発現低下が歯周組織の恒常性の維
持を破綻させ,老化を促進し,歯周疾患の発症・進行に影響を及ぼすことが示唆され
た.さらに,これらの DNA メチル化の改善をターゲットとした新たな歯周治療に対
する応用できる可能性が見出された.
14
Ⅵ.謝辞
本稿を終えるにあたり,研究精進のため御指導,御高閲を賜りました北海道医療大学 歯学部生体機能・病態学系臨床口腔病理学分野 安彦 善裕 教授に感謝の意を表しま す.また,本研究を行うにあたり,ご助言,ご協力いただきました口腔構造・機能発 育学系保健衛生学分野 千葉 逸朗 教授,口腔生物学系生化学分野 田隈 泰信 教授,
個体差健康科学研究所人類遺伝学分野 太田 亨 教授,臨床口腔病理学分野 西村
学子 講師,佐藤 惇 講師,吉田 光希 助教,保健衛生学分野 植原 治 助教をは
じめ,諸先生に心よりお礼申し上げます.
15
Ⅶ.文 献
Abiko Y, Uehara O, Fukumoto S, Ohta T. Epigenetics of oral infection and inflammatory diseases - DNA methylation changes in infections and inflammation diseases. J Oral Biosci 56: 105-109, 2014.
Aizawa H, Saito Y, Nakamura T, Inoue M, Imanari T, Ohyama Y, Matsumura Y, Masuda H, Oba S, Mise N, Kimura K, Hasegawa A, Kurabayashi M, Kuro-o M, Nabeshima Y, Nagai R.
Downregulation of the Klotho gene in the kidney under sustained circulatory stress in rats.
Biochem Biophys Res Commun. 249(3):865-71, 1998
Anastasi G. An immunohistochemical, histological, and electron-microscopic study of the human periodontal ligament during orthodontic treatment. Int J Mol Med 21: 545-554, 2008.
Ao M, Brewer BM, Yang L, Franco Coronel OE, Hayward SW, Webb DJ, Li D. Stretching fibroblasts remodels fibronectin and alters cancer cell migration. Sci Rep 5: 8334-8341, 2015.
Arai M, Yokosuka O, Hirasawa Y, Fukai K, Chiba T, Imazeki F, Kanda T, Yatomi M, Takiguchi Y, Seki N, Saisho H, Ochiai T. Sequential gene expression changes in cancer cell lines after treatment with the demethylation agent 5-Aza-2'-deoxycytidine. Cancer 106(11):
2514-25, 2006.
Bailey MT, Kinsey SG, Padgett DA, Sheridan JF, Leblebicioglu B. Social stress enhances IL-1beta and TNF-alpha production by Porphyromonas gingivalis lipopolysaccharide-stimulated CD11b+ cells. Physiol Behav 98(3): 351-8, 2009.
Barros SP, Offenbacher S. Modifiable risk factors in periodontal disease: epigenetic regulation of gene expression in the inflammatory response. Periodontol 2000 64: 95-110, 2014.
Bartova J, Sommerova P, Lyuya-Mi Y, Mysak J, Prochazkova J, Duskova J, Janatova T, Podzimek S. Periodontitis as a risk factor of atherosclerosis. J Immunol Res 2014: 636893, 2014
Baylin SB, Jones PA. A decade of exploring the cancer epigenome - biological and translational implications. Nat Rev Cancer 11: 726-734, 2011.
Ben-Avraham D. Epigenetics of aging. Adv Exp Med Biol 847:179-91, 2015.
Bian A, Neyra JA, Zhan M, Hu MC. Klotho, stem cells, and aging. Clin Interv Aging 10:
1233-43, 2015.
Cedar H, Bergman Y. Linking DNA methylation and histone modification: patterns and paradigms. Nat Rev Genet 10(5): 295-304, 2009
Chiquet M. Collagen XII: Protecting bone and muscle integrity by organizing collagen fibrils.
Int J Biochem Cell Biol 53: 51-54, 2014.
Clementz AG, Harris A. Collagen XV: exploring its structure and role within the tumor microenvironment. Mol Cancer Res 11: 1481-1486, 2013.
Colombatti A, Spessotto P, Doliana R, Mongiat M, Bressan GM, Esposito G. The
16
EMILIN/Multimerin family. Front Immunol 2: 93, 2012.
Cullinan MP, Seymour GJ. Periodontal disease and systemic illness: will the evidence ever be enough? Periodontol 2000 62(1):271-86, 2013.
Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell 150: 12-27, 2012.
de Camargo Pereira G, Guimarães GN, Planello AC, Santamaria MP, de Souza AP, Line SR, Marques MR. Porphyromonas gingivalis LPS stimulation downregulates DNMT1, DNMT3a, and JMJD3 gene expression levels in human HaCaT keratinocytes. Clin Oral Investig 17(4):
1279-85, 2013.
Ding HY, Ma HX. Significant roles of anti-aging protein klotho and fibroblast growth factor23 in cardiovascular disease.J Geriatr Cardio 12(4): 439-47 l, 2015.
Ding PH, Jin LJ. The role of lipopolysaccharide-binding protein in innate immunity: a revisit and its relevance to oral/periodontal health. J Periodontal Res 49: 1-9, 2014.
Doi M, Nagano A, Nakamura Y. Molecular cloning and characterization of a novel gene, EMILIN-5, and its possible involvement in skeletal development. Biochem Biophys Res Commun 313(4): 888-893, 2004.
Doyle CJ, Fitzsimmons TR, Marchant C, Dharmapatni AA, Hirsch R, Bartold PM.
Azithromycin suppresses P. gingivalis LPS-induced pro-inflammatory cytokine and chemokine production by human gingival fibroblasts in vitro. Clin Oral Investig 19(2):221-7, 2015.
Egger G, Liang G, Aparicio A, Jones PA. Epigenetics in human disease and prospects for epigenetic therapy. Nature 429: 457-463, 2004.
Gasche JA, Goel A. Epigenetic mechanisms in oral carcinogenesis. Future Oncol 8(11):
1407-25, 2012.
Halper J, Kjaer M. Basic components of connective tissues and extracellular matrix: elastin, fibrillin, fibulins, fibrinogen, fibronectin, laminin, tenascins and thrombospondins. Adv Exp Med Biol 802: 31-47, 2014.
Hosoya S, Suzuki H, Yamamoto M, Kobayashi K, Abiko Y. Alkaline phosphatase and type I collagen gene expressions were reduced by hydroxyl radical-treated fibronectin substratum.
Mol Genet Metab 65: 31-34, 1998.
Ishikawa T. Novel therapeutic strategies using hypomethylating agents in the treatment of myelodysplastic syndrome. Int J Clin Oncol 19(1):10-5, 2014.
Johansson A, Bergenholtz A, Holm SE. Strong cytotoxicity to human gingival fibroblasts by Porphyromonas gingivalis ATCC 33277. J Periodontal Res 31: 477-482, 1996.
Jones KJ, Ekhlassi S, Montufar-Solis D, Klein JR, Schaefer JS. Differential cytokine patterns
in mouse macrophages and gingival fibroblasts after stimulation with porphyromonas
gingivalis or Escherichia coli lipopolysaccharide. J Periodontol 81(12): 1850-7, 2010.
17
Karimbux NY, Nishimura I. Temporal and spatial expressions of type XII collagen in the remodeling periodontal ligament during experimental tooth movement. J Dent Res 74:
313-318, 1995.
Kim TK, Gore SD, Zeidan AM. Epigenetic Therapy in Acute Myeloid Leukemia: Current and Future Directions. Semin Hematol 52(3):172-83, 2015.
Larsson L, Castilho RM, Giannobile WV. Epigenetics and its Role in Periodontal Diseases - A State-of-the-Art Review. J Periodontol 86(4):556-68, 2015 Apr.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25: 402-408, 2001.
Lu L, Katsaros D, de la Longrais IA, Sochirca O, Yu H. Hypermethylation of let-7a-3 in epithelial ovarian cancer is associated with low insulin-like growth factor-II expression and favorable prognosis. Cancer Res 67: 10117-10122, 2007.
Mäkinen LK, Häyry V, Hagström J, Sorsa T, Passador-Santos F, Keski-Säntti H, Haukka J, Mäkitie AA, Haglund C, Atula T. Matrix metalloproteinase-7 and matrix metalloproteinase-25 in oral tongue squamous cell carcinoma. Head Neck 36:1783-1788, 2014.
Manya H, Chiba A, Yoshida A, Wang X, Chiba Y, Jigami Y, Margolis RU, Endo T.
Demonstration of mammalian protein O-mannosyltransferase activity: coexpression of POMT1 and POMT2 required for enzymatic activity. Proc Natl Acad Sci USA 101: 500-505, 2004.
Mascolo M, Siano M, Ilardi G, Russo D, Merolla F, De Rosa G, Staibano S. Epigenetic disregulation in oral cancer. Int J Mol Sci 13(2):2331-53, 2012.
Miao D, Godovikova V, Qian X, Seshadrinathan S, Kapila YL, Fenno JC. Treponema denticola upregulates MMP-2 activation in periodontal ligament cells: interplay between epigenetics and periodontal infection. Arch Oral Biol 59(10): 1056-64, 2014.
Milavetz BI, Balakrishnan L. Viral epigenetics. Methods Mol Biol 1238: 569-596, 2015.
Montalban-Bravo G, Garcia-Manero G. Novel drugs for older patients with acute myeloid leukemia. Leukemia 29(4):760-9, 2015.
Nanbara H, Wara-aswapati N, Nagasawa T, Yoshida Y, Yashiro R, Bando Y, Kobayashi H, Khongcharoensuk J, Hormdee D, Pitiphat W, Boch JA, Izumi Y. Modulation of Wnt5a expression by periodontopathic bacteria. PLoS One 7(4):e34434, 2012.
Nemoto T, Kajiya H, Tsuzuki T, Takahashi Y, Okabe K. Differential induction of collagens by mechanical stress in human periodontal ligament cells. Arch Oral Bio 55: 981-987, 2010.
Nociti FH Jr, Foster BL, Barros SP, Darveau RP, Somerman MJ. Cementoblast gene expression is regulated by Porphyromonas gingivalis lipopolysaccharide partially via toll-like receptor-4/MD-2. J Dent Res 83: 602-607, 2004.
Ohshima M, Yamaguchi Y, Otsuka K, Sato M, Ishikawa M. Laminin expression by human
periodontal ligament fibroblasts. Connect Tissue Res 47: 149-156, 2006.
18
Pereira IT, Ramos EA, Costa ET, Camargo AA, Manica GC, Klassen LM, Chequin A, Braun-Prado K, Pedrosa Fde O, Souza EM, Costa FF, Klassen G. Fibronectin affects transient MMP2 gene expression through DNA demethylation changes in non-invasive breast cancer cell lines. PLoS One 9(9): e105806, 2014.
Pérez-Novo CA, Bachert C. DNA methylation, bacteria and airway inflammation: latest insights. Curr Opin Allergy Clin Immunol 15(1): 27-32, 2015.
Potempa J, Banbula A, Travis J. Role of bacterial proteinases in matrix destruction and modulation of host responses. Periodontol 2000 24: 153-92, 2000
Ramos MP, Wijetunga NA, McLellan AS, Suzuki M, Greally JM.DNA demethylation by 5-aza-2′-deoxycytidine is imprinted, targeted to euchromatin, and has limited transcriptional consequences. Epigenetics Chromatin 8: 11-27, 2015.
Ricard-Blum S. The collagen family. Cold Spring Harb Perspect Biol 3: a004978, 2011.
Sakan H, Nakatani K, Asai O, Imura A, Tanaka T, Yoshimoto S, Iwamoto N, Kurumatani N, Iwano M, Nabeshima Y, Konishi N, Saito Y. Reduced renal α-Klotho expression in CKD patients and its effect on renal phosphate handling and vitamin D metabolism. PLoS One;9(1): e86301, 2014
Santos JC, Ribeiro ML. Epigenetic regulation of DNA repair machinery in Helicobacter pylori-induced gastric carcinogenesis. World J Gastroenterol 21(30): 9021-37, 2015.
Saredi S, Ardissone A, Ruggieri A, Mottarelli E, Farina L, Rinaldi R, Silvestri E, Gandioli C, D'Arrigo S, Salerno F, Morandi L, Grammatico P, Pantaleoni C, Moroni I, Mora M. Novel POMGNT1 point mutations and intragenic rearrangements associated with muscle-eye-brain disease. J Neurol Sci 318: 45-50, 2012.
Schiavinato A, Becker AK, Zanetti M et al. EMILIN-3, peculiar member of elastin microfibril interface-located protein (EMILIN) family, has distinct expression pattern, forms oligomeric assemblies, and serves as transforming growth factor β (TGF-β) antagonist. J Biol Chem 287:
11498-11515, 2012.
Sopjani M, Rinnerthaler M, Kruja J, Dermaku-Sopjani M. Intracellular signaling of the aging suppressor protein Klotho. Curr Mol Med 15(1): 27-37, 2015.
Uehara O, Abiko Y, Saitoh M, Miyakawa H, Nakazawa F. Lipopolysaccharide extracted from Porphyromonas gingivalis induces DNA hypermethylation of runt-related transcription factor 2 in human periodontal fibroblasts. J Microbiol Immunol Infect 47(3):176-81, 2014.
Uitto VJ, Overall CM, McCulloch C. Proteolytic host cell enzymes in gingival crevice fluid.
Periodontol 2000 31: 77-104, 2003.
Uno K, Abiko Y, Takita H, Ukegawa T, Satoh N, Ueda Y, Ohata N, Kaku T, Kuboki Y. Effects of mechanical stress on the expression of type XII collagen mRNA in human periodontal ligament cells. J Hard Tissue Biol 10: 116-122, 2001.
Ushijima T, Asada K. Aberrant DNA methylation in contrast with mutations. Cancer Sci
19
101(2): 300-5, 2010.
Ushijima T. Epigenetic field for cancerization. J Biochem Mol Biol 40(2): 142-50, 2007.
Wen WC, Kuo PJ, Chiang CY, Chin YT, Fu MM, Fu E. Epigallocatechin-3-gallate attenuates Porphyromonas gingivalis lipopolysaccharide-enhanced matrix metalloproteinase-1 production through inhibition of interleukin-6 in gingival fibroblasts. J Periodontol 85(6):868-75, 2014
Wilcox-Adelman SA, Wilkins-Port CE, McKeown-Longo PJ. Localization of urokinase type plasminogen activator to focal adhesions requires ligation of vitronectin integrin receptors.
Cell Adhes Commun 7: 477-490, 2000.
Woodley DT, Stanley JR, Reese MJ, O'Keefe EJ. Human Dermal Fibroblasts Synthesize Laminin. J Invest Dermatol 90: 679-683, 1988.
Yiemwattana I, Kaomongkolgit R. Alpha-mangostin suppresses IL-6 and IL-8 expression in P.
gingivalis LPS-stimulated human gingival fibroblasts. Odontology 103(3): 348-55, 2015.
Yin L, Chung WO. Epigenetic regulation of human β-defensin 2 and CC chemokine ligand 20 expression in gingival epithelial cells in response to oral bacteria. Mucosal Immuno l4(4):
409-19, 2011.
前田勝正,濵地貴文:第 1 章 歯周病の原因と症状.臨床歯周病学 第 2 版.医歯薬 出版:2013,18-22.
宮下元:第 12 章 高齢者と有病者の治療.臨床歯周病学 第 2 版.医歯薬出版: 2013,
342-348.
表 ・ 付図
表 1 mRNA 発現解析に用いたプライマー配列
Name Sequences Size
FANK1-F 5’-AAGGAGGCCGTGTTAAGGTT-3’ 168 bp
FANK1-R 5’-CATAGCACGCCAGCATTAGA-3’
POMT1-F 5’-CTTCAACTGCCAAAAGCACA-3’ 202 bp
POMT1-R 5’-GCACTGGACATCAGCACCTA-3’
COL12A1-F 5’-AGGTCATGATTCTTATCATCACGG-3’ 286 bp COL12A1-R 5’-AGATCCTTTGGAGGAACGTAA-3’
COL4A1,2-F 5’-CTGGTCCAAGAGGATTTCCA-3’ 193 bp
COL4A1,2-R 5’-TCATTGCCTTGCACGTAGAG-3’
LAMA5-F 5’-TGACCTTTTCTGGCTCGTCT-3’ 202 bp
LAMA5-R 5’-GTTCAGCACAAAGGGCTCTC-3’
LAMB1-F 5’-AACGTGGTTGGAAGAACCTG-3’ 151 bp
LAMB1-R 5’-ACACTCCCTGGAAACAGTGG-3’
MMP25-F 5’-GACCTGGACTTTTGGGTCAA-3’ 171 bp
MMP25-R 5’-TCCTGAGACAGGCGGTACTT-3’
COL15A1-F 5’-GTTGTCCACCTACCGAGCAT-3’ 242 bp
COL15A1-R 5’-AGCCATGCCAAATGACTTTC-3’
EMILIN3-F 5’-GGTTTGTGCAGGAAGGTGAT-3’ 182 bp
EMILIN3-R 5’-GGGGAGGATCAAAGGGAATA-3’
KL-F 5’-AGGGTCCTAGGCTGGAATGT-3’ 158 bp
KL-R 5’-CCTCAGGGACACAGGGTTTA-3’
表 2 MSP 解析に用いたプライマー配列
Name Sequences(小文字がCpG Position) Size Target Region FANK1-Methyl-F 5’-GTTTTcGcGATTTAGGAGAc-3’ 173
bp -936~
-723 FANK1-Methyl-R 5’-AACgAAACCCTACgAACAAC-3’
FANK1-Unmethyl-F 5’-GGTTTTtGtGATTTAGGAGAt-3’ 173
bp -937~
-722 FANK1-Unmethyl-R 5’-AAACaAAACCCTACaAACAAC-3’
POMT1-Methyl-F 5’-TTGTGTGGATTcGGAGAAAc-3’ 149
bp -171~
POMT1-Methyl-R 5’-CCAACTCCgAAAAAACgAA-3’ +16
POMT1-Unmethyl-F 5’-GTTTTGTGTGGATTtGGAGAAAt-3’ 149
bp -174~
+19 POMT1-Unmethyl-R 5’-CCCCAACTCCaAAAAAACaAAA-3’
COL12A1-Methyl-F 5’-AAAGATAGAAGGcGcGTc-3’ 171
bp -915~
-702 COL12A1-Methyl-R 5’-TCgAAAATACAAAAATACgTAACC-3’
COL12A1-Unmethyl-F 5’-GAAAAAGATAGAAGGtGtGTt-3’ 171
bp -918~
-700 COL12A1-Unmethyl-R 5’-TTTCaAAAATACAAAAATACaTAACCC-3’
COL4A1,2-Methyl-F 5’-GGTTTTAAGcGGAGATTTGAGc-3’ 100
bp -77~
+62 COL4A1,2-Methyl-R 5’-CTCCCCTTTAAAACgCCg-3’
COL4A1,2-Unmethyl-F 5’-GGTTTTAAGtGGAGATTTGAGt-3’ 100 bp -77
+62~
COL4A1,2-Unmethyl-R 5’-CTCCCCTTTAAAACaCCa-3’
LAMA5-Methyl-F 5’-cGTATAGAAGTGATTATGGGTTTAGc-3’ 151
bp -6536~
-6363 LAMA5-Methyl-R 5’-CgAACCCCTACgAAACTAAATA-3’
LAMA5-Unmethyl-F 5’-TTAtGTATAGAAGTGATTATGGGTTTAGt-3’ 151
bp -6564~
-6360 LAMA5-Unmethyl-R 5’-CCaAACCCCTACaAAACTAAATATA-3’
LAMB1-Methyl-F 5’-GGAAGATATTGTAGATGTcGc-3’ 154
bp -334~
-140 LAMB1-Methyl-R 5’-CgACTAAAACTCTCgTACgA-3’
LAMB1-Unmethyl-F 5’-TTAGGAAGATATTGTAGATGTtGt-3’ 154
bp -337~
-137 LAMB1-Unmethyl-R 5’-CCACaACTAAAACTCTCaTACaA-3’
MMP25-Methyl-F 5’-cGAGAGATAGGGATTTGGAGATc-3’ 133
bp -9968~
-9793 MMP25-Methyl-R 5’-GACAAAAACTCAACCgTCgAA-3’
MMP25-Unmethyl-F 5’-GtGAGAGATAGGGATTTGGAGATt-3’ 133
bp -9969~
-9791 MMP25-Unmethyl-R 5’-CAACAAAAACTCAACCaTCaAA-3’
COL15A1-Methyl-F 5’-GTTAGcGTTTAGcGATTTTTc-3’ 103
bp -221~
-79 COL15A1-Methyl-R 5’-ACCTTAATACCATCTCGCG-3’
COL15A1-Unmethyl-F 5’-GGTTAGtGTTTAGtGATTTTTt-3’ 103
bp -222~
COL15A1-Unmethyl-R 5’-TACCTTAATACCATCTCaCa-3’ -78
EMILIN3-Methyl-F 5’-AGTTGAGTTTGGGATTGTTTTTAAGc-3’ 187
bp -333~
EMILIN3-Methyl-R 5’-AAAAACCAAAACTATACCCTCCgAA-3’ -96
EMILIN3-Unmethyl-F 5’-TGAGTTTGGGATTGTTTTTAAGtGA-3’ 190
bp -330~
EMILIN3-Unmethyl-R 5’-AAAAACCAAAACTATACCCTCCaAA-3’ -93
KL-Methyl-F 5’- AGTAGGTGTTTTTTTAGCGGC-3’ 169
bp -477~
-268 KL-Methyl-R 5’-AAAACgTTTCgTAAACgCTC-3’
KL-Unmethyl-F 5’GAGAGTAGGTGTTTTTTTAGtGGT -3’ 169
bp -480~
-265 KL-Unmethyl-R 5’-ACAAAACaTTTCaTAAACaCTCA -3’
表 3 メチレーションアレイ解析結果
Probes Genes
4-folds Hypermethylation 12,918 probes 2,910 genes 4-folds Hypomethylation 10,330 probes 3,375 genes
2~5 folds 5~10 folds 10~20 folds 20~ folds Hypermethylation 15,165 probes 9,553 probes 3,961 probes 1,433 probes Hypomethylation 12,257 probes 4,585 probes 3,282 probes 5,784 probes
Promoter Inside Downstream Unknown Hypermethylation 5,887 probes 17,740 probes 1,727 probes 797 probes Hypomethylation 9,291 probes 15,113 probes 1,176 probes 307 probes
表 4 細胞外マトリックス関連 25 遺伝子
Normalizedratio Gene Symbol Description From
transcription start site 63.491898 FANK1 fibronectin type III and ankyrin
repeat domains 1 -917
46.09059 FOXF2 forkhead box F2 -6762
31.689793 COL2A1 collagen, type II, alpha 1 -1171 21.313196 PAPLN papilin, proteoglycan-like sulfated
glycoprotein -61
16.347855 POMT1 protein-O-mannosyltransferase 1 -152 13.42935 COL24A1 collagen, type XXIV, alpha 1 -786 13.241217 ADAMTS7 ADAM metallopeptidase with
thrombospondin type 1 motif, 7 -863 11.571117 COL2A1 collagen, type II, alpha 1 -879 11.541119 MMP21 matrix metallopeptidase 21 -74
10.859601 FOXF2 forkhead box F2 -6096
10.206867 FOXF2 forkhead box F2 -6820
9.621575 GPC4 glypican 4 -704
8.6972885 COL12A1 collagen, type XII, alpha 1 -898 7.951499 FANK1 fibronectin type III and ankyrin
repeat domains 1 -798
7.2812815 NTN2L netrin 2-like (chicken) -3399
7.2207212 FOXF2 forkhead box F2 -6014
7.043698 FOXF2 forkhead box F2 -5926
6.756929 ADAMTS16 ADAM metallopeptidase with
thrombospondin type 1 motif, 16 -1123 6.718919 COL2A1 collagen, type II, alpha 1 -960
6.6792035 FOXF2 forkhead box F2 -6526
6.3270826 COL4A1-COL4A2 collagen, type IV, alpha 1 -56 6.1432166 COL2A1 collagen, type II, alpha 1 -1105 5.84885 APLP1 amyloid beta (A4) precursor-like
protein 1 -36
5.648105 FOXF2 forkhead box F2 -5853
5.603655 ADAMTS9 ADAM metallopeptidase with thrombospondin type 1 motif, 9 -593 5.5399475 ADAMTS9 ADAM metallopeptidase with
thrombospondin type 1 motif, 9 -552
5.3450613 FOXF2 forkhead box F2 -7030
5.1396294 ADAMTS5 ADAM metallopeptidase with thrombospondin type 1 motif, 5 -492
4.994581 MATN4-RBPSUHL matrilin 4 -898
4.8238945 COL12A1 collagen, type XII, alpha 1 -784
4.78614 LAMA5 laminin, alpha 5 -6536
4.7603045 LAMB1 laminin, beta 1 -314
4.6712303 LAMB1 laminin, beta 1 -159
4.5837936 CHAD chondroadherin -392
4.5642667 COL6A1 collagen, type VI, alpha 1 -7810 4.5466533 MMP25 matrix metallopeptidase 25 -9946
4.444609 NTN2L netrin 2-like (chicken) 3323
4.3141513 COL15A1 collagen, type XV, alpha 1 -201
4.2577662 LTBP1 netrin G2 -796
4.0327334 EMILIN3 latent transforming growth factor beta binding protein 1 -308
表 5 老化抑制関連 4 遺伝子
Normalizedratio Gene Symbol Description From
transcription start site
29.006138 TBX2 T-box 2 -3874
21.288427 KL Klotho -460
15.312082 KL Klotho -584
14.857672 TBX2 T-box 2 -3768
10.019271 TBX3 T-box 3 -3079
9.454075 TBX3 T-box 3 -2929
9.42881 TBX3 T-box 3 -2768
8.151492 NPM1 Nucleophosmin 1 -132
7.7796607 TBX3 T-box 3 -3240
7.4601097 TBX3 T-box 3 -8044
7.167736 TBX3 T-box 3 -817
7.0751767 TBX3 T-box 3 -3021
5.007787 TBX2 T-box 2 -4165
4.778714 TBX3 T-box 3 -3176
4.4233327 TBX3 T-box 3 -2828
HPDLFs 前培養
(10%FBS 含有 DMEM)
↓
↓
↓
↓ 3 日後
洗浄,培養液交換 LPS,滅菌水添加
↓
↓
↓
↓ 3 日後
洗浄,培養液交換
↓
↓
↓
↓ 3 日後
洗浄,培養液交換 LPS ,滅菌水添加
↓
↓
↓
↓ 3 日後
洗浄,培養液交換
↓
↓
↓ 継続
↓
↓
↓ 1 か月後 LPS 長期刺激 細胞サンプル
CONTROL LPS 図 1 LPS 長期刺激下での細胞培養フローチャート
細胞を前培養後,LPS 添加・非添加培地にて 3 日毎交互に交換しながら 1 ヶ月間培養
した.
図 2 各遺伝子の CpG islands と Methylation array, MSP の関係
: Methylation array での高メチル化部位および MSP Primer の設計位置
図 3 細胞毒性試験結果(n=3)
LPS 添加後 72 時間(3 日間)および 144 時間(6 日間)において, LPS 濃度 10, 100 µg/ml で有意な細胞数の減少がみられた.A:72 時間,B:144 時間
*p<0.05
*p<0.05
図 4 LPS 長期刺激下での HPDLFs の細胞形態(位相差顕微鏡像)
HPdLFs に 1 ヶ月間 LPS 1 µg/ml 刺激を加えても, 細胞に形態的変化はみられなかった.
A:Control 群,B:LPS 添加群
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
図 5 mRNA 発現解析のグラフ(n=3)
Real-time PCR 法による mRNA の発現解析を行ったところ,細胞外マトリックス関連
および老化抑制関連の 10 遺伝子で LPS 長期刺激により有意な発現低下が認められた A :FANK1, B: COL4A1-A2, C: COL12A1, D: COL15A1, E :LAMA5, F :LAMB1,
G:MMP25,H:POMT1,I:EMILIN3,J:KL.
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
図 6 MSP 解析のグラフ(n=3)
MSP 法によるメチレーションレベル解析において, mRNA 発現に有意な低下がみら れた細胞外マトリックス関連および老化抑制関連の 10 遺伝子で,LPS 長期刺激によ りメチレーションレベルの有意な上昇が認められた.
A : FANK1 , B : COL4A1-A2 , C : COL12A1 , D : COL15A1 , E : LAMA5 , F : LAMB1 , G:MMP25,H:POMT1,I:EMILIN3,J:KL.
*p<0.05
*p<0.05
図 7 タンパク質発現解析のグラフ(n=3)
CELISA 法によるタンパク質の発現解析を行ったところ, mRNA の発現低下がみられ
た FANK1, COL12A1, KL において LPS 長期刺激により有意な発現低下が認められた
A:FANK1,B:COL12A1,C:KL.
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
*p<0.05
図 8 脱メチル化解析のグラフ (n=3)
5-Aza により脱メチル化処理を加えたところ, LPS 長期刺激群と比較して mRNA 発現
に有意な回復が認められた.しかし,コントロール群と 5-Aza 添加群を比較すると
FANK1, EMILIN3, KL では有意に発現が低下しており,完全な回復までには至らなか
った.
A :FANK1, B: COL4A1-A2, C: COL12A1, D: COL15A1, E :LAMA5, F :LAMB1,
G : MMP25 , H : POMT1 , I : EMILIN3 , J : KL .
*p<0.05
*p<0.05