Posted at the Institutional Resources for Unique Collection and Academic Archives at Tokyo Dental College, Available from http://ir.tdc.ac.jp/
Title
Down-regulated genes in mouse dental papillae and
pulp
Author(s)
Alternative
Sasaki, H; Muramatsu, T; Kwon, HJ; Yamamoto, H;
Hashimoto, S; Jung, HS; Shimono, M
Journal
Journal of dental research, 89(7): 679-683
URL
http://hdl.handle.net/10130/2408
DOI: 10.1177/0022034510366844
Received October 13, 2008; Last revision February 4, 2010; Accepted February 10, 2010
A supplemental appendix to this article is published elec-tronically only at http://jdr.sagepub.com/supplemental.
H. Sasaki
1,2, T. Muramatsu
1,2*,
H.-J. Kwon
3, H. Yamamoto
4,
S. Hashimoto
1,2, H.-S. Jung
2,3,
and M. Shimono
1,21Oral Health Science Center HRC7, Tokyo Dental College,
Japan; 2Department of Pathology, Tokyo Dental College, 1-2-2
Masago, Mihama-ku, Chiba, 261-8502, Japan; 3Department of
Oral Biology, Research Center for Orofacial Hard Tissue Regeneration, Oral Science Research Center, College of Dentistry, Brain Korea 21 Project for Medical Science, Yonsei University, Korea; and 4Department of Histology, Cytology
and Developmental Anatomy, Nihon University, School of Dentistry at Matsudo, Japan; *corresponding author, [email protected]
J Dent Res 89(7):679-683, 2010
AbstrAct
Important factors involved in odontogenesis in mouse dental papillae disappear between the pre- and post-natal stages of development. Therefore, we hypothesized that certain genes involved in odontogenesis in dental papillae were subject to pre-/post-natal down-regulation. Our goal was to identify, by microarray analysis, which genes were down-regulated. Dental papillae were isolated from embryonic 16-day-, 18-day- (E16, E18), and post-natal 3-day-old (P3) murine first mandibular molar germs and analyzed by microarray. The number of down-regulated genes was 2269 between E16 and E18, and 3130 between E18 and P3. Drastic down-regulation (fold change > 10.0) of Adamts4, Aldha1a2, and Lef1 was observed at both E16 and E18, and quantitative RT-PCR revealed a post-natal reduction in their expression (Adamts4, 1/3; Aldh1a2, 1/13; and Lef1, 1/37). These results suggest that down-regulation of these three genes is an important factor in normal odontogenesis in dental papillae.
KEY WOrDs:
dental papilla, dental pulp, odon-togenesis, gene expression, tooth morphogenesis.IntrODuctIOn
t
ooth morphogenesis is a complex process that has been characterized as a series of reciprocal epithelial-mesenchymal interactions, culminating in differentiation of the interacting tissues. At the molecular level, these interac-tions are mainly regulated by signaling molecules, including bone morpho-genetic proteins (Bmp), fibroblast growth factors (Fgf), Sonic hedgehog and Wnt (Hurmerinta et al., 1980; Jernvall and Thesleff, 2000). The functions of these molecules have been investigated in knockout and transgenic mice, and by experiments with antisense oligonucleotides. However, many of the mol-ecules, signal transcriptions, processes, and functions involved in tooth mor-phogenesis remain to be determined.Classic studies on tissue recombinations have investigated interactions between dental epithelium and dental papillae. Such studies have demon-strated that dental epithelium can induce tooth formation prior to the bud stage. However, after the bud stage, this initiative shifts to the dental mesen-chyme (Kollar and Baird, 1970; Mina and Kollar, 1987). It has been demon-strated that the combination of embryonic 16-day-old (E16) enamel organs and E16 dental papillae formed teeth, while that of E16 enamel organs and post-natal 3-day-old (P3) pulps did not, producing dysplastic dentin only (Palmer and Lumsden, 1987). This led us to hypothesize that certain genes related to odontogenesis were down-regulated in dental papillae after the late bell stage, which corresponds to mouse tooth development between the pre- and post-natal stages. To confirm this hypothesis, we investigated and com-pared the gene profiles of pre- and post-natal-stage dental papillae.
One current advance in biomolecular technology, GeneChip® microarray
technology, allows for precise genetic profiling during tooth morphogenesis. These arrays consist of more than 30,000 oligonucleotide probes positioned in a known location within the target cell. By this method, it is possible for samples to be compared at different stages of tooth development, facilitating the identification of genes and variations in levels of expression during tooth morphogenesis.
The aim of this study was to identify, by microarray analysis, which genes were down-regulated between the pre- and post-natal stages of development in dental papillae.
Down-regulated Genes in Mouse
Dental Papillae and Pulp
680 Sasaki et al. J Dent Res 89(7) 2010
MAtErIAls & MEtHODs
Animals & tissue Preparation
ICR mice were obtained from Sankyo Laboratory Service Corporation (Tokyo, Japan). E16, E18, and P3 mice were killed under deep anesthesia with sodium thiopental. The ages of embryos were determined by confirmation of vaginal plugs on the day after mating and confirmed by external developmental features. Stages of tooth germ development were classified as (1) early bell-stage (E16), (2) late bell-stage (E18), and (3) den-tin and enamel maturation period (P3). All experiments were carried out according to the Guidelines for the Treatment of Animals at Tokyo Dental College.
First-molar tooth germs were dissected from the mandible under a dissecting microscope. After being washed in phos-phate-buffered saline, tooth germs were incubated in 1.2 U/mL dispase I (Roche, Mannheim, Germany) for 5 min at room tem-perature (RT) to separate dental epithelium and mesenchyme, and de-activated in minimum essential medium (MEM, Invitrogen, Grand Island, NY, USA) with 15% fetal bovine serum (Sigma-Aldrich, St. Louis, MO, USA). Dental papillae were isolated from the tooth germs under a dissecting micro-scope with tungsten needles. Dental papillae and pulp tissues obtained at E16, E18, and P3 were used for microarray analysis.
rnA Extraction
Total RNA was extracted from 3 pooled groups of tissue—E16-papillae (n = 20), E18-tissue—E16-papillae (n = 20), and P3-pulp tissues (n = 20)—with TRIzol® (Invitrogen) according to the manufacturer’s
instructions. RNA extractions were performed in duplicate for each of the 3 pooled groups of tissue from each developmental time-point, to provide 2 biological replicates at each time-point for which the tissue pool n = 20 for each replicate at each developmen-tal time-point. The RNA was concentrated from the 20 homoge-nates. RNA samples were quantified spectrophotometrically, and the integrity of the RNA was confirmed with the Agilent 2100 Bioanalyzer (Agilent Technologies, Foster City, CA, USA).
Microarray Hybridization
Double-stranded complementary DNA (cDNA) was synthesized by a two-cycle target-labeling method. A 200-ng quantity of total RNA was converted into first-cycle double-stranded cDNA. As the first cycle, in vitro transcription (IVT) amplification of cRNA was carried out with MEGAscript T7 Kit (Ambion, Austin, TX, USA). As a second-cycle, double-stranded cDNA synthesis was performed with the SuperScript Choice System (Invitrogen). The samples were synthesized into Biotin-labeled cRNA by means of a transcript labeling kit (Enzo Diagnostics, Farmingdale, NY, USA) and fragmented randomly. Fragmented cRNA was hybridized with the GeneChip® Mouse Genome 430
2.0 Array (Affymetrix, Santa Clara, CA, USA) at 45°C for 16 hrs. The microarray chip was analyzed with the Affymetrix®
Fluidic Station 450, and the images scanned. RNA was applied from each of the 2 biological replicates of each pooled tissue
(n = 20 per replicate) at each developmental time-point to the microarray without inclusion of any technical replicates.
Data Analysis
Data were analyzed with the GeneChip® Operating Software
ver-sion 1.1 (GCOS; Affymetrix). The data were subsequently trans-ferred to GeneSpring® software (Silicon Genetics, Redwood City,
CA, USA) for normalization and statistical analysis. Where the software indicated low or unstable gene expression, these data were excluded from further analysis, and only those genes flagged as “present” were selected. From these data, which included unknown genes, we selected only those genes whose difference in expression exhibited a more than two-fold change. Down-regulated genes selected from those data were categorized accord-ing to the Gene Ontology database (GO). Those genes among the three stages (E16, E18, and P3) were then categorized with the GeneSpring® software according to their corresponding
ontolo-gies as follows: Calcification, Development, or Cell Death.
Quantitative real-time Pcr (qrt-Pcr)
Total RNA was reverse-transcribed into complementary DNA (cDNA) with Sensiscript® Reverse Transcript (QIAGEN,
Alameda, CA, USA), and qRT-PCR with TaqMan MGB probes (Applied Biosystems, Foster City, CA, USA) was carried out. The TaqMan MGB probes and primer sets for the mouse genes (Adamts4, Aldh1a2, Lef1) and mouse glyceraldehydes-3-phosphate dehydrogenase (GAPDH), as an internal control, were purchased from Applied Biosystems. Quantification and differentiation of mRNA expression was analyzed with ABI 7700 system software (Applied Biosystems) and compared by the ΔΔCt method. Differences among E16, E18, and P3 were analyzed statistically by the non-parametric Mann-Whitney U-test with Bonferroni correction. Values of p < 0.01 or 0.05 were considered significant.
In situ Hybridization
The upper and lower first molars were dissected from E16, E18, and P3 mice and hybridized in situ with Adamts4, Aldh1a2, and Lef1 RNA probes. Digoxigenin-labeled antisense RNA probes were generated from cDNA subcloned in pBluescript SK with T7 RNA polymerase, and sense probe was done with SP6 poly-merase, as described elsewhere (Cho et al., 2007).
rEsults
Gene Expression Analysis in Dental Papillae by Microarray
We utilized microarray technology to compare multiple gene expression profiles between pre- and post-natal dental papillae. Information and profile of gene expression were submitted to the Gene Expression Omnibus (GEO) database: E16 (GEO accession number GSM485768), E18 (GEO accession number GSM485769), and P3 (GEO accession number GSM485770).
The number of up-regulated and down-regulated genes showing a more than two-fold change between each period is shown in Table 1. More than 2000 genes showed either up- or down-regulation between E16 and E18. The number of up- regulated genes decreased following tooth morphogenesis. In contrast, the largest number of down-regulated genes (3130) was found between E18 and P3.
Down-regulated genes showing a more than four-fold change were categorized according to GO (Appendix Table), and genes showing a more than 10-fold change are listed in Table 2. In the “Calcification” category, expression of Adamts4 and Lef1 decreased considerably (fold change > 10.0) between E16 and E18. In the “Development” category, expression of Aldh1a2 showed a significant reduction (fold change > 10.0) between E16 and E18. In the “Cell Death” category, no gene showed a more than 10-fold change. Moreover, in all categories, no genes showed a more than 10-fold change between E18 and P3.
Based on the results of the microarray analysis, we focused on 3 genes—Adamts4, Aldh1a2, and Lef1—since all of them displayed conspicuous change (> 10-fold change) following tooth germ maturation.
confirmation of Gene Expression by qrt-Pcr
The expression of these 3 genes as measured by qRT-PCR was similar to that obtained with the microarray (Fig. 1). QRT-PCR for Adamts4 confirmed a 3.1-fold reduction in its mRNA expression between E16 and E18 (Fig. 1A), at a statistically significant level (p < 0.05). QRT-PCR demonstrated a 12.8-fold reduction in expression of Aldh1a2 mRNA between E16 and E18 (Fig. 1B), at a statistically significant level (p < 0.01). QRT-PCR indicated a 37.4-fold reduction in expression of Lef1 between E16 and E18, at a statistically significant level (p < 0.01) (Fig. 1C).
localization of Identified Genes by in situ Hybridization
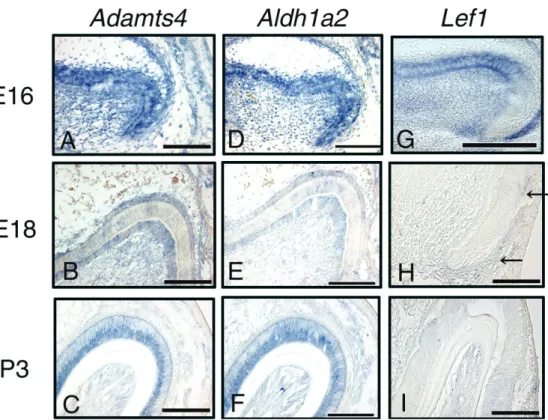
The three genes, Adamts4 (Figs. 2A-2C), Aldh1a2 (Figs. 2D-2F), and Lef1 (Figs. 2G-2I), showed similar patterns of expres-sion during tooth development. At E16, they were localized in inner and outer dental epithelia and dental papilla cells adjacent to the inner dental epithelium (Figs. 2A, 2D, 2G). At E18, local-ization of Aldh1a2 and Adamts4 was also observed in dental epithelia consisting of inner dental epithelium and stratum inter-medium (Figs. 2B, 2E). However, expression of Lef1 showed the greatest decrease, and localization was observed in the outer dental epithelium at the cervical area (Fig. 2H). Expression of Aldh1a2 and Adamts4 became weaker or disappeared in odon-toblasts differentiated from dental papilla cells and pulp cells at P3, but persisted in ameloblasts differentiated from inner dental epithelial cells (Figs. 2C, 2F). No expression of Lef1 was observed in odontoblasts, dental pulp cells, or dental epithelium (Fig. 2I).
DIscussIOn
The tooth is an ectodermal organ like hair, feathers, lungs, and mammary glands, and its formation is controlled by reciprocal
table 1. Number of Up- and Down-regulated Genes Showing Greater than Two-fold Change between Each Period
Up-regulated Genes Down-regulated Genes
Between E16 and E18 2439 2269
Between E18 and P3 1129 3130
Number of up-regulated genes decreased following tooth morphogen-esis. Largest number of down-regulated genes was found between E18 and P3.
interactions between dental epithelium and dental mesenchyme (Thesleff, 2003). However, this interaction is dynamic and com-plex. Many of the molecules, signal transcriptions, processes, and functions that play a part in tooth morphogenesis remain to be characterized, and more than 300 genes have been reported to be involved (http://bite-it.helsinki.fi). High-throughput tech-nologies such as DNA microarrays can comprehensively profile and monitor gene expression in biological processes, including development (Hiratsuka et al., 2002; Brown et al., 2003). In this study, we compared pre- and post-natal gene profiles using microarray technology, and found over 2000 expressed, a higher number than that found in previous reports. Our results also showed that the highest number of down-regulated genes was recognized between E18 and P3, which corresponds to the period between pre- and post-natal mouse dental papillae, when pre-odontoblasts differentiate into mature odontoblasts. Furthermore, down-regulated genes showing a more than four-fold change belonged to the “Development” category (data in Appendix). These results suggest that the expression of develop-ment-related signaling molecules and interactions in dental papillae is quite complicated during tooth development, and that as-yet-unreported genes are involved in the process of tooth development.
The Adamts family is a zinc-dependent metalloproteinase that plays an important role in normal and pathological events (Kuno et al., 1997). It is composed of at least 19 molecules, and Adamts4 (aggrecanase-1) degrades proteoglycans, including aggrecan, versican, and brevican (Nakamura et al., 2000; Tortorella et al., 2000; Westling et al., 2004). Expression of Adamts4 has been identified, by in situ hybridization, in odonto-blasts, pulp cells, and periodontal ligament cells during rat molar eruption (Sone et al., 2005). Furthermore, proteoglycans were found in dental papillae after the early bell stage and in the post-natal dental stage, and may play an important role in dif-ferentiation of odontoblasts (Hikake et al., 2003). In the present study, we found that expression of Adamts4, which degrades proteoglycans, decreased significantly between E16 and E18, suggesting an increase in proteoglycans in the dental papillae. Our results suggest that reduced expression of Adamts4 induces an increase in proteoglycans in dental papillae, leading to dif-ferentiation of odontoblasts.
In the present study, mRNA expression of Aldh1a2 (Raldh2) showed a marked reduction between E16 and E18. Adh1a2 belongs to the aldehyde dehydrogenase family, and has been reported to play a significant role during early embryogenesis of limb, heart, and tooth germ (Zhao et al., 1996; Niederreither
682 Sasaki et al. J Dent Res 89(7) 2010
Figure 1. mRNA expression of Adamts4 (A), Aldh1a2 (b), and Lef1 (c) by qRT-PCR. Vertical bars indicate mean ± SD (significant difference: *p < 0.05, **p < 0.01). (A) QRT-PCR confirmed a 3.1-fold reduction in mRNA expression of Adamts4 between E16 and E18 (p < 0.05). (B) QRT-PCR demonstrated a 12.8-fold reduction in expression of
Aldh1a2 mRNA between E16 and E18 (p < 0.01). (C) QRT-PCR
showed a 37.4-fold decrease in expression of Lef1 between E16 and E18 (p < 0.01). Mann-Whitney’s U-test with Bonferroni correction. A, n = 4; B, n = 5; C, n = 5.
table 2. Summary of Down-regulated Genes in Dental Papillae during Odontogenesis
Probe Set ID Accession No. Gene Symbol Between E16 and E18 Between E18 and P3
Calcification according to GO
1455965_at BG064671 Adamts4 14.1
1421299_a_at NM_010703 Lef1 12.8
Development according to GO
1422789_at NM_009022 Aldh1a2 70.8
List of genes showing four-fold change in down-regulation and categorized by gene ontology (GO) as “Calcification”, “Development”, or “Cell Death”. In the “Calcification” category, expression of Adamts4 and Lef1 decreased considerably (fold change > 10.0) between E16 and E18. In the “Development” category, a greater than 70-fold change was recognized in the expression of Aldh1a2 between E16 and E18. However, no genes showing a greater than 10-fold reduction were categorized as “Cell Death”.
et al., 1997; Mic et al., 2002). Aldh1a2 was expressed in part of the dental papillae and dental follicles at E18.5 (Niederreither et al., 1997, 2002). Retinoic acid (RA), a metabolic product of Aldh1a2, and RA receptor have been reported to regulate dif-ferentiation of odontoblasts (Bloch-Zupan et al., 1994; Kronmiller et al., 1994; Berkovitz and Maden, 1995). Furthermore, an excess of RA appeared to reduce alkaline phos-phatase activity and retard growth and differentiation of molar explants (Jones et al., 2008). Our results, together with those of previous reports, suggest that a reduction in the expression of Aldh1a2 between E16 and E18 is involved in regular growth and differentiation from dental papilla cells to odontoblasts.
Lymphoid enhancer-binding factor 1 (Lef1) is a cell-type-specific transcription factor essential in the development of whisker follicles, mammary glands, and tooth germs (van Genderen et al., 1994). In one study in an Lef1-deficient mouse, it was reported that Lef1 regulated expression of Shh, Bmp, Fgf, and Wnt, important factors in odontogenesis, revealing a sig-nificant function for this gene in the determination of tooth morphology. The mRNA expression of Lef1 was reported in both dental epithelium and mesenchyme at E12.5 and E14.5 (Sasaki et al., 2005). Loss of function of Lef1 resulted in
arrested tooth development at the bud stage, demonstrating functional significance for Lef1 during early tooth development (van Genderen et al., 1994). Furthermore, Lef1 directly regu-lated Fgf4 expression, which regulates expression of Fgf3 in dental mesenchyme and Shh in dental epithelium to mediate the critical epithelial mesenchymal interaction (Kratochwil et al., 1996, 2002). However, the expression and localization of Lef1 remain to be clarified in late-stage tooth development. Our results showed that expression of Lef1 was down-regulated and localized to pre-odontoblasts in the cervical loop region at E18, whereas localization was observed at the inner enamel epithelium and dental papilla at E16. This suggests that Lef1 plays an important role in regulating formation of odontoblasts during tooth development, probably via the Fgf pathway, as demon-strated previously (Kratochwil et al., 2002; Sasaki et al., 2005). In summary, we found that 3 genes showed a marked reduction in expression between the pre- and post-natal stages: Adamts4, Aldh1a2, and Lef1. Down-regulation of these genes may represent an important factor in dental papillae during tooth development.
AcKnOWlEDGMEnts
This work was supported by the Japanese Ministry of Education, Culture, Sports, Science and Technology (Grant-in-Aid No. 17591926). We thank Associate Professor Jeremy Williams, Tokyo Dental College (Lab. of Information Support), for his editing of the manuscript. Hodaka Sasaki and Takashi Muramatsu contributed equally to this work.
rEFErEncEs
Berkovitz BK, Maden M (1995). The distribution of cellular retinoic acid-binding protein I (CRABPI) and cellular retinol-acid-binding protein I (CRBPI) during molar tooth development and eruption in the rat.
Connect Tissue Res 32:191-199.
Bloch-Zupan A, Decimo D, Loriot M, Mark MP, Ruch JV (1994). Expression of nuclear retinoic acid receptors during mouse odontogenesis.
Differentiation 57:195-203.
Brown NL, Knott L, Halligan E, Yarram SJ, Mansell JP, Sandy JR (2003). Microarray analysis of murine palatogenesis: temporal expression of genes during normal palate development. Dev Growth Differ 45:153-165. Cho SW, Lee HA, Cai J, Lee MJ, Kim JY, Ohshima H, et al. (2007). The
primary enamel knot determines the position of the first buccal cusp in developing mice molars. Differentiation 75:441-451.
Hikake T, Mori T, Iseki K, Hagino S, Zhang Y, Takagi H, et al. (2003). Comparison of expression patterns between CREB family transcription factor OASIS and proteoglycan core protein genes during murine tooth
development. Anat Embryol
(Berl) 206:373-380.
Hiratsuka K, Kamino Y, Nagata T, Takahashi Y, Asai S, Ishikawa K, et
al. (2002). Microarray analysis of
gene expression changes in aging in mouse submandibular gland. J
Dent Res 81:679-682.
Hurmerinta K, Thesleff I, Saxén L (1980). In vitro inhibition of mouse odontoblast differentiation by vita-min A. Arch Oral Biol 25:385-393. Jernvall J, Thesleff I (2000). Reiterative signaling and patterning during mammalian tooth morphogenesis.
Mech Dev 92:19-29.
Jones DM, Fabian B, Kramer B (2008). The effect of retinoic acid on mouse mandibular molar development in vitro, using alka-line phosphatase as a molecular indicator of differentiation. S Afr
Dent J 63:276, 278-280.
Kollar EJ, Baird GR (1970). Tissue interactions in embryonic mouse tooth germs. I. Reorganization of the dental epithelium during tooth-germ reconstruction. J Embryol
Exp Morphol 24:159-171.
Kratochwil K, Dull M, Farinas I, Galceran J, Grosschedl R (1996). Lef1 expression is activated by BMP-4 and regulates inductive tissue interactions in tooth and hair development. Genes Dev 10:1382-1394.
Kratochwil K, Galceran J, Tontsch S, Roth W, Grosschedl R (2002). FGF4, a direct target of LEF1 and Wnt signaling, can rescue the arrest of tooth organogenesis in Lef1(-/-) mice. Genes Dev 16:3173-3185.
Kronmiller JE, Beeman CS, Kwiecien
K, Rollins T (1994). Effects of the intermediate retinoid metabolite retinal on the pattern of the dental lamina in vitro. Arch Oral Biol 39:839-845. Kuno K, Kanada N, Nakashima E, Fujiki F, Ichimura F, Matsushima K
(1997). Molecular cloning of a gene encoding a new type of metal-loproteinase-disintegrin family protein with thrombospondin motifs as an inflammation associated gene. J Biol Chem 272:556-562. Mic FA, Haselbeck RJ, Cuenca AE, Duester G (2002). Novel retinoic
acid generating activities in the neural tube and heart identified by conditional rescue of Raldh2 null mutant mice. Development 129:2271-2282.
Mina M, Kollar EJ (1987). The induction of odontogenesis in non-dental mesenchyme combined with early murine mandibular arch epithelium.
Arch Oral Biol 32:123-127.
Nakamura H, Fujii Y, Inoki I, Sugimoto K, Tanzawa K, Matsuki H, et al. (2000). Brevican is degraded by matrix metalloproteinases and aggrecanase-1 (ADAMTS4) at different sites. J Biol Chem 275:38885-38890.
Niederreither K, McCaffery P, Drager UC, Chambon P, Dolle P (1997). Restricted expression and retinoic acid-induced downregulation of the retinaldehyde dehydrogenase type 2 (RALDH-2) gene during mouse development. Mech Dev 62:67-78.
Niederreither K, Fraulob V, Garnier JM, Chambon P, Dolle P (2002). Differential expression of retinoic acid-synthesizing (RALDH) enzymes during fetal development and organ differentiation in the mouse. Mech Dev 110:165-171.
Palmer RM, Lumsden AG (1987). Development of periodontal ligament and alveolar bone in homografted recombinations of enamel organs and
papillary, pulpal and follicular mesenchyme in the mouse. Arch Oral
Biol 32:281-289.
Sasaki T, Ito Y, Xu X, Han J, Bringas P Jr, Maeda T, et al. (2005). LEF1 is a critical epithelial survival factor during tooth morphogenesis. Dev
Biol 278:130-143.
Sone S, Nakamura M, Maruya Y, Takahashi I, Mizoguchi I, Mayanagi H,
et al. (2005). Expression of Versican and ADAMTS during rat tooth
eruption. J Mol Histol 36:281-288.
Thesleff I (2003). Epithelial-mesenchymal signalling regulating tooth mor-phogenesis. J Cell Sci 116(Pt 9):1647-1648.
Tortorella M, Pratta M, Liu RQ, Abbaszade I, Ross H, Burn T, et al. (2000). The thrombospondin motif of aggrecanase-1 (ADAMTS-4) is critical for aggrecan substrate recognition and cleavage. J Biol Chem 275:25791-25797.
van Genderen C, Okamura RM, Farinas I, Quo RG, Parslow TG, Bruhn L,
et al. (1994). Development of several organs that require inductive
epithelial-mesenchymal interactions is impaired in LEF-1-deficient mice. Genes Dev 8:2691-2703.
Westling J, Gottschall PE, Thompson VP, Cockburn A, Perides G, Zimmermann DR, et al. (2004). ADAMTS4 (aggrecanase-1) cleaves human brain versican V2 at Glu405-Gln406 to generate glial hyaluro-nate binding protein. Biochem J 377(Pt 3):787-795.
Zhao D, McCaffery P, Ivins KJ, Neve RL, Hogan P, Chin WW, et al. (1996). Molecular identification of a major retinoic-acid-synthesiz-ing enzyme, a retinaldehyde-specific dehydrogenase. Eur J Biochem 240:15-22.
Figure 2. Expression of Adamts4, Aldh1a2, and Lef1 during tooth development (E16, E18, and P3) in mouse tooth germs. In situ hybridization analysis was carried out with frontal sections of E16 (A, D, G), E18 (b, E, H), and P3 (c, F, I) mouse tooth germs. Adamts4 (A-C), Aldh1a2 (D-F), and Lef1 (G-I) showed similar expression patterns during tooth development. At E16, they were localized in inner and outer dental epithelia, stratum intermedium, stellate reticulum, and dental papilla cells adjacent to inner dental epithelium (A, D, G). At E18, localization of Aldh1a2 and Adamts4 was also observed in dental epithelia consisting of inner dental epithelium and stratum intermedium (B, E). However, expression of
Lef1 showed the greatest decrease, and localization was observed in outer dental epithelium of the
cervical area (H). Expression of Aldh1a2 and Adamts4 became weaker or disappeared in odontoblasts differentiated from dental papilla cells and pulp cells at P3, but persisted in ameloblasts differentiated from inner dental epithelial cells (C, F). No expression of Lef1 was observed in odontoblasts, dental pulp cells, or dental epithelium (I). Bar = 100 µm.