IRUCAA@TDC : Four gbpC Gene Homologues in Streptococcus sobrinus
18
0
0
全文
(2) Short Communication Four gbpC gene homologues in Streptococcus sobrinus.. Yutaka Sato1), Ayako Ishikawa1), Kazuko Okamoto-Shibayama1), Kazuko Takada2), and Masatomo Hirasawa2). 1). Department of Biochemistry and Oral Health Science Center, Tokyo Dental College. 2-2, Masago 1-chome, Mihama-ku, Chiba City, 261-8502 Japan 2). Department of Microbiology and Immunology, Nihon University School of Dentistry at Matsudo 2-870-1, Sakaechonishi, Matsudo City, 271-8587 Japan. Corresponding author: Yutaka Sato (E-mail) [email protected] (Phone)81-43-270-3750 (Fax)81-43-270-3752 Runing title: Four gbpC gene homologues in S. sobrinus Field: Microbiology. 1.
(3) Abstract We previously identified the glucan-binding protein C gene, gbpC, solely involved in dextran-dependent aggregation (ddag) of Streptococcus mutans. Recently, we identified two gbpC gene homologues, gbpC and dbl, in Streptococcus sobrinus and suggested that the dbl gene was very likely responsible for ddag of this species. However, homology searches with the gbpC or dbl genes against the incomplete TIGR S. sobrinus ATCC 6715 database suggested that this strain may have other gbpC homologues. PCR-based chromosomal walking from the gbpC and dbl genes revealed two additional homologous genes designated as gbpC2 and dblB. Proteins encoded by these genes exhibited alpha-1,6 glucan-binding activities. Therefore, gbpC2 and dblB are also logical candidates responsible for the ddag phenotype.. Key words Streptococcus sobrinus, glucan-binding protein C, gbpC2 gene, dblB gene. 2.
(4) Introduction Streptococcus mutans and Streptococcus sobrinus are regarded as the two major principal etiologic agents of human dental caries. 1). . Several putative virulence factors involved in dental. caries, e.g. surface protein antigens or glucosyltransfereses (GTFs), are commonly found in both organisms. However, one clear difference in phenotype between these two organisms has been well-known since the 1980’s and is that S. sobrinus exhibits more active dextran (α-1,6 glucan)-dependent aggregation (ddag) than S. mutans. 2). . Therefore, we have used this phenotype as a simple. convenient method to distinguish S. sobrinus from S. mutans as suggested previously. 3). . We initially identified the gbpC gene. 4). involved in ddag of S. mutans. In addition, we recently identified two gbpC gene homologues in clinically isolated S. sobrinus strain 100-4 and suggested that the second homologue gene is very likely involved in ddag of this organism. 5). . Ma et al. proposed to define. glucan-binding proteins as glucan-binding lectins (Gbls) since they would confer the property of the aggregation on bacteria in the presence of exogenously added dextran. 6). . Similar to this suggestion,. and in order to avoid confusion between the two gbpC gene homologues within a species, we have proposed the designation Dbls (Dextran-binding lectins) as referring to a wall-anchored protein family characterized by dextran-binding activity and have designated this second homologue as the dbl gene as described previously. 5). . Both glucan-binding protein C homologues (GbpC and. Dbl) are classified as cell wall-anchored surface proteins in. 3.
(5) Gram-positive bacteria. 7). possessing LPXTG motifs by which sortase. mediates to tether the proteins to cell wall peptidoglycans. However, we could detect neither GbpC nor Dbl proteins in a wall-fraction prepared from S. sobrinus type strain ATCC6715 with Western blot analyses using anti-GbpC or anti-Dbl sera. However, both proteins were detected in concentrated culture supernatants with the same analyses. When homology searches with the gbpC or dbl genes against the incomplete TIGR (The Institute for Genomic Research) S. sobrinus 6715 database were carried out, multiple contigs containing parts of homologous sequence regions with 70-80% identities were detected in each search. These results suggested that strain 6715 may have other cell-associated gbpC homologues involved in ddag of this strain, although the search results at TIGR provided no information concerning chromosomal locations of other gbpC homologues. Therefore, we provisionally carried out PCR-based chromosomal walking of both upstream and downstream regions from the gbpC and dbl genes. Materials and Methods S. sobrinus strain 6715 was maintained and cultured in Todd-Hewitt (TH) broth. Isolation of chromosomal DNA was carried out as previously described. 4). . A genome walking library was. constructed based on nucleotide sequence information obtained from the gbpC and dbl gene regions with the Universal GenomeWalker Kit (BD Biosciences Clontech, Palo Alto, CA) as described previously 8). . Purification of amplified fragments and sequencing analyses were. 4.
(6) carried out as described previously. 5). .. We used BLAST programs to search the S. sobrinus 6715 genome database at TIGR (http://www.tigr.org) for identical nucleotide sequences, and preliminary sequence data obtained from the TIGR website at http://tigrblast.tigr.org/ufmg/index.cgi?database=s_sobrinus%7 Cseq were used to compare with our determined sequences. Gene fragments expressing the extracellular domains of the S. sobrinus GbpC homologues without the signal sequences predicted by the SignalP program 3.0 9) were amplified by PCR and then subcloned into pBAD/His vectors (Invitrogen) as described previously. His-tagged GbpC homologue proteins were overexpressed in E. coli Top10, purified with Ni-Sepharose 6 Fast Flow resin (GE Healthcare Biosciences) and used for glucan-bindig assays as described previously. 5). .. The nucleotide sequence data reported in this communication will appear in the DDBJ, EMBL and GenBank nucleotide sequence databases under accession numbers AB281278, AB281280, AB294108, and AB294109.. Results and Discussion The S. sobrinus 6715 gbpC gene encoded an intact 621 amino acid protein, which was the same size as that in strain 100-4. 5). .. Differences between the two homologous sequences of strain 6715 and 100-4 were detected only at 6 nucleotide positions (99·7% identity) and 2 of these are non-synonymous substitutions, which. 5.
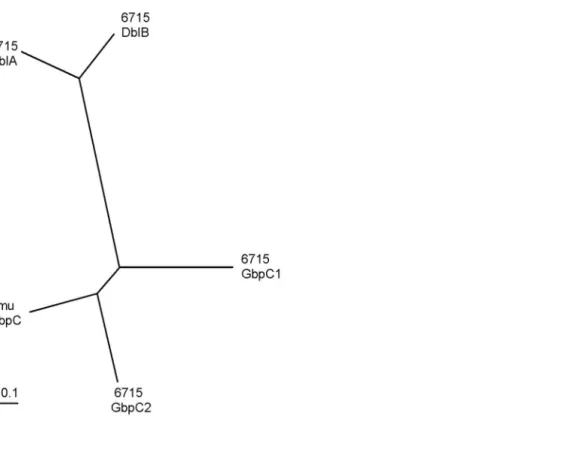
(7) resulted in amino acid substitutions at L94 to S and L427 to H in the strain 6715 sequence. 687 bp downstream from the gbpC termination codon, another open reading frame (ORF) quite similar to the gbpC gene was found on the same strand. The entire sequence of this ORF was determined as described above and was designated as the gbpC2 gene encoding a 632 amino acid protein, which was slightly larger than the GbpC protein. Homology, allowing for gaps between the gbpC and gbpC2 gene sequences, was 54.9%, which was slightly less than that between the gbpC2 and S. mutans gbpC genes (57.2%). In order to avoid confusion between the S. mutans and S. sobrinus gbpC genes, we would like to rename S. sobrinus gbpC gene to gbpC1. The dbl gene sequence in strain 6715 was also determined and encoded an intact 1188-amino acid protein that was 82aa-smaller than the strain 100-4 homologous protein. The dbl gene contains two direct repeats (DR) comprised of the 168nt and 78nt DR-units near the 5’ and 3’ ends, respectively. The numbers of repeating units in both repeats (DR-5’ and DR-3’) in strain 6715 were one unit-smaller than those in strain 100-4. The DR regions and difference of the DR numbers were schematically indicated by the Harr plot analysis (Fig. 1). Both DR-5’ and DR-3’regions were separately amplified from chromosomal DNA of 8 strains of S. sobrinus isolated in our institutions and the amplicons, together with those from strains 6715 and 100-4 as controls, were subjected to agarose gel electrophoretic analysis to estimate their sizes. Only one strain from the 8 isolates exhibited the same sizes of. 6.
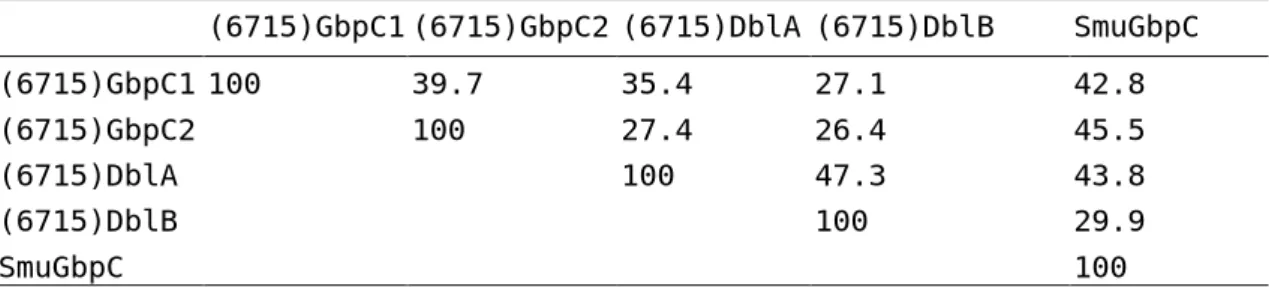
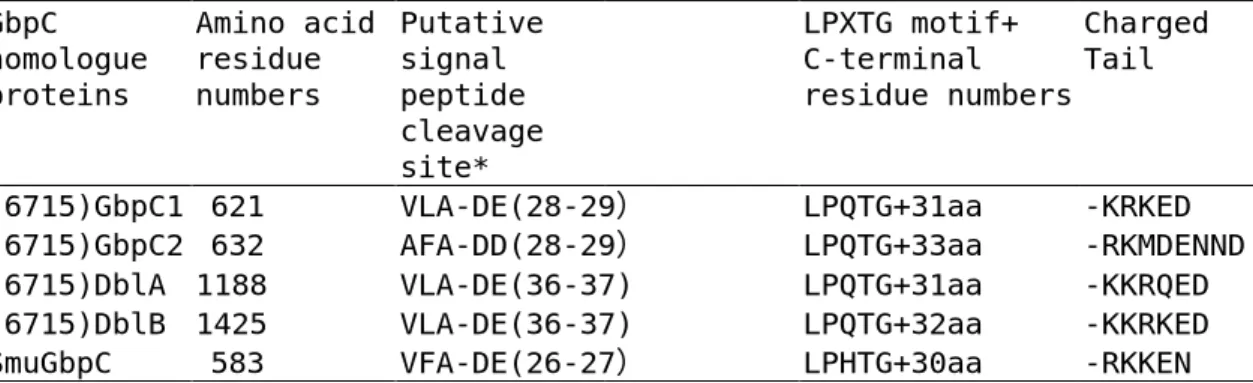
(8) the DR-5’ and DR-3’ amplicons as those amplified from strain 6715 and the other 7 strains appear to contain the same repeating structures as strain 100-4 in both DR-5’ and DR-3’ regions (data not shown). The strain 6715-type dbl gene may not belong to major gene population in S. sobrinus species. Similar to gbpC1 and gbpC2 gene arrangements, another open reading frame (ORF) similar to the dbl gene was found to start at 505bp downstream from the dbl gene termination codon on the same strand. Therefore, we propose that the previously reported dbl gene is renamed dblA and that the downstream ORF is designated as the dblB gene. The dblB gene encoded an 1425 amino acid protein, which was 337aa larger than the DblA protein. Homology, allowing gaps between the dblA and dblB gene sequences, was 55.8%. BLAST searches with these 4 gbpC homologue sequences against the S. sobrinus 6715 genome database at TIGR on the date of June 20, 2007 did not hit any completely identical sequences. Detected homologous sequence regions on the database revealed still 70-80% identities in each search. Pairwise comparisons and phylogenetic tree construction using the four S. sobrinus GbpC homologue proteins with S. mutans GbpC protein are shown in Table 1 and Fig. 2, respectively. These results indicated that the S. sobrinus GbpC2 protein was evolutionally more related to S. mutans GbpC protein than the S. sobrinus GbpC1 protein. Comparisons of other features of wall-anchored proteins are summarized in Table 2. These features are well conserved among these. 7.
(9) proteins. Purified recombinant proteins corresponding to the extracellular domains of these S. sobrinus GbpC homologue proteins were subjected to alpha-1,6 glucan-binding assays as described in Materials and Methods. All of the GbpC2, DblA, and DblB recombinant proteins derived from strain 6715 exhibited alpha-1,6 glucan-binding activities (Fig. 3), in contrast to the S. sobrinus GbpC (GbpC1) protein defective in such activity as recently reported 5). . We have identified the GbpC protein solely involved in ddag. of S. mutans 4), although three other proteins designated as GbpA, GbpB, and GbpD. 10). exhibiting glucan-binding activity are encoded. by the genome of this organism. In addition, the genome project of this organism revealed the absence of other gbpC gene homologues 11). . The S. sobrinus genome contained at least four gbpC gene. homologues presumed to be duplicated from their antecedent genes in contrast to S. mutans genome. Epidemiological studies have indicated that S. sobrinus is more frequently isolated from highly caries-susceptible patients compared to S. mutans 12). The cariogenic potential in rats of S. sobrinus is also greater than that of S. mutans. 13). . These results may be related to the genome structure. of this organism, which is highly adapted to sucrose-rich environments where multiple glycosidic linkage types of glucans are abundantly synthesized by mutans streptococci or other streptococci. Proteins encoded by duplicated genes have often expanded a repertoire of functions. S. mutans GtfB/C/D gordonii SspA/B. 15). 14). and S.. proteins may be such examples. The relative. 8.
(10) affinities of the three glucan-binding proteins for alpha-1,6 glucan were slightly different. The GbpC2 protein appeared to exhibit higher affinity in the lower glucan concentration range than the other two proteins and the GbpC (GbpC1) protein exhibited no alpha-1,6 glucan-binding activity but bound Sephadex resin as reported recently. 5). . Each protein may bind a preferable type of. glucans. We could detect more Dbl (DblA) protein from the strain 100-4 cell wall but detected neither GbpC (GbpC1) nor Dbl (DblA) proteins in a wall-fraction prepared from strain 6715 with Western blot analyses using anti-GbpC (GbpC1) or anti-Dbl (DblA) sera. Expression or the repertoire of each protein may be different from strain to strain. However, more precise kinetic analyses will be necessary to clarify this hypothesis and such works are in progress in our laboratory. We suggested that the dbl (dblA) gene in S. sobrinus, but not the gbpC (gbpC1), is a logical candidate responsible for the ddag phenotype in a recent report. 5). . However, GbpC2 and DblB also. exhibited alpha-1,6 glucan-binding activities. Therefore, the gbpc2 and dblB genes are also logical candidates contributing to the ddag phenotype in strain 6715. To resolve this question, it would be ideal to introduce gene-derected mutagenesis in strains of S. sobrinus.. 9.
(11) Acknowledgments Preliminary sequence data was obtained from The Institute for Genomic Research website at http://www.tigr.org. This investigation was supported in part by Grants-in-Aid for Scientific Research (17591925) from the Research Fund of MEXT, and in part by a Grant from The Tsuchiya Foundation for Culture and Science to Y.S. 10.
(12) Fig. 1. The Harr plot analysis with the strains 100-4 and 6715 dblA genes. Unit size to compare was 10 nt and dot plot matching number was 8 nt.. 11.
(13) Fig. 2. Phylogenetic relationship among the S. sobrinus GbpC protein homologue sequences and S. mutans GbpC sequence. Distances were calculated by the neighbor-joining (NJ) method using the DDBJ ClustalW program and the tree was drawn with the TreeView program. 16). using the calculated result.. 12.
(14) Fig. 3. Biotin-dextran binding by histidine-tagged derivatives of the S. sobrinus GbpC1, GbpC2, DblA, and DblB proteins. GbpC1, GbpC2, DblA, and DblB proteins were purified as described in the text. BSA was used as a control. Higher A490 values represent higher binding activities. Wells of the HisSorb plates were coated with approximately 9 µg of protein. Each well was reacted with the indicated concentrations of biotin-dextran, washed, and detected as described previously. Data presented are averages from three independent determinations for each sample.. 13.
(15) Table 1. Pairwise comparisons among the four S. sobrinus GbpC homologue proteins with the S. mutans GbpC protein (6715)GbpC1 (6715)GbpC2 (6715)DblA (6715)DblB. SmuGbpC. (6715)GbpC1 100. 39.7. 35.4. 27.1. 42.8. (6715)GbpC2 (6715)DblA (6715)DblB. 100. 27.4 100. 26.4 47.3 100. 45.5 43.8 29.9. SmuGbpC. 100. 14.
(16) Table 2. Comparison of features for wall-anchored proteins among the four S. sobrinus GbpC homologue proteins with the S. mutans GbpC protein GbpC homologue proteins. (6715)GbpC1 (6715)GbpC2 (6715)DblA (6715)DblB SmuGbpC. Amino acid Putative residue signal numbers peptide cleavage site* 621 VLA-DE(28-29) 632 AFA-DD(28-29) 1188 VLA-DE(36-37) 1425 VLA-DE(36-37) 583 VFA-DE(26-27). Charged LPXTG motif+ Tail C-terminal residue numbers. LPQTG+31aa LPQTG+33aa LPQTG+31aa LPQTG+32aa LPHTG+30aa. *Predicted by the hidden Markov Model at SignalP website (http://www.cbs.dtu.dk/services/SignalP/).. 15. -KRKED -RKMDENND -KKRQED -KKRKED -RKKEN.
(17) References 1. 2.. 3.. 4.. 5.. 6.. 7.. 8.. 9.. 10.. 11.. Loesche, W.J.: Role of Streptococcus mutans in human dental caries. Microbiol Rev 50: 353-380, 1986. Drake, D., Taylor, K.G., Bleiweis, A.S. and Doyle, R.J.: Specificity of the glucan-binding lectin of Streptococcus cricetus. Infect Immun 56: 1864-1872., 1988. Wu-Yuan, C.D. and Gill, R.E.: An 87-kilodalton glucan-binding protein of Streptococcus sobrinus B13. Infect Immun 60: 5291-5293, 1992. Sato, Y., Yamamoto, Y. and Kizaki, H.: Cloning and sequence analysis of the gbpC gene encoding a novel glucan-binding protein of Streptococcus mutans. Infect Immun 65: 668-675, 1997. Kagami, A., Okamoto-Shibayama, K., Yamamoto, Y., Sato, Y. and Kizaki, H.: One of two gbpC gene homologues is involved in dextran-dependent aggregation of Streptococcus sobrinus. Oral Microbiol Immunol in press: 2007. Ma, Y.S., Lassiter, M.O., Banas, J.A., Galperin, M.Y., Taylor, K.G. and Doyle, R.J.: Multiple glucan-binding proteins of Streptococcus sobrinus. J Bacteriol 178: 1572-1577, 1996. Navarre, W.W. and Schneewind, O.: Surface proteins of gram-positive bacteria and mechanisms of their targeting to the cell wall envelope. Microbiol Mol Biol Rev 63: 174-229., 1999. Okamoto-Shibayama, K., Sato, Y., Yamamoto, Y., Ohta, K. and Kizaki, H.: Identification of a glucan-binding protein C gene homologue in Streptococcus macacae. Oral Microbiol Immunol 21: 32-41, 2006. Nielsen, H., Engelbrecht, J., Brunak, S. and von Heijne, G.: Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Protein Eng 10: 1-6., 1997. Banas, J.A., Russell, R.R. and Ferretti, J.J.: Sequence analysis of the gene for the glucan-binding protein of Streptococcus mutans Ingbritt. Infect Immun 58: 667-673., 1990. Ajdic, D., McShan, W.M., McLaughlin, R.E., Savic, G., Chang, J., Carson, M.B., Primeaux, C., Tian, R.Y., Kenton, S., Jia, H.G., Lin, S.P., Qian, Y.D., Li, S.L., Zhu, H., Najar, F.,. 16.
(18) 12.. 13.. 14.. 15.. 16.. Lai, H.S., White, J., Roe, B.A. and Ferretti, J.J.: Genome sequence of Streptococcus mutans UA159, a cariogenic dental pathogen. Proc. Natl. Acad. Sci. USA 99: 14434-14439, 2002. Hirose, H., Hirose, K., Isogai, E., Miura, H. and Ueda, I.: Close association between Streptococcus sobrinus in the saliva of young children and smooth-surface caries increment. Caries Res 27: 292-297, 1993. de Soet, J.J., van Loveren, C., Lammens, A.J., Pavicic, M.J., Homburg, C.H., ten Cate, J.M. and de Graaff, J.: Differences in cariogenicity between fresh isolates of Streptococcus sobrinus and Streptococcus mutans. Caries Res 25: 116-122, 1991. Ooshima, T., Matsumura, M., Hoshino, T., Kawabata, S., Sobue, S. and Fujiwara, T.: Contributions of three glycosyltransferases to sucrose-dependent adherence of Streptococcus mutans. J Dent Res 80: 1672-1677, 2001. Egland, P.G., Du, L.D. and Kolenbrander, P.E.: Identification of independent Streptococcus gordonii SspA and SspB functions in coaggregation with Actinomyces naeslundii. Infect Immun 69: 7512-7516, 2001. Page, R.D.M.: TREEVIEW: An application to display phylogenetic trees on personal computers. Computer Applications in the Biosciences 12: 357-358, 1996.. 17.
(19)
図


+2
関連したドキュメント
FIGURE 1: The poly-T region in intron 8 of the cystic fibrosis transmembrane conductance regulator (CFTR) gene amplified by polymerase chain reaction and analysed by a direct
Octn1 gene knockout mice (octn1 −/− ) have been constructed and exhibit a marked reduction in small- intestinal absorption, tissue distribution in various organs, and
ABUNDANT EXPRESSION OF NUCLEOSOME ASSEMBLY PROTEIN 1 (NAP1) GENE IN GOLDFISH SCALE WITH LATERAL LINE
ABUNDANT EXPRESSION OF NUCLEOSOME ASSEMBLY PROTEIN 1 (NAP1) GENE IN GOLDFISH SCALE WITH LATERAL LINE.. LiHua LI 1 , Hironobu KATSUYAMA 2 , Son Ngoc DO 1 , Masayuki SAITO 1 ,
Transporter adaptor protein PDZK1 regulates several influx transporters (PEPT1 and OCTN2) in small intestine, and their expression on the apical membrane is diminished in pdzk1
In [1, 2, 17], following the same strategy of [12], the authors showed a direct Carleman estimate for the backward adjoint system of the population model (1.1) and deduced its
Comparing the Gauss-Jordan-based algorithm and the algorithm presented in [5], which is based on the LU factorization of the Laplacian matrix, we note that despite the fact that
A., Some application of sample Analogue to the probability integral transformation and coverages property, American statiscien 30 (1976), 78–85.. Mendenhall W., Introduction
“Breuil-M´ezard conjecture and modularity lifting for potentially semistable deformations after
