*Corresponding author: e-mail genkai@kochi-u.ac.jp
Regime Shifts in Response to Human Impacts and Management
in Ecosystems
Motomi Genkai-Kato
*Graduate School of Kuroshio Science, Kochi University, (Kochi, Japan) Abstract
Ecosystems may have more than one state with a self-stabilizing mechanism, so that a shift between states does not occur frequently and is not readily reversible. These big changes of ecosystem states are termed regime shifts. Regime shifts are difficult to predict and are problematic for ecosystem managers because of irreversibility. Here I briefly introduce a theoretical approach to predict the shift between a clear-water state and a turbid state in lakes, the best-studied example of regime shifts. This paper also discusses regime shifts in other systems besides ecosystems.
Key words: ecosystem, human impact, management, mathematical model, regime shift
1. Introduction
Regime shifts are increasingly reported in natural environments (Scheffer et al., 2001). Increased evidence of regime shifts seems to be related to the enhanced human pressure in ecosystems (Millennium Ecosystem Assessment, 2005). Often regime shifts are not revers-ible, or can be reversed only slowly, with long time delays (Carpenter, 2003). The irreversibility and time lags are related to the phenomenon called "hysteresis." This phenomenon was first identified in magnetic mate-rials by Sir James Alfred Ewing (Ewing, 1891). Because of the property that systems react slowly or do not return completely to their original state, hysteresis is problem-atic for people who depend on ecosystems, that is to say all of us.
In lake ecosystems, for example, nutrient loading from surrounding watersheds does not necessarily lead to turbid water or eutrophication, because the effect of nutrient loading on water clarity also depends on changes in biotic communities. Algal blooms are due to changes in biotic communities and may act as a buffer against alterations of ecosystems by humans, but may cause hysteretic phenomena (i.e., regime shifts) in nature. Thus ecosystem managers who neglect the indirect effects of biotic communities could fail at ecosystem restoration.
The nutrient loading and water clarity of a lake can be illustrated as a spring and weight diagram where the
mass of the weight represents the amount of nutrient loading (Fig. 1a). The more mass, the more the spring stretches downward and the more turbid the lake water. The resilience of the spring to the weight corresponds to the response of biotic communities to nutrient loading. Managers who consider biotic communities, while reducing human impacts on ecosystems, may restore water quality without the loss of biotic integrity (Fig. 1b). On the other hand, managers who ignore the effects of biotic communities and instead adopt technological methods, such as dredging and the application of chem-ical agents, may restore water quality but irreversibly alter the natural state of biotic communities. In such cases, biotic communities cannot react in response to reduced nutrient loading, becoming something like a dead spring (Fig. 1c).
This paper presents the use of ecological theories as an application tool for ecosystem management. As an example, I describe a mathematical model to predict the threshold values of drivers such as nutrient loading in lakes. In doing so, I show how mathematical models are powerful tools to prevent ecological surprises such as regime shifts in ecosystem management. While the paper starts with a case study on the prediction of the possibility of regime shifts in lakes, I also show some examples of regime shifts in other systems to draw more attention to the research currently being performed on regime shifts.
2. A Model for Prediction of Regime Shifts in a Lake Ecosystem
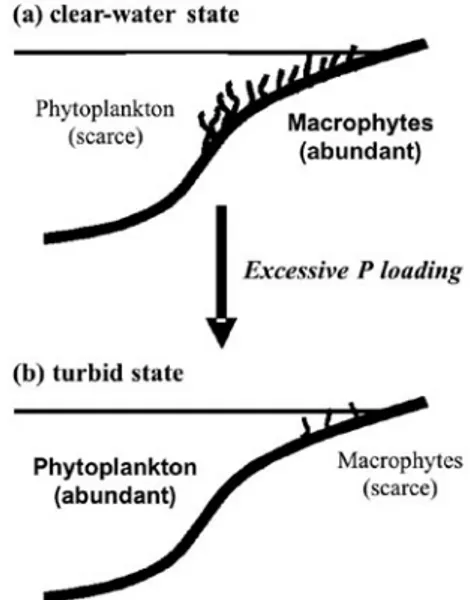
Lakes may have two contrasting states in water clarity due to excessive phosphorus (P) input: a water state vs. a turbid one (Scheffer, 1998). The clear-water state is often characterized by dense macrophytes in the littoral area and the turbid one by algal blooms in the pelagic area (Timms and Moss, 1984). Macrophytes are suggested to reduce P recycling from lake bottoms to the water column by stabilizing sediments; and pro-vide refuge for zooplankton against fish predation, thus increasing grazing pressure on phytoplankton (Scheffer et
al., 1993). As long as a lake is clear, the clear-water state
is self-maintained by these feedback mechanisms (Fig. 2a). On the other hand, once phytoplankton are dominant due to excessive P input, rooted macrophytes become sparse because of shading by dense phytoplankton. In this case, the nutrient resource for phytoplankton growth is recycled from sediments and grazing pressure from zooplankton is diminished, self-maintaining the turbid state (Fig. 2b). Thus the interaction between pelagic and littoral areas through competition between phytoplankton and macrophytes is crucial for regime shifts, and the vul-nerability of lakes to regime shifts as well as lake water quality are both likely to depend on lake morphometry such as lake area and lake depth.
Here I follow a theoretical approach by Genkai-Kato and Carpenter (2005) where a mathematical model was used to predict the possibility of regime shifts in relation to basic limnological characteristics such as the area and mean depth of lakes, and water temperature. The frame of the model is composed of the dynamics of algal density and phosphorus concentration (Fig. 3; see Genkai-Kato and Carpenter (2005) for the model and results in detail). The algal density in the model repre-sents the mean chlorophyll concentration in the epilim-nion in summer. We incorporated into the frame model the relationships between lake area and epilimnion depth, between chlorophyll concentration and the maximum distribution depth of macrophytes, between hypolim-nion thickness, temperature at hypolimhypolim-nion, chlorophyll concentration, and the P recycling rate from sediments. The role of macrophytes was embedded in the model by preventing P recycling from sediments, and thus P recycling was assumed to occur from sediments where macrophytes were absent (i.e., pelagic area). The shape of the lake basin was modeled by quadric surfaces.
There were three types of lake response to eutro-phication: "reversible," eutrophication is gradual and recovery is proportional to the reduction in P input; "hysteretic," eutrophication involves a regime shift and recovery requires severe reductions in P input for a period of time; "irreversible," eutrophication involves a regime shifts and recovery is impossible by means of
Fig. 1. Weight-and-spring model for eutrophication in a lake.
The weight and its position stand for the amount of nutrient loading and the resultant water quality, respectively. A bigger weight represents heavier nutrient loading in the lake. The spring represents biotic communities in the lake which can play a role in mitigating the effect of nutrient loading on lake eutro-phication. (a) A heavy weight results in its lower position, indi-cating turbid water. (b) Restoration of clear water by means of a management policy featuring a reduction in nutrient loading. (c) Restoration by management policies using technological methods (scissors correspond to, for example, the application of chemical agents) that have the potential to impair the biotic communities in the lake.
Fig. 2. Two alternative states in water quality in a lake. (a) A clear-water state is characterized by abundant macro-phytes in the littoral area and scarce phytoplankton. (b) A turbid state is characterized by phytoplankton blooms in the pelagic area and scarce macrophytes. Excessive phosphorus loading of a lake is suggested to cause a shift from a clear-water state (a) to a turbid state (b).
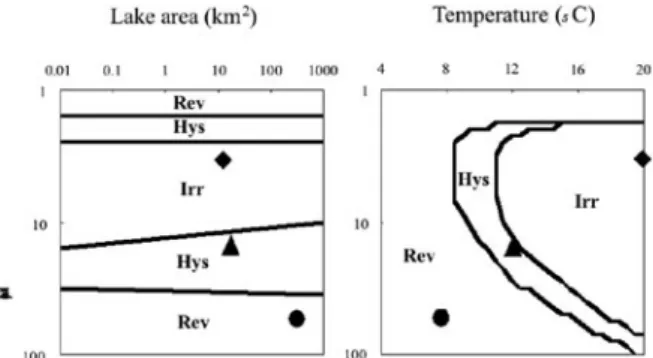
reductions in P input alone. The model predicted that lakes with intermediate mean depths and higher tempera-tures at hypolimnion would be most vulnerable to regime shifts and restoration could not be accomplished in such lakes (Fig. 4). Macrophytes were effective in the reduc-tion of P recycling from sediments in shallow lakes; while the volume of water contained within the hypolim-nion of large, deep lakes greatly diluted nutrients coming in from the watershed. Thus, the model predicted that lakes with intermediate mean depths and higher hypolim-nion temperatures would be most vulnerable to regime shifts making restoration very difficult in such lakes (Fig. 4). The model clarified the existence of peculiar depths in an intermediate range where neither the macrophyte nor dilution effect occurred. Notably, lake area was not a helpful predictor of the possibility of regime shifts or restoration.
The model showed that the vulnerability of lakes to regime shifts can be categorized by mean depth and tem-perature at hypolimnion. Here let me give an example for each type (Fig. 4). Lake Biwa in central Japan (north basin, mean depth: 43 m, temperature at hypolimnion: 7°C, lake area: 613 km2) is categorized as reversible,
Lake Mendota in Wisconsin, USA (12.7 m, 12°C, 40 km2) as hysteretic, and Lake Suwa in central Japan (4.7
m, 20°C, 13.3 km2) as irreversible. As the region of the
hysteretic type is small, most lakes in the world could be categorized as either reversible or irreversible.
The existence of lakes vulnerable to regime shifts at intermediate mean depths is worthy of note. The value
of a lake in terms of its recreational utility for uses such as fishing and the value of its drinking water are consid-ered to increase with its size. By contrast, the number of lakes decreases in general with lake size. Thus lakes of intermediate sizes may be most valuable for us in terms of easy accessibility and considerable utility. The model results suggest that more cautious management may be necessary for such lakes, rather than for other shallow lakes or deep lakes which are naturally protected by the macrophyte or dilution effects respectively.
3. Regime Shifts as Phenomena Happening at Hand
In ecosystems the term "regime shift" was origi-nally used for fisheries in oceanography (Francis and Hare, 1994). Hare and Mantua (2000) showed that the physical and biological environments in the North Pacific shifted remarkably several times in the past. The study of physical environments included mostly climatic char-acteristics and the biological studies focused on biomass, recruitments and catches of oceanic species ranging from zooplankton to salmon. Scheffer et al. (2001) reported other examples of regime shifts: coral reefs (state with corals vs. state with fleshy macroalgae), woodlands (woodlands vs. open landscape with grasses), and deserts (vegetation cover vs. arid desert). In addition to these, river ecosystems may have abrupt transitions in river channel (wide, shallow valleys with converted flow vs. entrenched arroyos with divergent flow) associated with vegetation (Dent et al., 2002).
Brock (2006) describes examples of multistable
Fig. 3. Basic model for predicting sudden eutrophication in lakes due to phosphorus recycling in relation to lake mor-phometry, temperature and macrophytes (after Genkai-Kato and Carpenter (2005)).
Fig. 4. Prediction of lake response types to eutrophica-tion by the model shown in Fig. 3 (after Genkai-Kato and Carpenter (2005)).
In the left panel (lake area vs. mean depth), water temperature is set at 12°C. In the right panel (water temperature vs. mean depth), lake area is set at 40 km2. Rev, reversible type; Hys,
hysteretic type; Irr, irreversible type. An example of each type of lake response was also indicated: ●, North Basin of Lake Biwa (Shiga, Japan); ▲, Lake Mendota (Wisconsin, USA); ◆, Lake Suwa (Nagano, Japan).
economic and social systems. Economies display two contrasting phases, prosperity and depression, and they fluctuate between these phases. These fluctuations are often referred to as economic cycles or business cycles. Although the fluctuations are called cycles, they are dif-ferent from regular cycles such as seasonal fluctuations in temperature; in that they do not necessarily repeat at regular time intervals or are not mechanical in their regu-larity. In the state of depression, for example, an excess of goods supply leads to (1) a decrease in the average price level, (2) a decrease in the producers' interests, (3) a reduction in incomes, and (4) a reduction in the consumers' purchasing power. These negative feedback effects often set off the so-called deflationary spiral. In fact unemployment displays a hysteretic phenomenon (Blanchard and Summers, 1987). As unemployment is closely related to negative economic feedback previously mentioned, unemployment tends to last for a long time and the recovery of employment is difficult.
Human health has also two alternative states: healthy and sick conditions. Sickness such as cold or fever usually results from lack of sleep, malnutrition, or overwork. Once a person gets sick, recovery is not immediately accomplished even if he/she takes sufficient rest and nutrition. The problem of human obesity may be viewed as a regime shift. Human weight is dependent upon the balance between the magnitude of exercise and dietary calories gained from foods. As being overweight generally makes it more difficult to exercise (e.g., the load on the knees increases with weight) and reduced calorie intake sometimes results in a reduction in human metabolic rate, recovery from obesity is hard to accom-plish healthily (i.e., without rebounding). Also, the gradual reduction of the metabolic rate as humans age is comparable to slow ecosystem variables, such as nutrient input and global warming, which have the potential to trigger regime shifts.
Regime shifts are increasingly being reported on in the last few decades. These shifts, whether ecological, economic or health-related, could be caused by the human pursuit of efficiency in agriculture and industrial production. It is likely that in many cases human altera-tions to ecosystems change the natural self-stabilizing mechanisms inherent in ecosystems such as those that resist daily and seasonal changes in temperature and light conditions. The key variables for regime shifts, such as global warming and nutrient loading, originate from emissions of CO2 and polluted waters from industrial
and agricultural activities. Automobiles and computers have greatly improved efficiency in human activities; on the other hand, they have led to a decrease in human
physical activity. Fast food allows more time for produc-tive activities with less time spent cooking, but tends to contain high amounts of fat and is therefore high in ener-getic calories. On the other hand, fast food is often poor in nutrition and leads to malnutrition problems, which, in conjunction with reduced exercise, causes the current obesity epidemic seen in many areas of the globe.
We realize the preciousness of health when we lose it. Health is a problem for each individual and treat-ment for sickness has been well established due to the dramatic progress in medical science. On the other hand, ecosystem regime shifts are infrequent compared to the number of medical patients, but have the potential to cause extensive damage to a considerable number of people just as economic depressions do. There is a need for extensive research on ecosystem regime shifts using multiple approaches such as theory, observation, and experiments in order to prevent damage to human well-being across society, while also trying not to make the same mistakes with human health on an individual level. As humans continue to impact nature, regime shifts in ecosystems are not just ecological, but economic problems the solutions to which lie at the crossroads of ecology and public management (Carpenter, 2003).
References
Blanchard, O., Summers, L. 1987. Hysteresis in unem-ployment. Eur. Econ. Rev., 31, 288-295.
Brock, W. A. 2006. Tipping points, abrupt opinion changes, and punctuated policy change. In: Repetto, R. (ed.) Punctuated equilibrium and the dynamics of U.S. environmental policy. Yale University Press, New Haven, pp. 47-77.
Carpenter, S. R. 2003. Regime shifts in lake ecosys-tems: pattern and variation. Volume 15 in the Excellence in Ecology Series. Ecology Institute, Oldendorf/Luhe.
Dent, C. L., Cumming, G. S., Carpenter, S. R. 2002. Multiple states in river and lake ecosystems. Philos. T. Roy. Soc. B, 357, 635-645.
Ewing, J. A. 1891. Magnetic induction in iron and other materials. The Electrician Printing and Publishing, London.
Francis, R. C., Hare, S. R. 1994. Decadal-scale regime shifts in the large marine ecosystems of the North-east Pacific: a case for historical science. Fish. Oceanogr., 3, 279-291.
Genkai-Kato, M., Carpenter, S. R. 2005. Eutrophication due to phosphorus recycling in relation to lake morphometry, temperature, and macrophytes.
Ecology, 86, 210-219.
Hare, S. R., Mantua, N. J. 2000. Empirical evidence for North Pacific regime shifts in 1977 and 1989. Prog. Oceanogr., 47, 103-145.
Millennium Ecosystem Assessment. 2005. Ecosystems and human well-being: our human planet. Island Press, Washington DC.
Scheffer, M., Hosper, S. H., Meijer, M. L., Moss, B., Jeppesen, E. 1993. Alternative equilibria in shallow lakes. Trends Ecol. Evol., 8, 275-279.
Scheffer, M. 1998. Ecology of shallow lakes. Chapman & Hall, New York.
Scheffer, M., Carpenter, S., Foley, J. A., Folke, C., Walker, B. 2001. Catastrophic shifts in ecosys-tems. Nature, 413, 591-596.
Timms, R.M., Moss, B. 1984. Prevention of growth of potentially dense phytoplankton populations by zooplankton grazing, in the presence of zooplank-tivorous fish, in a shallow wetland ecosystem. Limnol. Oceanogr., 29, 472-486.