身体・神経系の相互作用による原初的運動創発の構成論
○ 山田 康智 (東京大学) 西川 鋭 (東京大学) 志田 和也 (東京大学)
新山 龍馬 (MIT) 國吉 康夫 (東京大学)
1. はじめに
事前に想定できない状況を含む実環境において,柔 軟に動き回ることのできる能力は実世界知能の基盤で ある.この運動スキルは,神経系が身体ダイナミクス を活かすと同時に,身体ダイナミクスが感覚情報を通 じ神経系のダイナミクスを構造化する連続的な身体・ 神経系の相互作用によって可能になる[1][2].
こうした身体・神経系の相互作用に基づき,生物の運 動スキルの理解を試みる研究は,動的に形成される機能 的な集団であるMotor synergy や,身体との相互引き こみ現象により運動を実現する内在性のリズム生成機 構である神経振動子を中心に展開されてきた[3][4][5]. しかし,これらの神経系レベルでのメカニズム,及びそ の形成原理に関してはほとんど知見が得られていない.
一方で,発達研究により胎児期から始まる自発的運 動が運動認知発達において重要な役割を果たす知見が 集まっている[6].ヒトの胎児・乳児においては,これ らの自発運動は多様な運動を包含し,そこから特定の 運動が分化していく形で運動発達が実現されていくと いう事が示唆されている[7].更に,これらの全身の自 発運動の発現メカニズムとして神経振動子が重要視さ れている[8].以上の知見を踏まえ,身体・神経系の相 互作用が潜在的に有する多様なダイナミクスの探索と して初期の自発運動を捉える.この探索的運動を生物 の適応的な運動スキルの原初と考え,本研究では「原 初的運動」と呼ぶ事にする.
本研究の目的は,この生物の「原初的運動」の形成 の神経系メカニズムについての構成論的な理解と,そ の工学的実現である.具体的には以下の手順で行う.
• 生物学的妥当な身体と神経系を有すシステムの構築
• 環境との相互作用を通じた振舞いの観察及び解析
• 構成要素の一部を変更し,相互作用による振舞い の比較
以上の構成論的アプローチを通して,「原初的運動」の 形成メカニズムを説明,再現可能な原理の抽出及び,自 律的な運動探索を実現するロボットシステムの実現を 目指す.
2. 四脚筋骨格ロボットシステム
感覚運動ループの構造を左右する身体・神経系の各 要素について生物学的妥当性を有する四脚ロボットを 開発した(図 1, 表 1).具体的には以下の四つの特徴か ら成る.
• 運動器:生体筋の特性を有する人工筋
• 形態:機能解剖学的知見を基にした筋骨格系
• 感覚器:生体筋を模した感覚フィードバック
• 神経系:神経科学的知見に基づく脊髄延髄系モデル
図1四脚筋骨格ロボット.
図2筋配置.下線付きは二関節筋.
生物の運動実現における身体と神経系の相互引きこ み現象において,重要な役割を担う運動器の粘弾性を, McKibben 型空気圧人工筋を用い再現した [9][10].空 気圧人工筋は圧力比例制御弁により連続的に制御を行 い,空気は外部に取り付けたコンプレッサにより供給 する.
運動器の配置に関しては,生物一般に多く存在する 多関節筋を考慮し,四脚動物の筋配置を基にした筋骨 格系として実現した [11] (図 2).
感覚フィードバックに関しては,生体筋の筋紡錘と ゴルジ腱器官を模し,各筋の長さと張力情報をフィー ドバックとして用いた.各筋の長さは,各関節に配置 されたポテンショメータから幾何的に計算した.張力 F [N] に関しては,得られた筋の長さ lm[m],及び各 筋の圧力センサp[Pa] の値から以下の理論式により求 めた[12][13].
F = p (
A(1 − lm Lmax
)2− B )
, (1) where A= 3
4πD
2
0cot2θ0, B= 1 4πD
2
0cosec2θ0.
ここで, Lmax [m] は筋の最大長, D0 [m] はラバー チューブの初期直径,そしてθ0は軸方向に対する繊維 の初期角度である.ここでは,以下の値を用いたD0= 0.008,θ0= 16.
表1ロボットの仕様. actuators 10 artificial pneumatic
muscles (McKibben type) passive elements 10 springs
(Misumi, AWY12-70) valve 10 proportional pressure
valves (Hoerbiger, tecno basic) pressure sensor Fujikura,
XFGM-6001MPGSR potentiometer Murata, PVS1A103A01
CPU board General Robotix, Lepracaun CPU board OS linux-2.6.21.1-ARTLinux mass of the robot 4.0 kg
materials ABS resin, CFRP pipes and aluminum boards length of body 350 mm
length of each part shoulder - elbow 140 mm
elbow - forefoot 180 mm hip - knee 150 mm knee - ankle 150 mm ankle - hind foot 105 mm
図3脊髄延髄系モデル
神経系モデルとしては,生物学的知見に基づいた脊 髄延髄系モデルを用いた[14] (図 3).このモデルは,伸 長反射以外は組み込まず,各筋で独立しており,神経 系レベルで筋間の結合を一切持たない.つまり,明示 的な運動表現や筋間の関係性を決める要素を一切含ま ない.しかし,振動子モデル及び身体の非線形性によ る相互引きこみ現象は,各筋に身体ダイナミクスを介 した相互作用を可能にし,その結果として多様な協調 運動の創発が実現される事が,シミュレーションによ り示されている[14].モデルは,一つの筋,α,γ 運動 ニューロン,脊髄内介在性感覚ニューロンS0,そして 神経振動子モデルから構成される.すべてのモジュー ルは,生物学的知見を考慮した遅れとゲインが設定さ れている.これらの値はKuniyoshi et al. によるもの を用いた[14].
3. 複数の協調的な運動実現
開発したロボットシステムを用い,身体・神経系の 動的な相互作用を通した振る舞いの観察及び解析を目 的とした実験を行った.すべての実験において,初期
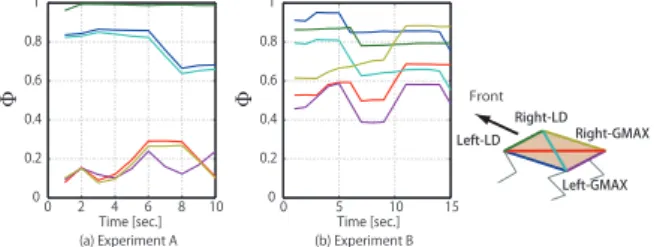
図5ロボット実験における位相同期率の時間変化.
図6 ロボット実験における筋の感覚から運動への情報 の流れ.
姿勢及び神経系モデルのパラメータは一定とした.ま た,神経系のパラメータはランダムで初期化を行った.
実験結果として,実現された運動パターンの中には, 前進してから後退し,再び前進する運動,前進/後退の みの運動,跳躍様運動,地団太するような運動パター ンの創発が観察された(図 4)1 .
運動パターンの協調関係を調べるため,各筋の運動 指令の位相同期率の解析を行った [15].時系列データ x(t) に対して,瞬時位相 φ を以下の式で求める.
φ= arctanxH(t)
x(t) . (2) ここで,xH(t) は Hilbert 変換である.この瞬時位相の 二つの変数間の位相差∆φ(t) を用いて位相同期率は以 下の式で定義される.
Φ =
√
<cos ∆φ(t) >2T + < sin ∆φ(t) >2T. (3)
ここで,<· >TはT ステップ間の時間平均である.位 相同期率は0 から 1 の間の範囲をとり,完全に位相同 期した時には1 を示す.その結果,筋間における高い 位相同期率と動的な協調関係の変化が観察された (図 5).
更に,このカップリングが,神経系モデルにおいて 独立な筋間に,身体ダイナミクスを介した相互作用が 生じた結果であるという仮説を検証するため,筋間の 情報の流れを解析した.各筋の感覚情報から運動情報
1動 画 は 以 下 を 参 照 .http://www.isi.imi.i.u-tokyo.ac. jp/~y-yamada/pigorass_with_spinobulbar_model_in_april. html
(a) Experiment A: 前進から後退,再び前進した運動. (b) Experiment B: 後退の みの運動.
図4ロボット実験のスナップショット.
への情報の流れを,時系列間の因果構造を捉える事が 可能なTransfer entropy を用いて解析した [16].二つ の時系列X,Y に対して以下の式により計算される.
TX→Y = ∑
yt,yt(l)
−1,x (k) t−1
p(yt, y(l)t−1, x(k)t−1)×
log2p(yt|y
(l) t−1, x
(k) t−1)
p(yt|yt−1(l) ) . (4) そして,p(·|·) は条件付き確率,k,l は埋め込み次元, そしてTX→Y はX から Y への因果の度合いを示して いる.解析の結果,神経系モデルでは独立な筋間にお いて情報の流れが生じている事がわかった(図 6).
4. 運動創発に寄与する身体構造の抽出
ロボットによる実験により観察された複数の協調的 運動パターンの実現に寄与している身体構造抽出を目 的として,動力学シミュレーションによる異なる形態 間の比較及び情報論的解析を行った.具体的には,多 関節筋による感覚情報の相関に着目し,ロボットの生 体規範の筋配置と,全ての多関節筋を単関節筋に置き 換え単関節筋のみで構成された筋配置との間で比較を 行った.二つのモデルを神経系モデルで駆動し得られ た感覚運動情報を用いて,感覚から運動への情報の流 れ及び創発した運動パターンの情報構造を解析した. まず,感覚から運動の情報流れをTransfer entropy を用いて解析した[16].解析の結果,二関節筋の生む 感覚情報間の相関が,神経系モデルでは独立な筋間に, 身体を介した情報の流れが生じている事がわかった(図 7).この身体を介した両者の情報の流れの総量は,単 関節筋のみから構成される身体モデルに対し,二関節 筋による身体モデルでは63% の増加が確認された.
運動パターンの情報構造解析について,まず二つの 筋間の協調関係という観点から,位相同期率について 解析を行った[15].この結果,前脚,後脚における全組 み合わせのうち,三つのペアが二関節筋を有する身体 モデルにおいて,2.5, 1.7, 3.1 倍の増加を示し,残りの 一つのペアにおいて11% の減少が確認された (図 8).
更に運動パターンの大域構造について,情報量に基 づく指標であるIntegration 及び Complexity を用いて 解析を行った[17][18].時系列データの集合から成るシ
(a) 単関節筋のみの身体モデル (b) 二関節筋を有する身体モ デル
図7 シミュレーションによる感覚から運動への情報の 流れの比較.
ステムX = (x1, x2, . . . , xN) に対して,Integration は以下の式で定義される.
I(X) =∑
i
H(xi) − H(X). (5)
ここで,H(·) はエントロピーである.Integration は相 互情報量の多次元への一般化であり,要素間の統計的 依存関係の度合いを測る指標である.要素間が独立な 場合には相互情報量と同様に0 となる.
一方でComplexity は,以下の式により計算される. C(X) = H(X) −∑
i
H(xi|X − xi). (6)
ここで,H(xi|X −xi) は条件付きエントロピーで,X − xiはX から xiを除いたシステムを示す.統計的依存 関係のネットワーク構造の分化及び統合の度合いを測 る指標で,ランダムや規則的な場合低く,局所的及び大 域的構造を併せ持つ時に高い値となる指標である.こ の両方の指標において,二関節筋を有する身体モデル が,単関節筋のみから成る身体モデルに比較して,約 2 倍の増加を示した (図 9).
5. 結論
本研究では,生物が身体・神経系の相互作用を通し て実現する運動スキルの形成について,多面的,総合 的理解を目指し,生物規範型の四脚ロボットを用いた 構成論的アプローチの提案を行った.
我々はまず,事前の明示的な運動表現や筋間の関係 性を決める要素なしに,身体・神経系の相互作用を通
図8シミュレーションにおける位相同期率の比較.
(a) Integration (b) Complexity
図9シミュレーションにおける創発した運動の情報構造.
して複数運動パターンが自律的に創発可能である事を 示した.これらの運動の中には,ロコモーション様運 動を例とした有意味な運動の創発も観察された.この 結果はランダムな運動生成により実現されたのではな く,身体ダイナミクスを介した情報のカップリングに よる協調関係が自律的に形成されたことによるもので ある事を示した.
更に,これらの「原初的運動」創発の背後にあるメ カニズムへの理解を深めるために,シミュレーション による解析を行った.二関節筋を有する生物規範型の 身体モデルと,単関節筋のみから成る身体モデルの比 較を通して,二関節筋の生む感覚情報間の相関が,神 経系モデルでは独立な筋間に身体を通した情報の流れ を生みだし,その結果協調的でありながら構造を持ち, 複雑な運動パターンの創発を可能にしている事を示し た.二関節筋は生物一般に多数存在するが,その機能 的意味については十分に理解されていない[19][20].そ の為この結果は,新たな提案を実証的かつ定量的に示 したという点において重要な成果である.
我々の提案する生物学的アプローチは,四脚に固有 な設計は入っていない.従って,得られた一連の結果 はより一般的な生物の身体・神経系の相互作用による 動的な運動パターン発現の機序へと迫る実証的理解と, その新しいアプローチの方法を示している.
今後の課題としては,多様な運動パターンからの分 化として,一貫した運動パターンの自律的獲得による, 実環境下での適応的な運動の実現が挙げられる. 参考文献
[1] R. A. Brooks, “Intelligence without representation,” Artificial Intelligence, vol. 47, no. 1-3, pp. 139–159, 1991.
[2] R. Pfeifer and J. C. Bongard, How the Body Shapes the Way We Think: A New View of Intelligence. MIT Press, 2006.
[3] E. Bizzi, V. Cheung, H. d ´Avella, P. Salitel, and M. Tresch, “Combining modules for movement,” Brain Research Reviews, vol. 57, pp. 125–133, 2007.
[4] J. Kelso, “Synergies: atoms of brain and behavior,” in Progress in Motor Control: A Multidisciplinary Per- spective, D. Sternad, Ed. Springer Verlag, 2009, pp. 83–91.
[5] S. Grillner, “Biological Pattern Generation: The Cel- lular and Computational Logic of Networks in Mo- tion,” Neuron, vol. 52, no. 5, pp. 751–766, 2006. [6] M. H. Johnson, Developmental Cognitive Neuro-
science. Blackwell Publishing, 2005.
[7] 多賀厳太郎, “脳と身体の動的デザイン運動・知覚の非線 形力学と発達”,金子書房, 2002.
[8] H. F. R. Prechtl, “State of the art of a new functional assessment of the young nervous system. an early pre- dictor of cerebral palsy,” Early Human Development, vol. 50, no. 1, pp. 1–11, 1997.
[9] D. Caldwell, G. Medrano-Cerda, and M. Goodwin,
“Control of pneumatic muscle actuators,” Control Systems, IEEE, vol. 15, no. 1, pp. 40–48, 1995. [10] G. Klute, J. Czerniecki, and B. Hannaford, “McK-
ibben artificial muscles: pneumatic actuators with biomechanical intelligence,” in Proc. IEEE/ASME International Conference on Advanced Intelligent Mechatronics, 1999, pp. 221–226.
[11] R. A. Miller, “Evolution of the pectoral girdle and fore limb in the primates,” American Journal of Phys- ical Anthropology, vol. 17, no. 1, pp. 1–56, 1932. [12] C. P. Chou and B. Hannaford, “Measurement and
modeling of McKibben pneumatic artificial muscles,” IEEE Transactions on Robotics and Automation, vol. 12, no. 1, pp. 90–102, 1996.
[13] G. K. Klute, J. M. Czerniecki, and B. Hannaford,
“Artificial muscles: Actuators for biorobotic sys- tems,” The International Journal of Robotics Re- search, vol. 21, no. 4, pp. 295–309, 2002.
[14] Y. Kuniyoshi and S. Sangawa, “Early motor de- velopment from partially ordered neural-body dy- namics: experiments with a cortico-spinal-musculo- sleletal model,” Biological Cybernetics, vol. 95, pp. 589–605, 2006.
[15] M. Rosenblum, A. Pikovsky, J. Kurths, C. Sch¨afer, and P. Tass, “Chapter 9 phase synchronization: From theory to data analysis,” in Neuro-Informatics and Neural Modelling, ser. Handbook of Biological Physics, F. Moss and S. Gielen, Eds. North-Holland, 2001, vol. 4, pp. 279–321.
[16] T. Schreiber, “Measuring information transfer,” Physical Review Letters, vol. 85, no. 2, pp. 461–464, January 2000.
[17] W. McGill, “Multivariate information transmission,” Information Theory, IRE Professional Group on, vol. 4, no. 4, pp. 93–111, 1954.
[18] G. Tononi, O. Sporns, and G. M. Edelman, “A mea- sure for brain complexity: relating functional segre- gation and integration in the nervous system,” Pro- ceedings of the National Academy of Sciences of the United States of America, vol. 91, no. 11, pp. 5033– 5037, 1994.
[19] G. J. van Ingen Schenau, M. F. Bobbert, and R. H. Rozendal, “The unique action of bi-articular muscles in complex movements,” Journal of Anatomy, vol. 155, pp. 1–5, December 1987.
[20] C. A. M. Doorenbosch and G. J. van Ingen Schenau,
“The role of mono- and bi-articular muscles during contact control leg tasks in man,” Human Movement Science, vol. 14, no. 3, pp. 279–300, 1995.